
Effects of Dietary Supplementation of Peanut Skin Proanthocyanidins on Growth Performance and Lipid Metabolism of the Juvenile American Eel (Anguilla rostrata)

Abstract
:Simple Summary
Abstract
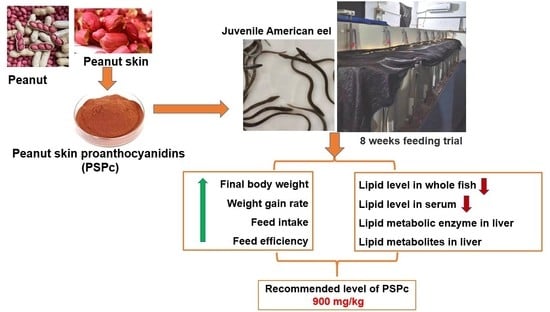
1. Introduction
2. Materials and Methods
2.1. Feeding Trial
2.2. Sample Collection
2.3. Calculations of Growth Performance Parameters
2.4. Proximate Composition Analysis
2.5. Measurement of Serum Lipids and Lipid Metabolic Enzymes in the Liver
2.6. Lipidomics Profiling in the Liver
2.7. Statistical Analysis
3. Results
3.1. Growth Performance and Proximate Composition of Whole Fish
3.2. Serum Lipid Parameters
3.3. Lipid Metabolic Enzymes in the Liver
3.4. Lipidomics Profiling in the Liver
3.5. Lipid Metabolism Pathways and Related Lipid Metabolites
4. Discussion
4.1. Effects of Dietary PSPc Supplementation on Growth Performance and Proximate Composition of Whole Fish of the Juvenile American Eel
4.2. Effects of Dietary PSPc Supplementation on Serum Lipids and Lipid Metabolic Enzymes of the Juvenile American Eel
4.3. Effects of Dietary PSPc Supplementation on Metabolites and Metabolic Pathways Involved in Lipid Metabolism in the Liver of the Juvenile American Eel
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mannino, G.; Chinigò, G.; Serio, G.; Genova, T.; Gentile, C.; Munaron, L.; Bertea, C.M. Proanthocyanidins and where to find them: A Meta-analytic approach to investigate their chemistry, biosynthesis, distribution, and effect on human health. Antioxidants 2021, 10, 1229. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Iahtisham-Ul-Haq; Patel, S.; Pan, X.; Naz, S.; Sanches Silva, A.; Saeed, F.; Rasul Suleria, H.A. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.H.; Cai, G.H.; Zhai, S.W. Grape seed proanthocyanidin extract regulates lipid metabolism of the American eel (Anguilla rostrata). Nat. Prod. Res. 2021; 1–5, online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.W.; Lu, J.J.; Chen, X.H. Effects of dietary grape seed proanthocyanidins on growth performance, some serum biochemical parameters and body composition of tilapia (Oreochromis niloticus) fingerlings. Ital. J. Anim. Sci. 2014, 13, 536–540. [Google Scholar] [CrossRef]
- Arslan, G.; Sönmez, A.Y.; Yank, T. Effects of grape Vitis vinifera seed oil supplementation on growth, survival, fatty acid profiles, antioxidant contents and blood parameters in rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2018, 49, 2256–2266. [Google Scholar] [CrossRef]
- Kesbiç, O.S.; Yigit, M. Structural and chemical changes of grape seed extract after thermal processing and its use in rainbow trout (Oncorhynchus mykiss) diets as an organic feed supplement. Aquaculture 2019, 503, 275–281. [Google Scholar] [CrossRef]
- Zhai, S.W.; Wang, Y.; He, Y.X.; Chen, X.H. Oligomeric proanthocyanidins counteracts the negative effects of high level of dietary histamine on American eel (Anguilla rostrata). Front. Mar. Sci. 2020, 7, 549145. [Google Scholar] [CrossRef]
- Mehrinakhi, Z.; Ahmadifar, E.; Sheikhzadeh, N.; Moghadam, M.S.; Dawood, M.A. Extract of grape seed enhances the growth performance, humoral and mucosal immunity, and resistance of common carp (Cyprinus carpio) against. Ann. Anim. Sci. 2021, 21, 217–232. [Google Scholar] [CrossRef]
- Mohammadi, Y.; Kamangar, B.B.; Zarei, M.A. Effects of diets containing grape seed proanthocyanidin extract on the growth and oxidative capacity of common carp (Cyprinus carpio). Aquaculture 2021, 540, 736689. [Google Scholar] [CrossRef]
- Xu, G.; Xing, W.; Li, T.; Yu, H.; Wei, S.; Jiang, N.; Ma, Z.; Lou, L. Dietary grape seed proanthocyanidins improved growth, immunity, antioxidant, digestive enzymes activities, and intestinal microbiota of juvenile hybrid sturgeon (Acipenser baeri Brandt♀× A. schrenckii Brandt♂). Aquac. Nutr. 2021, 27, 1983–1995. [Google Scholar] [CrossRef]
- Lu, R.H.; Qin, C.B.; Yang, F.; Zhang, W.Y.; Zhang, Y.R.; Yang, G.K.; Yang, L.P.; Meng, X.L.; Yan, X.; Nie, G.X. Grape seed proanthocyanidin extract ameliorates hepatic lipid accumulation and inflammation in grass carp (Ctenopharyngodon idella). Fish. Physiol. Biochem. 2020, 46, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Toomer, O.T.; Vu, T.; Pereira, M.; Williams, K. Dietary supplementation with peanut skin polyphenolic extracts (PSPE) reduces hepatic lipid and glycogen stores in mice fed an atherogenic diet. J. Funct. Foods 2019, 55, 362–370. [Google Scholar] [CrossRef]
- Bansode, R.R.; Randolph, P.; Ahmedna, M.; Williams, L.L.; Yu, J. Bioavailability and hypolipidemic effects of peanut skin polyphenols. J. Med. Food 2015, 18, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Yuan, Y.; Dai, Y.; Gong, Y.; Yuan, Y. Development status and trends in the eel farming industry in Asia. N. Am. J. Aquac. 2022, 84, 3–17. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, P.; Chen, X.H.; Zhai, S.W. Effects of dietary proanthocyanidins supplementation on growth performance, digestive enzymes activities and microbiota in the intestine of juvenile American eels (Anguilla rostrata) cultured in cement tanks. Isr. J. Aquacult-Bamid. 2020, 72, 1226208. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Yang, H.L.; Hu, L.H.; Yang, W.; Ai, C.X.; Sun, Y.Z. Dose-dependent effects of histamine on growth, immunity and intestinal health in juvenile grouper (Epinephelus coioides). Front. Mar. Sci. 2021, 8, 685720. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis of Official Analytical Chemists International, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2002. [Google Scholar]
- Ma, Z.R.; Zhao, P.Y.; Zhai, S.W. Effects of dietary bile acids supplementation on lipid metabolism in liver of European eel (Anguilla anguilla) juveniles. Acta Hydrobiol. Sin. 2022; 1–8, online ahead of print. [Google Scholar] [CrossRef]
- Li, M.; Ren, W.; Chai, W.; Zhu, M.; Man, L.; Zhan, Y.; Qin, H.; Sun, M.; Liu, J.; Zhang, D.; et al. Comparing the profiles of raw and cooked donkey meat by metabonomics and lipidomics assessment. Front. Nutr. 2022, 9, 851761. [Google Scholar] [CrossRef]
- Jia, Z.; Tan, Y.; Liu, Y.; Cai, G.; Chen, X.; Zhai, S. Grape seed proanthocyanidins alleviate the negative effects of dietary cadmium on pearl gentian grouper (Epinephelus fuscoguttatus female x Epinephelus lanceolatus male). Isr. J. Aquacult-Bamid. 2021, 73, 1427267. [Google Scholar] [CrossRef]
- Zhai, S.W.; Lu, J.J.; Zhao, P.Y.; Chen, X.H. Effect of grape seed proanthocyanidins on alleviating dietary cadmium (Cd) induced growth retardation and oxidative stress in hepatopancreas of juvenile tilapia (Oreochromis niloticus). Isr. J. Aquacult-Bamid. 2018, 70, 1513. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Ashour, E.A.; Elaraby, G.M.; Osman, A.O.; Arif, M. Influences of dietary supplementation of peanut skin powder (Arachis Hypogaea) on growth performance, carcass traits, blood chemistry, antioxidant activity and meat quality of broilers. Anim. Prod. Sci. 2018, 58, 965–972. [Google Scholar] [CrossRef]
- Zhang, R.; Song, X.; Yu, J.; Meng, W.; Li, C. The research progress of the procyanidins from peanut skin coat. Food Res. Dev. 2020, 41, 202–210. [Google Scholar] [CrossRef]
- Mousavi, S.; Sheikhzadeh, N.; Tayefi-Nasrabadi, H.; Alizadeh-Salteh, S.; Khani Oushani, A.; Firouzamandi, M.; Mardani, K. Administration of grape (Vitis vinifera) seed extract to rainbow trout (Oncorhynchus mykiss) modulates growth performance, some biochemical parameters, and antioxidant-relevant gene expression. Fish. Physiol. Biochem. 2020, 46, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Shimizu-Ibuka, A.; Udagawa, H.; Kobayashi-Hattori, K.; Mura, K.; Tokue, C.; Takita, T.; Arai, S. Hypocholesterolemic effect of peanut skin and its fractions: A case record of rats fed on a high-cholesterol diet. Biosci. Biotech. Bioch. 2009, 73, 205–208. [Google Scholar] [CrossRef]
- Yang, Q.; Ding, M.; Tan, B.; Dong, X.; Chi, S.; Zhang, S.; Liu, H. Effects of dietary vitamin A on growth, feed utilization, lipid metabolism enzyme activities, and fatty acid synthase and hepatic lipase mRNA expression levels in the liver of juvenile orange spotted grouper, Epinephelus coioides. Aquaculture 2017, 479, 501–507. [Google Scholar] [CrossRef]
- He, L.; Qin, Y.; Wang, Y.; Li, D.; Chen, W.; Ye, J. Effects of dietary replacement of fish oil with soybean oil on the growth performance, plasma components, fatty acid composition and lipid metabolism of groupers Epinephelus coioides. Aquac. Nutr. 2021, 27, 1494–1511. [Google Scholar] [CrossRef]
- Dong, G.F.; Zou, Q.; Wang, H.; Huang, F.; Liu, X.C.; Chen, L.; Yang, C.Y.; Yang, Y.O. Conjugated linoleic acid differentially modulates growth, tissue lipid deposition, and gene expression involved in the lipid metabolism of grass carp. Aquaculture 2014, 432, 181–191. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, F.; Jin, M.; Wang, X.; Hu, X.; Zhao, M.; Chen, X.; Lou, J.; Jiao, L.; Betancor, M.B.; et al. Untargeted lipidomics reveals metabolic responses to different dietary n-3 PUFA in juvenile swimming crab (Portunus trituberculatus). Food Chem. 2021, 354, 129570. [Google Scholar] [CrossRef]
- Jia, R.; Cao, L.P.; Du, J.L.; He, Q.; Gu, Z.Y.; Jeney, G.; Xu, P.; Yin, G.J. Effects of high-fat diet on steatosis, endoplasmic reticulum stress and autophagy in liver of tilapia (Oreochromis niloticus). Front. Mar. Sci. 2020, 7, 363. [Google Scholar] [CrossRef]
- Guo, Y.F.; Su, T.; Yang, M.; Li, C.J.; Guo, Q.; Xiao, Y.; Huang, Y.; Liu, Y.; Luo, X.H. The role of autophagy in bone homeostasis. J. Cell Physiol. 2021, 236, 4152–4173. [Google Scholar] [CrossRef]
- Shin, D.W. Lipophagy: Molecular mechanisms and implications in metabolic disorders. Mol. Cells 2020, 43, 686. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.; Czaja, M. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Nebauer, R.; Rosenberger, S.; Daum, G. Phosphatidylethanolamine, a limiting factor of autophagy in yeast strains bearing a defect in the carboxypeptidase Y pathway of vacuolar targeting. J. Biol. Chem. 2007, 282, 16736–16743. [Google Scholar] [CrossRef]
- Ran, G.; Ying, L.; Li, L.; Yan, Q.; Yi, W.; Ying, C.; Wu, H.; Ye, X. Resveratrol ameliorates diet-induced dysregulation of lipid metabolism in zebrafish (Danio rerio). PLoS ONE 2017, 12, e0180865. [Google Scholar] [CrossRef] [PubMed]
- Régnier, M.; Polizzi, A.; Guillou, H.; Loiseau, N. Sphingolipid metabolism in non-alcoholic fatty liver diseases. Biochimie 2019, 159, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Chocian, G.; Chabowski, A.; Żendzian-Piotrowska, M.; Harasim, E.; Łukaszuk, B.; Górski, J. High fat diet induces ceramide and sphingomyelin formation in rat’s liver nuclei. Mol. Cell. Biochem. 2010, 340, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R.; Bendiksen, E.Å.; Campbell, P.J.; Bell, J.G. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell. Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Cullis, P.R.; Hope, M.J. Physical properties and functional roles of lipids in membranes. New Compreh. Biochem. 1991, 20, 1–41. [Google Scholar] [CrossRef]
- Tian, J.; Wen, H.; Lu, X.; Liu, W.; Wu, F.; Yang, C.G.; Jiang, M.; Yu, L. Dietary phosphatidylcholine impacts on growth performance and lipid metabolism in adult Genetically Improved Farmed Tilapia (GIFT) strain of Nile tilapia Oreochromis niloticus. Br. J. Nutr. 2018, 119, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Hermansson, M.; Hokynar, K.; Somerharju, P. Mechanisms of glycerophospholipid homeostasis in mammalian cells. Prog. Lipid Res. 2011, 50, 240–257. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.T.; Ramesh, T.; Toh, X.R.; Nguyen, L.N. Emerging roles of lysophospholipids in health and disease. Prog. Lipid Res. 2020, 80, 101068. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Lin, F.; Liu, G.; Yu, J.; Li, S.; Wen, X. The suppressive mechanism of dietary macroalgae Saccharina japonica against hepatic lipid accumulation in black seabream A. schlegelii. Aquaculture 2022, 547, 737357. [Google Scholar] [CrossRef]
- Yang, X.X.; Wei, J.D.; Mu, J.K.; Liu, X.; Dong, J.C.; Zeng, L.X.; Gu, W.; Li, J.P.; Yu, J. Integrated metabolomic profiling for analysis of antilipidemic effects of Polygonatum kingianum extract on dyslipidemia in rats. World. J. Gastroenterol. 2018, 24, 5505–5524. [Google Scholar] [CrossRef]
- Wang, S.; Chen, W.; Ruan, D.; Wang, S.L.; Lin, Y.C.; Zheng, C.T. Effects of dietary linoleic acid level on laying performance, egg quality and lipids metabolism of ducks during the early laying period. Chin. J. Anim. Nutr. 2015, 27, 731–739. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, N.; Song, P.; He, T.; Levesque, C.; Bai, Y.Y.; Zhang, A.Z.; Ma, X. Grape seed proanthocyanidin affects lipid metabolism via changing gut microflora and enhancing propionate production in weaned pigs. J. Nutr. 2019, 149, 1523–1532. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Groups | ||||
|---|---|---|---|---|---|
| PSPc0 | PSPc300 | PSPc600 | PSPc900 | PSPc1200 | |
| IBW (g/fish) | 10.48 ± 0.04 a | 10.50 ± 0.02 a | 10.54 ± 0.04 a | 10.45 ± 0.02 a | 10.53 ± 0.03 a |
| FBW (g/fish) | 16.40 ± 0.29 a | 17.45 ± 0.28 b | 17.96 ± 0.22 b | 19.22 ± 0.54 c | 17.97 ± 0.65 b |
| WGR (%) | 56.67 ± 2.45 a | 66.58 ± 3.28 b | 70.27 ± 1.89 b | 83.90 ± 5.44 c | 70.68 ± 5.67 b |
| FE (%) | 55.68 ± 1.89 a | 56.04 ± 2.44 a | 62.30 ± 2.90 b | 67.51 ± 3.14 c | 63.81 ± 2.23 bc |
| FR (%) | 1.41 ± 0.01 a | 1.51 ± 0.04 bc | 1.46 ± 0.05 ab | 1.56 ± 0.05 c | 1.44 ± 0.04 ab |
| SR (%) | 100 a | 96.67 ± 3.51 a | 99.00 ± 1.73 a | 100 a | 99.00 ± 1.73 a |
| Item | Groups | ||||
|---|---|---|---|---|---|
| PSPc0 | PSPc300 | PSPc600 | PSPc900 | PSPc1200 | |
| Moisture (%) | 69.03 ± 1.14 a | 68.37 ± 0.67 a | 69.13 ± 0.50 a | 68.20 ± 1.76 a | 68.27 ± 1.48 a |
| Crude lipids (%) | 9.94 ± 0.47 c | 9.64 ± 0.10 c | 9.00 ± 0.10 b | 8.41 ± 0.31 a | 8.82 ± 0.16 ab |
| Crude protein (%) | 18.44 ± 0.37 a | 18.78 ± 0.27 a | 18.85 ± 0.38 a | 18.88 ± 0.31 a | 18.69 ± 0.53 a |
| Ash (%) | 3.37 ± 0.26 a | 3.35 ± 0.07 a | 3.32 ± 0.20 a | 3.49 ± 0.19 a | 3.35 ± 0.17 a |
| Item | Groups | ||||
|---|---|---|---|---|---|
| PSPc0 | PSPc300 | PSPc600 | PSPc900 | PSPc1200 | |
| TC (mmol/L) | 12.77 ± 1.46 b | 12.67 ± 0.40 b | 12.07 ± 0.45 ab | 10.67 ±0.33 a | 11.84 ± 1.39 ab |
| TG (mmol/L)) | 7.13 ± 061 c | 5.90 ± 0.50 b | 5.04 ± 0.48 b | 3.77 ± 0.42 a | 5.21 ± 0.92 b |
| LDL-C (mmol/L) | 2.35 ± 0.30 c | 1.94 ± 0.12 b | 1.84 ± 0.24 b | 1.44 ± 0.22 a | 1.78 ± 0.11 ab |
| HDL-C (mmol/L) | 2.12 ± 0.36 a | 2.79 ± 0.13 b | 3.12 ± 0.60 b | 5.50 ± 0.33 c | 3.38 ± 0.23 b |
| Item | Groups | ||||
|---|---|---|---|---|---|
| PSPc0 | PSPc300 | PSPc600 | PSPc900 | PSPc1200 | |
| LPL (U/mgprot) | 6.53 ± 0.31 a | 7.18 ± 0.17 a | 9.52 ± 0.33 b | 10.87 ± 0.77 c | 9.83 ± 0.52 b |
| HL (U/mgprot) | 9.06 ± 0.50 a | 9.18 ± 0.32 a | 11.42 ± 0.61 b | 13.00 ± 0.38 c | 11.24 ± 0.79 b |
| TL (U/mgprot) | 15.59 ± 0.22 a | 16.36 ± 0.47 a | 20.94 ± 0.89 b | 23.87 ± 1.15 c | 21.08 ± 1.09 b |
| FAS (ng/mgprot) | 1.76 ± 0.08 c | 1.27 ± 0.09 b | 1.20 ± 0.04 b | 1.05 ± 0.02 a | 1.21 ± 0.10 b |
| ACC (ng/mgprot) | 1.28 ± 0.10 c | 1.03 ± 0.10 b | 1.01 ± 0.12 b | 0.81 ± 0.09 a | 1.03 ± 0.04 b |
| Differential Lipid Metabolites | VIP | p-Value | log2 (FC) |
|---|---|---|---|
| Methyl phosphatidylcholine (MePC) | |||
| 35:3 | 12.7635 | 0.0092 | 0.3470 |
| 34:3 | 7.1823 | 0.0092 | 0.3913 |
| 32:2 | 4.9457 | 0.0092 | 0.6206 |
| Phosphatidylcholine (PC) | |||
| 38:6 | 12.7635 | 0.0092 | 0.3470 |
| 19:1_15:0 | 12.1506 | 0.0092 | 0.2950 |
| 34:1 | 11.7509 | 0.0092 | 0.2760 |
| 38:5 | 7.2244 | 0.0092 | 0.3623 |
| 35:6 | 6.4893 | 0.0092 | 0.3596 |
| 35:5 | 4.9457 | 0.0092 | 0.6206 |
| 20:5_13:0 | 3.0010 | 0.0092 | 0.4176 |
| Sphingomyelin (SM) | |||
| d16:1_24:1 | 8.7552 | 0.0092 | 0.2740 |
| Phosphatidylethanolamine (PE) | |||
| 18:1_18:2 | 6.4893 | 0.0092 | 0.3596 |
| 16:0_18:2 | 3.0517 | 0.0092 | 0.4240 |
| Phosphatidylethanol (PEt) | |||
| 16:0_22:6 | 7.6820 | 0.0092 | 3.4540 |
| 18:1_22:6 | 5.5258 | 0.0092 | 3.1546 |
| 16:0_20:5 | 4.2147 | 0.0092 | 3.9162 |
| 16:0_18:1 | 3.5621 | 0.0092 | 3.3659 |
| 18:1_22:5 | 3.1935 | 0.0092 | 2.6640 |
| Lysophosphatidylethanol (LPEt) | |||
| 22:6 | 4.3802 | 0.0092 | 3.0020 |
| Phosphatidylmethanol (PMe) | |||
| 19:0_22:6 | 3.1865 | 0.0092 | 2.6425 |
| Pathway Name | Total | Hits | Lipid Metabolite | Up/Down |
|---|---|---|---|---|
| Autophagy | 6 | 1 | Phosphatidylethanolamine (PE) | Up |
| Sphingolipid metabolism | 25 | 1 | Ceramides (Cer) | Down |
| Phosphatidylcholine (PC) | Up | |||
| Glycerophospholipid metabolism | 52 | 3 | Phosphatidylethanolamine (PE) | Up |
| 2-Lysophosphatidylcholine (2-LPC) | Down | |||
| Linoleic acid metabolism | 28 | 1 | Phosphatidylcholine (PC) | Up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Chen, X.-H.; Wu, X.-Y.; Cai, G.-H.; Zhai, S.-W. Effects of Dietary Supplementation of Peanut Skin Proanthocyanidins on Growth Performance and Lipid Metabolism of the Juvenile American Eel (Anguilla rostrata). Animals 2022, 12, 2375. https://doi.org/10.3390/ani12182375
Wang Y, Chen X-H, Wu X-Y, Cai G-H, Zhai S-W. Effects of Dietary Supplementation of Peanut Skin Proanthocyanidins on Growth Performance and Lipid Metabolism of the Juvenile American Eel (Anguilla rostrata). Animals. 2022; 12(18):2375. https://doi.org/10.3390/ani12182375
Chicago/Turabian StyleWang, Yue, Xue-Hao Chen, Xin-Yi Wu, Guo-He Cai, and Shao-Wei Zhai. 2022. "Effects of Dietary Supplementation of Peanut Skin Proanthocyanidins on Growth Performance and Lipid Metabolism of the Juvenile American Eel (Anguilla rostrata)" Animals 12, no. 18: 2375. https://doi.org/10.3390/ani12182375
APA StyleWang, Y., Chen, X.-H., Wu, X.-Y., Cai, G.-H., & Zhai, S.-W. (2022). Effects of Dietary Supplementation of Peanut Skin Proanthocyanidins on Growth Performance and Lipid Metabolism of the Juvenile American Eel (Anguilla rostrata). Animals, 12(18), 2375. https://doi.org/10.3390/ani12182375