
The Effect of Different Extenders on the Quality Characteristics of European Red Deer Epididymal Sperm Stored at 5 °C
 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sperm Samples
2.2. Procedure for Storing Spermatozoa in A Liquid State
2.3. Evaluation of Epididymal Sperm Quality Characteristics
2.3.1. Assessment of Sperm Motility and Motility Parameters by the CASA System
2.3.2. Plasma Membrane Integrity
2.3.3. Acrosomal Membrane Integrity
2.3.4. Mitochondrial Activity
2.3.5. DNA Integrity
2.3.6. Viability and Apoptotic-like Changes in the Plasma Membrane
2.4. Statistical Analysis
3. Results
3.1. The Effect of Different Extenders and Storage Time on Sperm Motility and Motility Parameters
3.2. The Effect of the Analyzed Extenders on Membrane Integrity, Mitochondrial Activity, and DNA Integrity
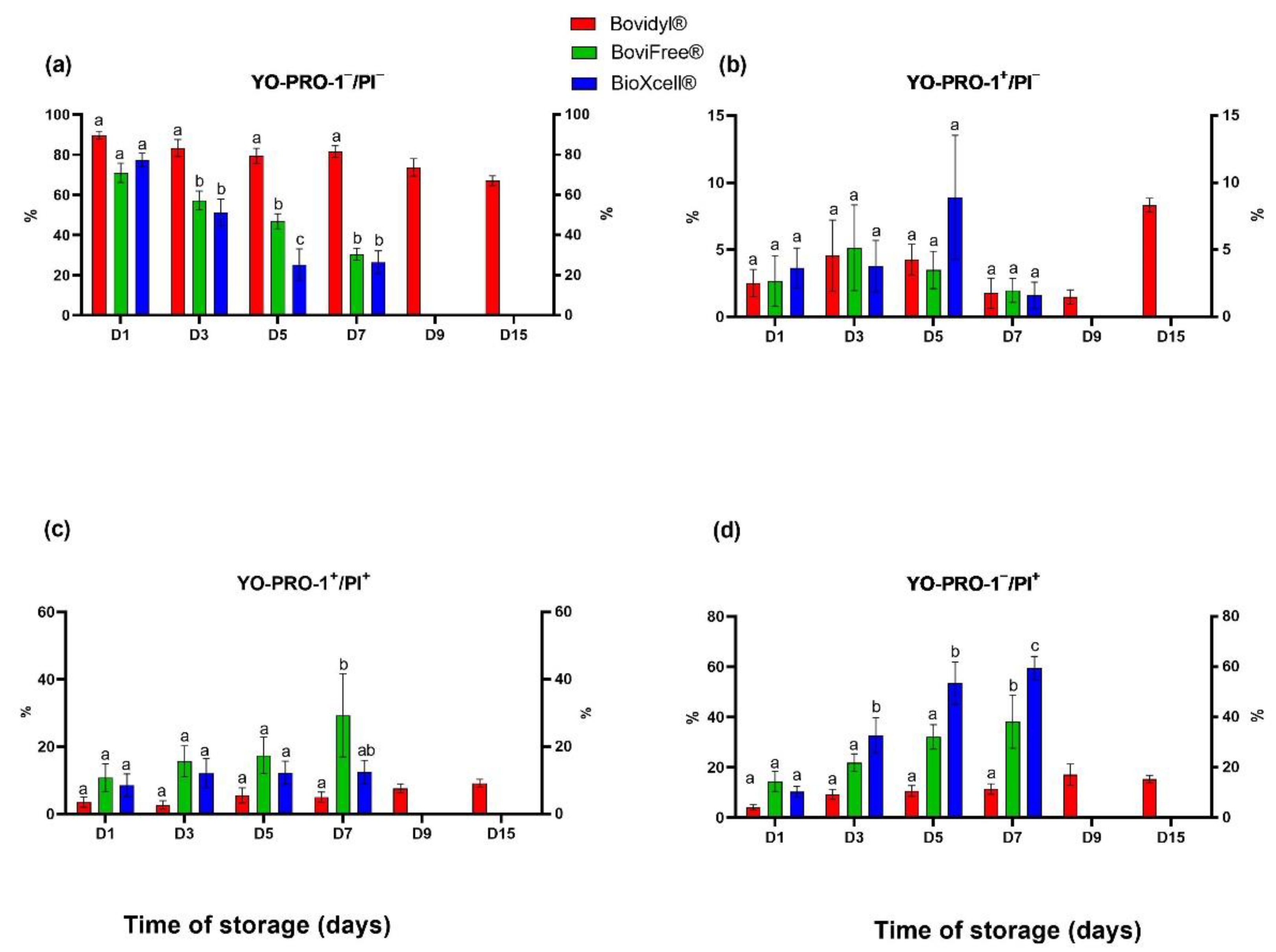
3.3. The Effect of the Analyzed Extenders and Storage Time on Sperm Viability and Apoptotic-like Changes in the Plasma Membrane
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Comizzoli, P.; Mermillod, P.; Mauget, R. Reproductive biotechnologies for endangered mammalian species. Reprod. Nutr. Dev. 2000, 40, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Faigl, V.; Vass, N.; Jávor, A.; Kulcsár, M.; Solti, L.; Amiridis, G.; Cseh, S. Artificial insemination of small ruminants-a review. Acta Vet. Hung. 2012, 60, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.W.; Berg, D.K.; Evans, G. Storage of semen and artificial insemination in deer. Anim. Reprod. Sci. 2000, 62, 195–211. [Google Scholar] [CrossRef]
- Garde, J.J.; Martínez-Pastor, F.; Gomendio, M.; Malo, A.F.; Soler, A.J.; Fernández-Santos, M.R.; Esteso, M.C.; García, A.J.; Anel, L.; Roldán, E.R.S. The application of reproductive technologies to natural populations of red deer. Reprod. Domest. Anim. 2006, 41, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Aller, J.F.; Fernandez, O.; Sanchez, E. Fixed-time artificial insemination in red deer (Cervus elaphus) in Argentina. Anim. Reprod. Sci. 2009, 115, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Macias, V.; Martinez-Pastor, F.; Alvarez, M.; Garde, J.J.; Anel, E.; Anel, L.; de Paz, P. Assessment of chromatin status (SCSA) in epididymal and ejaculated sperm in Iberian red deer, ram and domestic dog. Theriogenology 2006, 66, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Gloria, A.; Contri, A.; De Amicis, I.; Robbe, D.; Carluccio, A. Differences between epididymal and ejaculated sperm characteristics in donkey. Anim. Reprod. Sci. 2011, 128, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Abella, D.F.; Da Costa, M.; Guérin, Y.; Dacheux, J.L. Fertility of undiluted ram epididymal spermatozoa stored for several days at 4 °C. Animal 2015, 9, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Santos, M.R.; Esteso, M.C.; Soler, A.J.; Montoro, V.; Garde, J.J. Effects of egg yolk and cooling rate on the survival of refrigerated red deer (Cervus elaphus hispanicus) epididymal spermatozoa. Reprod. Domest. Anim. 2006, 41, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Medina-Chávez, D.A.; Soler, A.J.; Martín-Maestro, A.; Villaverde, S.; Sánchez-Ajofrín, I.; Peris-Frau, P.; Del Olmo, E.; Bisbal, A.; OGarcía-Álvarez, O.; Fernández-Santos, M.D.R.; et al. Freezing Protocol Optimization for Iberian Red Deer (Cervus elaphus hispanicus) Epididymal Sperm under Field Conditions. Animals 2022, 12, 869. [Google Scholar] [CrossRef]
- Malcotti, V.; Pelufo, V.; Bergamo, N.; Aisen, E. Recovery of epididymal spermatozoa from bull and red deer, stored at different times and temperatures before freezing–thawing. Anim. Prod. Sci. 2012, 52, 741–745. [Google Scholar] [CrossRef]
- Wusiman, A.; Wang, Y.P.; Ren, K.; Zhou, G.B.; Fu, X.W.; Suo, L.; Fan, Z.Q.; Wang, L.; Zhu, S.E. Semen Storage at 23, 4 or −196 °C and Its Applications to Artificial Insemination in Small-Tail Han Sheep. Asian J. Anim. Vet. Adv. 2012, 7, 299–308. [Google Scholar] [CrossRef]
- Fernandez-Santos, M.R.; Dominguez-Rebolledo, A.E.; Esteso, M.C.; Garde, J.J.; Martinez-Pastor, F. Refrigerated storage of red deer epididymal spermatozoa in the epididymis, diluted and with vitamin C supplementation. Reprod. Domest. Anim. 2009, 44, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Dziekońska, A.; Niedźwiecka, E.; Niklewska, M.E.; Koziorowska-Gilun, M.; Kordan, W. Viability longevity and quality of epididymal sperm stored in the liquid state of European red deer (Cervus elaphus elaphus). Anim. Reprod. Sci. 2020, 213, 106269. [Google Scholar] [CrossRef]
- Fernandez-Novo, A.; Santos-Lopez, S.; Barrajon-Masa, C.; Mozas, P.; de Mercado, E.; Caceres, E.; Garrafa, A.; Gonzalez-Martin, J.V.; Perez-Villalobos, N.; Oliet, A.; et al. Effects of Extender Type, Storage Time, and Temperature on Bull Semen Parameters. Biology 2021, 10, 630. [Google Scholar] [CrossRef]
- Martinez-Pastor, F.; Martinez, F.; Alvarez, M.; Maroto-Morales, A.; Garcia-Alvarez, O.; Soler, A.J.; Garde, J.J.; de Paz, P.; Anel, L. Cryopreservation of Iberian red deer (Cervus elaphus hispanicus) spermatozoa obtained by electroejaculation. Theriogenology 2009, 71, 628–638. [Google Scholar] [CrossRef]
- Cheng, F.P.; Wu, J.T.; Chan, J.P.; Wang, J.S.; Fung, H.P.; Colenbrander, B.; Tung, K.C. The effect of different extenders on post-thaw sperm survival, acrosomal integrity and longevity in cryopreserved semen of Formosan Sika deer and Formosan Sambar deer. Theriogenology 2004, 61, 1605–1616. [Google Scholar] [CrossRef]
- Giżejewski, Z. Effect of season on characteristics of red deer/Cervus elaphus L./semen collected using modified artificial vagina. Reprod. Biol. 2004, 4, 51–66. [Google Scholar]
- Garner, D.L.; Johnson, L.A. Viability assessment of mammalian sperm using Sybr-14 and propidium iodide. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Lange-Consiglio, A.; Meucci, A.; Cremonesi, F. Fluorescent multiple staining and CASA system to assess boar sperm viability and membranes integrity in short and long-term extenders. Open Vet. J. 2013, 3, 21–35. [Google Scholar]
- Partyka, A.; Rodak, O.; Bajzert, J.; Kochan, J.; Niżański, W. The Effect of L-Carnitine, Hypotaurine, and Taurine Supplementation on the Quality of Cryopreserved Chicken Semen. Biomed. Res. Int. 2017, 2017, 7279341. [Google Scholar] [CrossRef] [PubMed]
- Trzcińska, M.; Bryła, M.; Smorąg, Z. Apoptotic-like changes in the spermatozoa of fresh and stored boar semen and the quality of embryos produced In Vivo. Anim. Reprod. Sci. 2011, 124, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Moce, E.; Graham, J.K. In Vitro evaluation of sperm quality. Anim. Reprod. Sci. 2008, 105, 104–118. [Google Scholar] [CrossRef]
- Dott, H.M.; Foster, G.C.A. The estimation of sperm motility in semen on membrane slide by measuring the area change frequency with inn image analysing computer. J. Reprod. Fertil. 1979, 55, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Ramón, M.; Martínez-Pastor, F.; García-Álvarez, O.; Maroto-Morales, A.; Soler, A.J.; Jiménez-Rabadán, P.; Fernández-Santos, M.R.; Bernabéu, R.; Garde, J.J. Taking advantage of the use of supervised learning methods for characterization of sperm population structure related with freezability in the Iberian red deer. Theriogenology 2012, 77, 1661–1672. [Google Scholar] [CrossRef]
- Akbarinejad, V.; Fathi, R.; Shahverdi, A.; Esmaeili, V.; Rezagholizadeh, A.; Ghaleno, L.R. The Relationship of Mitochondrial Membrane Potential, Reactive Oxygen Species, Adenosine Triphosphate Content, Sperm Plasma Membrane Integrity, and Kinematic Properties in Warmblood Stallions. J. Equine Vet. Sci. 2020, 94, 103267. [Google Scholar] [CrossRef]
- Hirohashi, N.; Yanagimachi, R. Sperm acrosome reaction: Its site and role in fertilization. Biol. Reprod. 2018, 99, 127–133. [Google Scholar] [CrossRef]
- Andrade, A.F.C.; Arruda, R.P.; Celeghini, E.C.C.; Nascimento, J.; Martins, S.M.M.K.; Raphael, C.F.; Moretti, A.S. Fluorescent stain method for the simultaneous determination of mitochondrial potential and integrity of plasma and acrosomal membranes in boar sperm. Reprod. Domest. Anim. 2007, 42, 190–194. [Google Scholar] [CrossRef]
- Yanagimachi, R. Mammalian fertilization. In The Physiology of Reproduction, 2nd ed.; Knobil, E., Neil, J.D., Eds.; Raven Press: New York, NY, USA, 1994; Volume 1, pp. 189–317. [Google Scholar]
- Barbagallo, F.; La Vignera, S.; Cannarella, R.; Aversa, A.; Calogero, A.E.; Condorelli, R.A. Evaluation of Sperm Mitochondrial Function: A Key Organelle for Sperm Motility. J. Clin. Med. 2020, 9, 363. [Google Scholar] [CrossRef]
- Travis, A.J.; Foster, J.A.; Rosenbaum, N.A.; Visconti, P.E.; Gerton, G.L.; Kopf, G.S.; Moss, S.B. Targeting of a germ cell-specific type 1 hexokinase lacking a porin-binding domain to the mitochondria as well as to the head and fibrous sheath of murine spermatozoa. Mol. Biol. Cell 1998, 9, 263–276. [Google Scholar] [CrossRef][Green Version]
- Gravance, C.; Garner, D.; Baumber, J.; Ball, B. Assessment of equine sperm mitochondrial function using JC-1. Theriogenology 2000, 53, 1691–1703. [Google Scholar] [CrossRef]
- Marchetti, C.; Jouy, N.; Leroy-Martin, B.; Defossez, A.; Formstecher, P.; Marchetti, P. Comparison of four fluorochromes for the detection of the inner mitochondrial membrane potential in human spermatozoa and their correlation with sperm motility. Hum. Reprod. 2004, 19, 2267–2276. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, Y.; Zhou, T.; Zhang, R.; Chen, H.; Chen, S.; Zhao, H. Mechanisms of ROS-induced mitochondria-dependent apoptosis underlying liquid storage of goat spermatozoa. Aging 2019, 11, 7880–7898. [Google Scholar] [CrossRef]
- Kumaresan, A.; Das Gupta, M.; Datta, T.K.; Morrell, J.M. Sperm DNA Integrity and Male Fertility in Farm Animals: A Review. Front. Vet. Sci. 2020, 7, 321. [Google Scholar] [CrossRef]
- Sadeghi, M.R.; Hodjat, M.; Lakpour, N.; Arefi, S.; Amirjannati, N.; Modarresi, T.; Jadda, H.H.; Akhondi, M.M. Effects of sperm chromatin integrity on fertilization rate and embryo quality following intracytoplasmic sperm injection. Avicenna J. Med. Biotechnol. 2009, 1, 173–180. [Google Scholar] [PubMed]
- Love, C.C.; Brinsko, S.P.; Rigby, S.L.; Thompson, J.A.; Blanchard, T.L.; Varner, D.D. Relationship of seminal plasma level and extender type to sperm motility and DNA integrity. Theriogenology 2005, 63, 1584–1591. [Google Scholar] [CrossRef]
- Kasimanickam, R.; Kasimanickam, V.; Tibary, A.; Pelzer, K. Effect of semen extenders on sperm parameters of ram semen during liquid storage at 4 °C. Small Rumin. 2011, 99, 208–213. [Google Scholar] [CrossRef]
- Fernández-Santos, M.R.; Martínez-Pastor, F.; Matias, D.; Domínguez-Rebolledo, A.E.; Esteso, M.C.; Montoro, V.; Garde, J.J. Effects of long-term chilled storage of red deer epididymides on DNA integrity and motility of thawed spermatozoa. Anim. Reprod. Sci. 2009, 111, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, S.; Del Gallego, R.; García-Colomer, B.; Gómez, E.A.; Yániz, J.L.; Gosálvez, J.; López-Fernández, C.; Silvestre, M.A. Effect of Sperm Concentration and Storage Temperature on Goat Spermatozoa during Liquid Storage. Biology 2020, 9, 300. [Google Scholar] [CrossRef]
- Bielas, W.; Niżański, W.; Partyka, A.; Rząsa, A.; Mordak, R. Effect of long-term storage in Safe Cell+ extender on boar sperm DNA integrity and other key sperm parameters. Acta Vet. Scand. 2017, 59, 58. [Google Scholar] [CrossRef] [PubMed]
- Boe-Hansen, G.B.; Ersbøll, A.K.; Greve, T.; Christensen, P. Increasing storage time of extended boar semen reduces sperm DNA integrity. Theriogenology 2005, 63, 2006–2019. [Google Scholar] [CrossRef]
- Wasilewska, K.; Zasiadczyk, Ł.; Fraser, L.; Mogielnicka-Brzozowska, M.; Kordan, W. The benefits of cooling boar semen in long-term extenders prior to cryopreservation on sperm quality characteristics. Reprod. Domest. Anim. 2016, 51, 781–788. [Google Scholar] [CrossRef]
- Bergeron, A.; Crete, M.H.; Brindle, Y.; Manjunath, P. Low-density lipoprotein fraction from hen’s egg yolk decreases the binding of the major proteins of bovine seminal plasma to sperm and prevents lipid efflux from the sperm membrane. Biol. Reprod. 2004, 70, 708–717. [Google Scholar] [CrossRef]
- Watson, P.F. The roles of lipid and protein in the protection of ram spermatozoa at 5 degrees C by egg-yolk lipoprotein. J. Reprod. Fertil. 1981, 62, 483–492. [Google Scholar] [CrossRef]
- Zarei, M.; Rostami, B.; Masoumi, R.; Sharafi, M.; Ahahir, M.H.; Steer, M.; Catt, S. Egg yolk enriched with polyunsaturated fatty acids (PUFAs) improves the shelf life of ram semen in liquid storage. Small Rumin. 2018, 166, 87–92. [Google Scholar] [CrossRef]
- Dziekońska, A.; Behrendt, D.; Strzeżek, R.; Kordan, W. Effect of ostrich egg lipoproteins and hen egg yolk on the quality of dog sperm during liquid storage at 5 °C. Ann. Anim. Sci. 2018, 18, 113–124. [Google Scholar] [CrossRef]
- de Ruigh, L.; Bosch, J.C.; Brus, M.C.; Landman, B.; Merton, J.S. Ways to improve the biosecurity of bovine semen. Reprod. Domest. Anim. 2006, 41, 268–274. [Google Scholar] [CrossRef]
- Ros-Santaella, J.L.; Pintus, E. Plant Extracts as Alternative Additives for Sperm Preservation. Antioxidants 2021, 10, 772. [Google Scholar] [CrossRef] [PubMed]
- Crespilho, O.M.; Sa Filho, M.F.; Dell’Aqua Jr, J.A.; Nichi, M.; Monteiro, G.A.; Avanzi, B.R.; Martins, A.; Papa, F.O. Comparison of in vitro and in vivo fertlizing potential of bovine semen frozen in egg yolk or new lecithin based extenders. Livest. Sci. 2012, 149, 1–6. [Google Scholar] [CrossRef]
- Nguyen Vui, V.; Ponchunchoovong, S.; Kupittayanant, S.; Kupittayanant, P. Effects of egg yolk soybean lecitjin on sperm quality determined by computer-assisted sperm analysis and confocal laser scanning microscope in chilled canine sperm. Vet. Med.Sci. 2019, 5, 345–360. [Google Scholar] [CrossRef]
- Pytlík, J.; Savvulidi, F.G.; Ducháček, J.; Codl, R.; Vrhel, M.; Nagy, S.; Stádník, L. Effect of extender on the quality and incubation resilience of cryopreserved Holstein bull semen. Czech J. Anim. Sci. 2022, 67, 75–86. [Google Scholar] [CrossRef]
- Silva, E.C.B.; Cajueiro, J.F.P.; Silva, S.V.; Vidal, A.H.; Soares, P.C.; Guerra, M.M.P. In vitro evaluation of ram sperm frozen with glycerol, ethylene glycol or acetamide. Anim. Reprod. Sci. 2012, 132, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Santos, M.R.; Esteso, M.C.; Montoro, V.; Soler, A.J.; Garde, J.J. Cryopreservation of Iberian red deer (Cervus elaphus hispanicus) epididymal spermatozoa: Effects of egg yolk, glycerol and cooling rate. Theriogenology 2006, 66, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Domingo, P.; Olaciregui, M.; González, N.; De Blas, I.; Gil, L. Effect of glycerol, n, n-dimethylformamide and n-methyl-2-pyrrolidone on rabbit sperm stored at 4 °C and 16 °C. Anim. Reprod. 2019, 16, 887–894. [Google Scholar] [CrossRef]
- Macías García, B.; Ortega Ferrusola, C.; Aparicio, I.M.; Miró-Morán, A.; Morillo Rodriguez, A.; Gallardo Bolaños, J.M.; González Fernández, L.; Balao da Silva, C.M.; Rodríguez Martínez, H.; Tapia, J.A.; et al. Toxicity of glycerol for the stallion spermatozoa: Effects on membrane integrity and cytoskeleton, lipid peroxidation and mitochondrial membrane potential. Theriogenology 2012, 77, 1280–1289. [Google Scholar] [CrossRef]
- Gilmore, J.A.; McGann, L.E.; Liu, J.; Gao, D.Y.; Peter, A.T.; Kleinhans, F.W.; Critser, J.K. Effect of cryoprotectant solutes on water permeability of human spermatozoa. Biol. Reprod. 1995, 53, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Okuda, Y.; Seita, Y.; Hisamatsu, S.; Sonoki, S.; Shino, M.; Masaoka, T.; Inomata, T.; Kamijo, S.; Kashiwazaki, N. Fertility of spermatozoa cryopreserved with 2% acetamide or glycerol through artificial insemination in the Japanese white rabbit. Exp. Anim. 2007, 56, 29–34. [Google Scholar] [CrossRef] [PubMed][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | Extender | Time of Storage | Extender × Time of Storage | |||
|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| TMOT | 54.230 | 0.001 | 40.373 | 0.001 | 5.799 | 0.001 |
| PMOT | 55.854 | 0.001 | 38.605 | 0.001 | 6.544 | 0.001 |
| VAP | 35.840 | 0.001 | 32.735 | 0.001 | 2.381 | 0.034 |
| VSL | 46.555 | 0.001 | 41.351 | 0.001 | 4.133 | 0.001 |
| VCL | 18.774 | 0.001 | 16.350 | 0.001 | 2.093 | n.s. |
| ALH | 14.535 | 0.001 | 3.168 | 0.028 | 1.568 | n.s. |
| BCF | 13.694 | 0.001 | 13.995 | 0.001 | 6.651 | 0.001 |
| LIN | 15.264 | 0.001 | 7.668 | 0.001 | 3.362 | 0.005 |
| STR | 5.004 | 0.001 | 5.157 | 0.002 | 3.357 | 0.005 |
| SYBR-14+/PI− | 113.853 | 0.001 | 41.861 | 0.001 | 11.250 | 0.001 |
| FITC-PNA−/PI− | 114.865 | 0.001 | 29.866 | 0.001 | 6.622 | 0.001 |
| DNA integrity | 1.800 | n.s. | 2.060 | n.s. | 2.560 | 0.024 |
| MMP | 124.589 | 0.001 | 28.370 | 0.001 | 7.828 | 0.001 |
| YO-PRO-1−/PI− | 76.867 | 0.001 | 29.398 | 0.001 | 5.733 | 0.001 |
| YO-PRO-1+/PI− | 0.291 | n.s. | 1.386 | n.s. | 0.541 | n.s. |
| YO-PRO-1+/PI+ | 9.892 | 0.001 | 1.406 | n.s. | 0.728 | n.s. |
| YO-PRO-1−/PI+ | 35.130 | 0.001 | 16.344 | 0.001 | 3.633 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziekońska, A.; Neuman, N.M.; Burdal, K.K.; Wiszniewska-Łaszczych, A.; Bogdaszewski, M. The Effect of Different Extenders on the Quality Characteristics of European Red Deer Epididymal Sperm Stored at 5 °C. Animals 2022, 12, 2669. https://doi.org/10.3390/ani12192669
Dziekońska A, Neuman NM, Burdal KK, Wiszniewska-Łaszczych A, Bogdaszewski M. The Effect of Different Extenders on the Quality Characteristics of European Red Deer Epididymal Sperm Stored at 5 °C. Animals. 2022; 12(19):2669. https://doi.org/10.3390/ani12192669
Chicago/Turabian StyleDziekońska, Anna, Nicoletta M. Neuman, Klaudia K. Burdal, Agnieszka Wiszniewska-Łaszczych, and Marek Bogdaszewski. 2022. "The Effect of Different Extenders on the Quality Characteristics of European Red Deer Epididymal Sperm Stored at 5 °C" Animals 12, no. 19: 2669. https://doi.org/10.3390/ani12192669
APA StyleDziekońska, A., Neuman, N. M., Burdal, K. K., Wiszniewska-Łaszczych, A., & Bogdaszewski, M. (2022). The Effect of Different Extenders on the Quality Characteristics of European Red Deer Epididymal Sperm Stored at 5 °C. Animals, 12(19), 2669. https://doi.org/10.3390/ani12192669