
The Effects of Oncolytic Pseudorabies Virus Vaccine Strain Inhibited the Growth of Colorectal Cancer HCT-8 Cells In Vitro and In Vivo
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. In Vitro Virus Replication and Virus Titration
2.3. Evaluation of Cytopathic Effects (CPEs) In Vitro
2.4. Cell Viability Assay
2.5. Animal Experiments
2.6. Ethics Statement
2.7. Real-Time PCR for the Detection of PRV
2.8. Histological Examination
2.9. Immunohistochemistry
2.10. Flow Cytometry
2.11. Statistical Analysis
3. Results
3.1. Replication of PRV in HCT-8 Cells
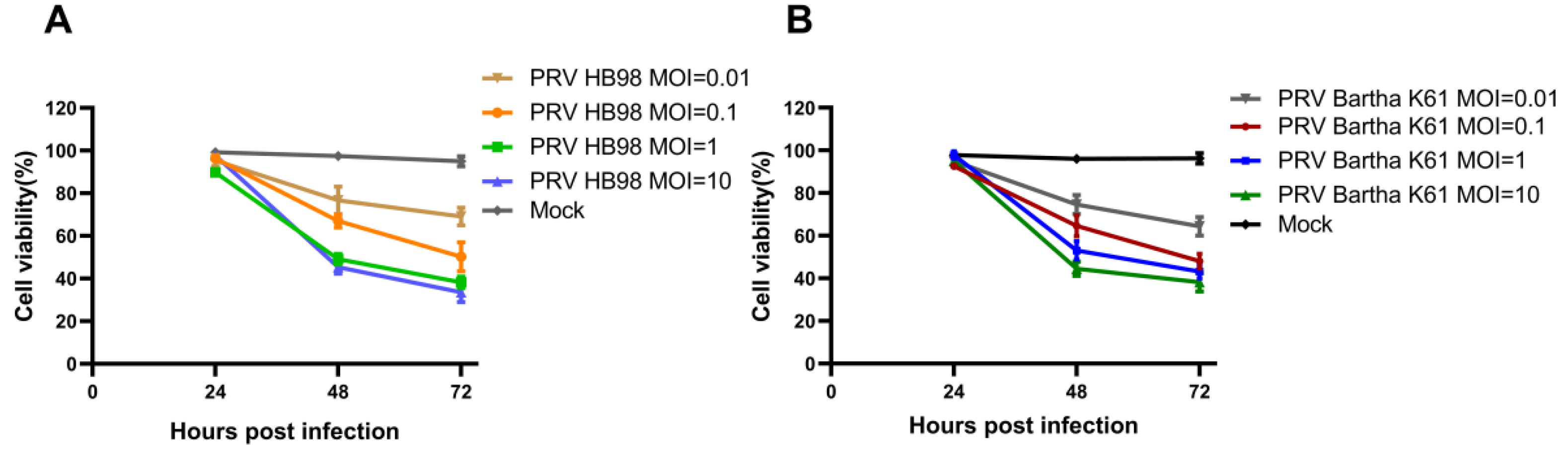
3.2. In Vitro Cell Viability
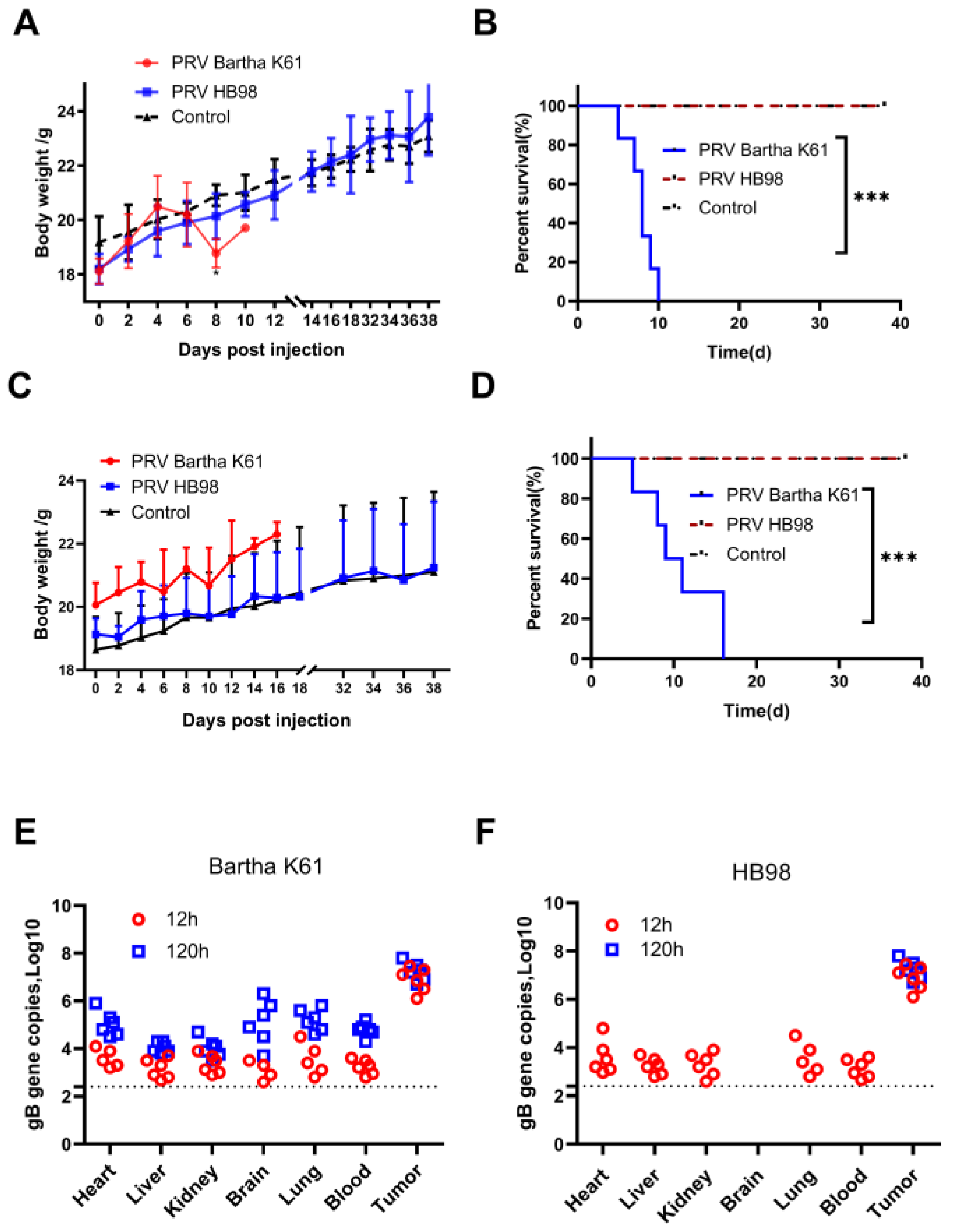
3.3. PRV Reduces Tumor Growth in BALB/c nu Mice Model
3.4. Evaluation of the Safety of PRV In Vivo
3.5. Pathological and Immunohistochemical Study of Colorectal Cancer Cells Treated with PRV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Kana, S.I.; Essani, K. Immuno-Oncolytic Viruses: Emerging Options in the Treatment of Colorectal Cancer. Mol. Diagn. Ther. 2021, 25, 301–313. [Google Scholar] [CrossRef]
- Aurelian, L.; Bollino, D.; Colunga, A. The oncolytic virus DeltaPK has multimodal anti-tumor activity. Pathog. Dis. 2016, 74, ftw050. [Google Scholar] [CrossRef]
- Lv, Y.; Zhou, D.; Hao, X.Q.; Zhu, M.Y.; Zhang, C.D.; Zhou, D.M.; Wang, J.H.; Liu, R.X.; Wang, Y.L.; Gu, W.Z.; et al. A recombinant measles virus vaccine strain rMV-Hu191 has oncolytic effect against human gastric cancer by inducing apoptotic cell death requiring integrity of lipid raft microdomains. Cancer Lett. 2019, 460, 108–118. [Google Scholar] [CrossRef]
- Haddad, D.; Chen, N.; Zhang, Q.; Chen, C.H.; Yu, Y.A.; Gonzalez, L.; Aguilar, J.; Li, P.; Wong, J.; Szalay, A.A.; et al. A novel genetically modified oncolytic vaccinia virus in experimental models is effective against a wide range of human cancers. Ann. Surg. Oncol. 2012, 19 (Suppl. S3), S665–S674. [Google Scholar] [CrossRef]
- Islam, S.; Lee, B.; Jiang, F.; Kim, E.K.; Ahn, S.C.; Hwang, T.H. Engineering and Characterization of Oncolytic Vaccinia Virus Expressing Truncated Herpes Simplex Virus Thymidine Kinase. Cancers 2020, 12, 228. [Google Scholar] [CrossRef]
- Cho, E.; Ryu, E.J.; Jiang, F.; Jeon, U.B.; Cho, M.; Kim, C.H.; Kim, M.; Kim, N.D.; Hwang, T.H. Preclinical safety evaluation of hepatic arterial infusion of oncolytic poxvirus. Drug Des. Dev. Ther. 2018, 12, 2467–2474. [Google Scholar] [CrossRef]
- Tong, W.; Li, G.; Liang, C.; Liu, F.; Tian, Q.; Cao, Y.; Li, L.; Zheng, X.; Zheng, H.; Tong, G. A live, attenuated pseudorabies virus strain JS-2012 deleted for gE/gI protects against both classical and emerging strains. Antivir. Res. 2016, 130, 110–117. [Google Scholar] [CrossRef]
- Freuling, C.M.; Muller, T.F.; Mettenleiter, T.C. Vaccines against pseudorabies virus (PrV). Vet. Microbiol. 2017, 206, 3–9. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A Novel Human Acute Encephalitis Caused by Pseudorabies Virus Variant Strain. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, e3690–e3700. [Google Scholar] [CrossRef]
- Wang, Y.; Nian, H.; Li, Z.; Wang, W.; Wang, X.; Cui, Y. Human encephalitis complicated with bilateral acute retinal necrosis associated with pseudorabies virus infection: A case report. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2019, 89, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.W.; Weng, S.S.; Cheng, Q.; Cui, P.; Li, Y.J.; Wu, H.L.; Zhu, Y.M.; Xu, B.; Zhang, W.H. Human Endophthalmitis Caused By Pseudorabies Virus Infection, China, 2017. Emerg. Infect. Dis. 2018, 24, 1087–1090. [Google Scholar] [CrossRef] [PubMed]
- Shiau, A.L.; Lin, Y.P.; Shieh, G.S.; Su, C.H.; Wu, W.L.; Tsai, Y.S.; Cheng, C.W.; Lai, M.D.; Wu, C.L. Development of a conditionally replicating pseudorabies virus for HER-2/neu-overexpressing bladder cancer therapy. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Delva, J.L.; Nauwynck, H.J.; Mettenleiter, T.C.; Favoreel, H.W. The Attenuated Pseudorabies Virus Vaccine Strain Bartha K61: A Brief Review on the Knowledge Gathered During 60 Years of Research. Pathogens 2020, 9, 897. [Google Scholar] [CrossRef]
- Zheng, H.H.; Wang, L.Q.; Fu, P.F.; Zheng, L.L.; Chen, H.Y.; Liu, F. Characterization of a recombinant pseudorabies virus expressing porcine parvovirus VP2 protein and porcine IL-6. Virol. J. 2020, 17, 19. [Google Scholar] [CrossRef]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. MMBR 2005, 69, 462–500. [Google Scholar] [CrossRef]
- Cuddington, B.P.; Verschoor, M.; Ashkar, A.; Mossman, K.L. Enhanced efficacy with azacytidine and oncolytic BHV-1 in a tolerized cotton rat model of breast adenocarcinoma. Mol. Ther. Oncolytics 2015, 2, 15004. [Google Scholar] [CrossRef]
- Hong, B.; Sahu, U.; Mullarkey, M.P.; Kaur, B. Replication and Spread of Oncolytic Herpes Simplex Virus in Solid Tumors. Viruses 2022, 14, 118. [Google Scholar] [CrossRef]
- Reale, A.; Vitiello, A.; Conciatori, V.; Parolin, C.; Calistri, A.; Palu, G. Perspectives on immunotherapy via oncolytic viruses. Infect. Agents Cancer 2019, 14, 5. [Google Scholar] [CrossRef]
- Rehman, H.; Silk, A.W.; Kane, M.P.; Kaufman, H.L. Into the clinic: Talimogene laherparepvec (T-VEC), a first-in-class intratumoral oncolytic viral therapy. J. Immunother. Cancer 2016, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Pol, J.; Kroemer, G.; Galluzzi, L. First oncolytic virus approved for melanoma immunotherapy. Oncoimmunology 2016, 5, e1115641. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Guo, X.; Zhu, Q.; Chao, A.; Fu, P.; Wei, Z.; Wang, S.; Chen, H.; Cui, B. Construction and immunogenicity of a recombinant pseudorabies virus co-expressing porcine circovirus type 2 capsid protein and interleukin 18. Virus Res. 2015, 201, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Lomniczi, B.; Watanabe, S.; Ben-Porat, T.; Kaplan, A.S. Genome location and identification of functions defective in the Bartha vaccine strain of pseudorabies virus. J. Virol. 1987, 61, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, K.S.; Glushchenko, A.V.; Gulyaeva, M.A.; Bi, Y.; Chen, J.; Shi, W.; Adamenko, L.S.; Shestopalov, A.M. Intratumoral Virotherapy with Wild-Type Newcastle Disease Virus in Carcinoma Krebs-2 Cancer Model. Viruses 2021, 13, 552. [Google Scholar] [CrossRef]
- Brittle, E.E.; Reynolds, A.E.; Enquist, L.W. Two modes of pseudorabies virus neuroinvasion and lethality in mice. J. Virol. 2004, 78, 12951–12963. [Google Scholar] [CrossRef]
- Kraft, R.M.; Nguyen, M.L.; Yang, X.H.; Thor, A.D.; Blaho, J.A. Caspase 3 activation during herpes simplex virus 1 infection. Virus Res. 2006, 120, 163–175. [Google Scholar] [CrossRef]
- Ghose, J.; Dona, A.; Murtadha, M.; Gunes, E.G.; Caserta, E.; Yoo, J.Y.; Russell, L.; Jaime-Ramirez, A.C.; Barwick, B.G.; Gupta, V.A.; et al. Oncolytic herpes simplex virus infects myeloma cells in vitro and in vivo. Mol. Ther. Oncolytics 2021, 20, 519–531. [Google Scholar] [CrossRef]
- Warner, S.G.; O’Leary, M.P.; Fong, Y. Therapeutic oncolytic viruses: Clinical advances and future directions. Curr. Opin. Oncol. 2017, 29, 359–365. [Google Scholar] [CrossRef]
- Szpara, M.L.; Tafuri, Y.R.; Parsons, L.; Shamim, S.R.; Verstrepen, K.J.; Legendre, M.; Enquist, L.W. A wide extent of inter-strain diversity in virulent and vaccine strains of alphaherpesviruses. PLoS Pathog. 2011, 7, e1002282. [Google Scholar] [CrossRef]
- Mettenleiter, T.C.; Zsak, L.; Kaplan, A.S.; Ben-Porat, T.; Lomniczi, B. Role of a structural glycoprotein of pseudorabies in virus virulence. J. Virol. 1987, 61, 4030–4032. [Google Scholar] [CrossRef] [Green Version]
- Husak, P.J.; Kuo, T.; Enquist, L.W. Pseudorabies Virus Membrane Proteins gI and gE Facilitate Anterograde Spread of Infection in Projection- Specific Neurons in the Rat. J. Virol. 2000, 74, 10975–10983. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.Q.; Tong, W.; Zheng, H.; Li, L.W.; Li, G.X.; Gao, F.; Wang, T.; Liang, C.; Ye, C.; Wu, J.Q.; et al. Variations in glycoprotein B contribute to immunogenic difference between PRV variant JS-2012 and Bartha-K61. Vet. Microbiol. 2017, 208, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kit, S.; Kit, M.; Pirtle, E.C. Attenuated properties of thymidine kinase-negative deletion mutant of pseudorabies virus. Am. J. Vet. Res. 1985, 46, 1359–1367. [Google Scholar]
- McGregor, S.; Easterday, B.C.; Kaplan, A.S.; Ben-Porat, T. Vaccination of swine with thymidine kinase-deficient mutants of pseudorabies virus. Am. J. Vet. Res. 1985, 46, 1494–1497. [Google Scholar] [PubMed]
- Croce, C.M.; Reed, J.C. Finally, An Apoptosis-Targeting Therapeutic for Cancer. Cancer Res. 2016, 76, 5914–5920. [Google Scholar] [CrossRef]
- Lawler, S.E.; Speranza, M.C.; Cho, C.F.; Chiocca, E.A. Oncolytic Viruses in Cancer Treatment: A Review. JAMA Oncol. 2017, 3, 841–849. [Google Scholar] [CrossRef]
- Loya, S.M.; Zhang, X. Enhancing the bystander killing effect of an oncolytic HSV by arming it with a secretable apoptosis activator. Gene Ther. 2015, 22, 237–246. [Google Scholar] [CrossRef]
- Wei, X.; Liu, L.; Wang, G.; Li, W.; Xu, K.; Qi, H.; Liu, H.; Shen, J.; Li, Z.; Shao, J. Potent antitumor activity of the Ad5/11 chimeric oncolytic adenovirus combined with interleukin-24 for acute myeloid leukemia via induction of apoptosis. Oncol. Rep. 2015, 33, 111–118. [Google Scholar] [CrossRef]
- Klupp, B.G.; Hengartner, C.J.; Mettenleiter, T.C.; Enquist, L.W. Complete, annotated sequence of the pseudorabies virus genome. J. Virol. 2004, 78, 424–440. [Google Scholar] [CrossRef]
- Olsen, L.M.; Ch’ng, T.H.; Card, J.P.; Enquist, L.W. Role of pseudorabies virus Us3 protein kinase during neuronal infection. J. Virol. 2006, 80, 6387–6398. [Google Scholar] [CrossRef]
- Lerma, L.; Alcala, S.; Pinero, C.; Torres, M.; Martin, B.; Lim, F.; Sainz, B., Jr.; Tabares, E. Expression of the immediate early IE180 protein under the control of the hTERT and CEA tumor-specific promoters in recombinant pseudorabies viruses: Effects of IE180 protein on promoter activity and apoptosis induction. Virology 2016, 488, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollmann, G.; Tattersall, P.; van den Pol, A.N. Targeting human glioblastoma cells: Comparison of nine viruses with oncolytic potential. J. Virol. 2005, 79, 6005–6022. [Google Scholar] [CrossRef] [PubMed]
- Boldogkoi, Z.; Bratincsak, A.; Fodor, I. Evaluation of pseudorabies virus as a gene transfer vector and an oncolytic agent for human tumor cells. Anticancer. Res. 2002, 22, 2153–2159. [Google Scholar] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chai, C.; Zhang, J.; Zhou, Y.; Yin, H.; Zhang, F.; Diao, Y.; Zan, X.; Ma, Y.; Wang, Y.; Wu, Y.; et al. The Effects of Oncolytic Pseudorabies Virus Vaccine Strain Inhibited the Growth of Colorectal Cancer HCT-8 Cells In Vitro and In Vivo. Animals 2022, 12, 2416. https://doi.org/10.3390/ani12182416
Chai C, Zhang J, Zhou Y, Yin H, Zhang F, Diao Y, Zan X, Ma Y, Wang Y, Wu Y, et al. The Effects of Oncolytic Pseudorabies Virus Vaccine Strain Inhibited the Growth of Colorectal Cancer HCT-8 Cells In Vitro and In Vivo. Animals. 2022; 12(18):2416. https://doi.org/10.3390/ani12182416
Chicago/Turabian StyleChai, Chunxia, Jinlong Zhang, Yanyan Zhou, Hua Yin, Fan Zhang, Yun Diao, Xiaohui Zan, Yanhua Ma, Yan Wang, Youzhi Wu, and et al. 2022. "The Effects of Oncolytic Pseudorabies Virus Vaccine Strain Inhibited the Growth of Colorectal Cancer HCT-8 Cells In Vitro and In Vivo" Animals 12, no. 18: 2416. https://doi.org/10.3390/ani12182416