
Phytoestrogens: A Review of Their Impacts on Reproductive Physiology and Other Effects upon Grazing Livestock
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Phytoestrogens
2.1. Isoflavones
2.2. Coumestans
2.3. Lignans
3. Metabolism and Metabolic Effects
4. Effects on Antioxidant and Immune Capacity
5. Effects on Growth and Performance
6. Effects on Female Reproductive Physiology
6.1. Cows
6.2. Ewes
6.3. Mares
7. Effects on Male Reproductive Physiology
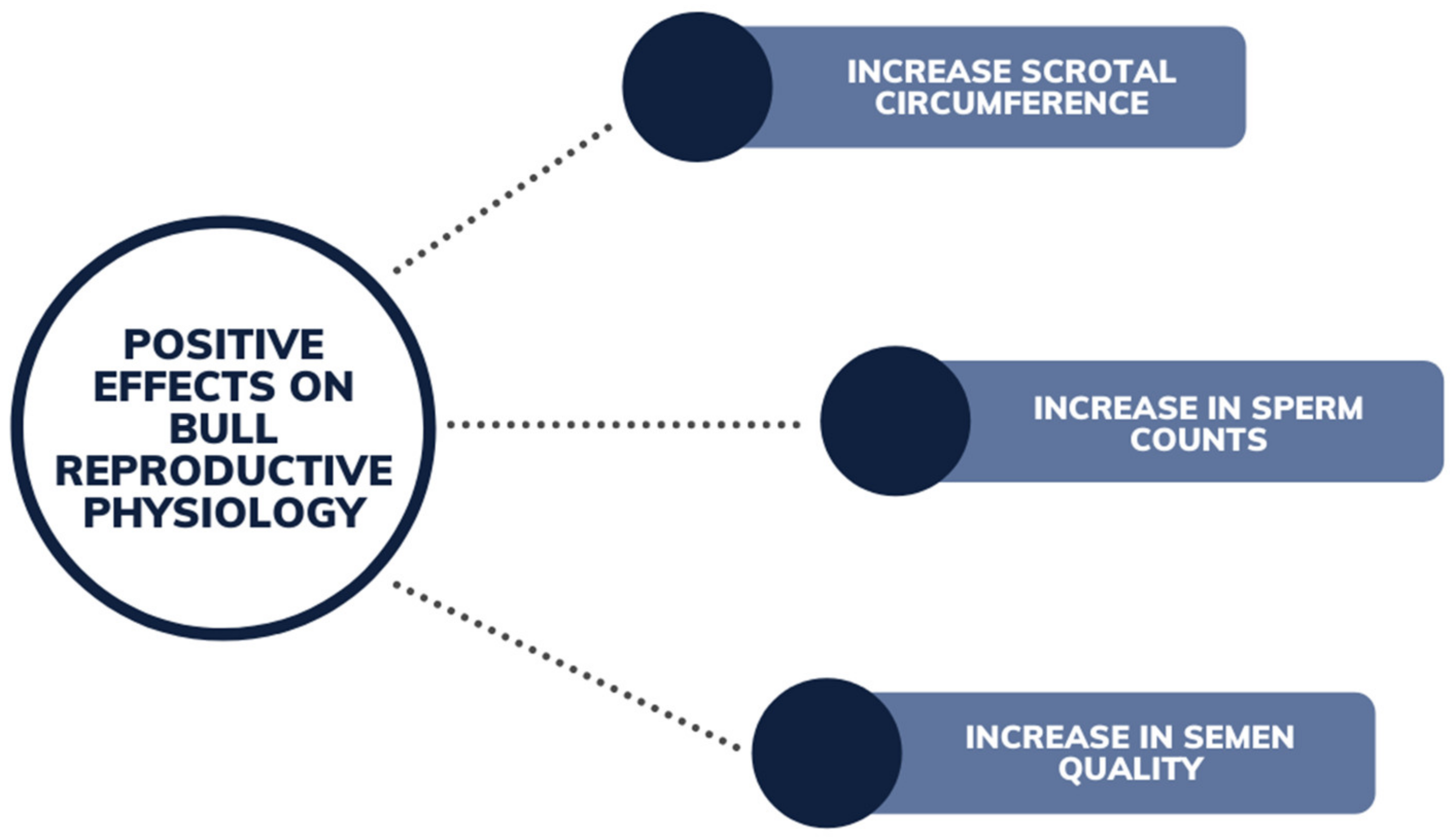
7.1. Bulls and Steers
7.2. Rams and Wethers
7.3. Stallions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kulkarni, K.P.; Tayade, R.; Asekova, S.; Song, J.T.; Shannon, J.G.; Lee, J.D. Harnessing the potential of forage legumes, alfalfa, soybean, and cowpea for sustainable agriculture and global food security. Front. Plant Sci. 2018, 9, 1314. [Google Scholar] [CrossRef] [PubMed]
- Melchior, E.A.; Myer, P.R. Fescue toxicosis and its influence on the rumen microbiome: Mitigation of production losses through clover isoflavones. J. Appl. Anim. Res. 2018, 46, 1280–1288. [Google Scholar] [CrossRef] [Green Version]
- Pannell, D.J. Economic aspects of legume management and legume research in dryland farming systems of southern Australia. Agric. Syst. 1995, 49, 217–236. [Google Scholar] [CrossRef]
- Hudson, D. Towards a Sustainable Australian Temperate Pasture Legume Planting Seed Market; The Australian Lucerne Seed Industry, Rural Industries Research and Development Corporation: Canberra, Australia, 2017. [Google Scholar]
- MLA. Fast Facts-Australia’s Beef Industry. In Market Information; MLA: North Sydney, NSW, Australia, 2020; p. 2. [Google Scholar]
- Jones, F.M.; Accioly, J.M.; Copping, K.J.; Deland, M.P.B.; Graham, J.F.; Hebart, M.L.; Herd, R.M.; Laurence, M.; Lee, S.J.; Speijers, E.J. Divergent breeding values for fatness or residual feed intake in Angus cattle. 1. Pregnancy rates of heifers differed between fat lines and were affected by weight and fat. Anim. Prod. Sci. 2017, 58, 33–42. [Google Scholar] [CrossRef]
- Kurzer, M.S.; Xu, X. Dietary Phyto-oestrogens. Ann. Rev. Nutr. 1997, 17, 353–381. [Google Scholar] [CrossRef] [PubMed]
- Karki, K.B.; Mishra, A.K.; Choi, S.J.; Baek, K.H. Effect of Ultraviolet C Irradiation on Isoflavone Concentrations in Different Cultivars of Soybean (Glycine max). Plants 2020, 9, 1043. [Google Scholar] [CrossRef]
- Hughes, C.L., Jr. Phytochemical mimicry of reproductive hormones and modulation of herbivore fertility by phytoestrogens. Environ. Health Perspect. 1988, 78, 171–174. [Google Scholar] [CrossRef]
- Dixon, R.A. Phytoestrogens. Annu. Rev. Plant Biol. 2004, 55, 225–261. [Google Scholar] [CrossRef] [Green Version]
- Patisaul, H.B. Infertility in the Southern White Rhino: Is diet the source of the problem? Endocrinology 2012, 153, 1568–1571. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.H.; Wang, S.C.; Hsu, C.; Lin, B.F.; Kuo, Y.H.; Huang, C.J. Phytoestrogenic compounds in alfalfa sprout (Medicago sativa) beyond coumestrol. J. Agric. Food Chem. 2011, 59, 131–137. [Google Scholar] [CrossRef]
- Hashem, N.; Soltan, Y. Impacts of phytoestrogens on livestock production: A review. Egypt. J. Nutr. Feeds 2016, 19, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Trenkle, A.; Burroughs, W. Physiological effects of estrogens in animal feeds with emphasis on growth ruminants. In Nutrition and Drug Interrelations; Hathcock, J.H., Coon, J., Eds.; Academic Press: New York, NY, USA, 1978; pp. 577–611. [Google Scholar]
- Wyse, J.M.; Latif, S.; Gurusinghe, S.; Berntsen, E.D.; Weston, L.A.; Stephen, C.P. Characterization of Phytoestrogens in Medicago sativa L. and Grazing Beef Cattle. Metabolites 2021, 11, 550. [Google Scholar] [CrossRef] [PubMed]
- Mostrom, M.; Evans, T.J. Phytoestrogens. Reprod. Develop. Toxicol. 2011, 707–722. [Google Scholar]
- Cos, P.; De Bruyne, T.; Apers, S.; Berghe, D.V.; Pieters, L.; Vlietinck, A.J. Phytoestrogens: Recent developments. Planta Med. 2003, 69, 589–599. [Google Scholar] [PubMed] [Green Version]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madej, A.; Lundh, T. Risk of adverse effects of phytoestrogens in animal feed. In Bioactive Compounds in Plants–Benefits and Risks for Man and Animals; Bernhoft, A., Ed.; The Norwegian Academy of Science and Letters: Oslo, Norway, 2008; pp. 94–103. [Google Scholar]
- Wocławek-Potocka, I.; Piskula, M.K.; Bah, M.M.; Siemieniuch, M.J.; Korzekwa, A.; Brzezicka, E.; Skarżyński, D.J. Concentrations of isoflavones and their metabolites in the blood of pregnant and non-pregnant heifers fed soy bean. J. Reprod. Dev. 2008, 54, 358–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shemesh, M.; Shore, L. Effects of environmental estrogens on reproductive parameters in domestic animals. Isr. J. Vet. Med. 2012, 67, 6–10. [Google Scholar]
- Bickoff, E.M.; Spencer, R.R.; Witt, S.C.; Knuckles, B.E. Studies on the Chemical and Biological Properties of Coumestrol and Related Compounds; Agricultural Research Service, Technical Bulletin No. 1408; United States Department of Agriculture: Washington, DC, USA, 1969; pp. 1–95.
- Bickoff, E.M.; Booth, A.N.; Lyman, R.L.; Livingston, A.L.; Thompson, C.R.; Deeds, F. Coumestrol, a new estrogen isolated from forage crops. Science 1957, 126, 969–970. [Google Scholar] [CrossRef]
- Bickoff, E.M.; Spencer, R.R.; Knuckles, B.E.; Lundin, R.E. 3′-Methoxycoumestrol from alfalfa: Isolation and characterization. J. Agric. Food Chem. 1966, 14, 444–446. [Google Scholar] [CrossRef]
- Seguin, P.; Zheng, W. Phytoestrogen content of alfalfa varieties grown in eastern Canada. J. Sci. Food Agric. 2006, 86, 765–771. [Google Scholar] [CrossRef]
- Francis, C.M.; Millington, A.J. The presence of methylated coumestans in annual Medicago species: Response to a fungal pathogen. Aust. J. Agric. Res. 1971, 22, 75–80. [Google Scholar] [CrossRef]
- Kelly, R.W.; Adams, N.R.; Lindsay, D.R. Effect of coumestans on reproduction in the ewe. Aust. J. Agric. Res. 1976, 27, 253–259. [Google Scholar] [CrossRef]
- Sirtori, C.R.; Arnoldi, A.; Johnson, S.K. Phyto-oestrogens: End of a tale? Ann. Med. 2005, 37, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M.; Martin, M. Biosynthesis of pterocarpan, isoflavan and coumestan metabolites of Medicago sativa: Chalcone, isoflavone and isoflavanone precursors. Phytochemistry 1979, 18, 597–602. [Google Scholar] [CrossRef]
- Wong, E. Detection and estimation of oestrogenic constituents in red clover. J. Sci. Food Agric. 1962, 13, 304–308. [Google Scholar] [CrossRef]
- Ferreira-Dias, G.; Botelho, M.; Zagrajczuk, A.; Rebordão, M.R.; Galvão, A.M.; Pinto Bravo, P.; Piotrowska-Tomala, K.; Szóstek, A.Z.; Wiczkowski, W.; Piskula, M.; et al. Coumestrol and its metabolite in mares’ plasma after ingestion of phytoestrogen-rich plants: Potent endocrine disruptors inducing infertility. Theriogenology 2013, 80, 684–692. [Google Scholar] [CrossRef]
- Reed, K. Fertility of herbivores consuming phytoestrogen-containing Medicago and Trifolium species. Agriculture 2016, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Wong, E.; Latch, G.C.M. Coumestans in diseased white clover. Phytochemistry 1971, 10, 466–468. [Google Scholar] [CrossRef]
- Willför, S.M.; Smeds, A.L.; Holmbom, B.R. Chromatographic analysis of lignans. J. Chromatogr. A 2006, 1112, 64–77. [Google Scholar] [CrossRef]
- Saleem, M.; Kim, H.J.; Ali, M.S.; Lee, Y.S. An update on bioactive plant lignans. Nat. Prod. Rep. 2005, 22, 696–716. [Google Scholar] [CrossRef]
- Brito, A.F.; Zang, Y. A review of lignan metabolism, milk enterolactone concentration, and antioxidant status of dairy cows fed flaxseed. Molecules 2018, 24, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Wang, G.; Han, Z.; Yao, W.; Zhu, W. Metabolism of flaxseed lignans in the rumen and its impact on ruminal metabolism and flora. Anim. Feed Sci. Technol. 2009, 150, 18–26. [Google Scholar] [CrossRef]
- Njåstad, K.M.; Adler, S.A.; Hansen-Møller, J.; Thuen, E.; Gustavsson, A.M.; Steinshamn, H. Gastrointestinal metabolism of phytoestrogens in lactating dairy cows fed silages with different botanical composition. J. Dairy Sci. 2014, 97, 7735–7750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.I. Plant estrogens affecting livestock in Australia. In Effects of Poisonous Plants on Livestock; Keeler, R.F., van Kampen, K.R., James, L.F., Eds.; Academic Press: London, UK, 1978; pp. 451–464. [Google Scholar]
- Třináctý, J.; Křížová, L.; Schulzová, V.; Hajšlová, J.; Hanuš, O. The effect of feeding soybean-derived phytoestogens on their concentration in plasma and milk of lactating dairy cows. Arch. Anim. Nutr. 2009, 63, 219–229. [Google Scholar] [CrossRef]
- Höjer, A.; Adler, S.; Purup, S.; Hansen-Møller, J.; Martinsson, K.; Steinshamn, H.; Gustavsson, A.M. Effects of feeding dairy cows different legume-grass silages on milk phytoestrogens concentration. J. Dairy Sci. 2012, 95, 4526–4540. [Google Scholar] [CrossRef] [Green Version]
- Burton, J.; Wells, M. The effect of phytoestrogens on the female genital tract. J. Clin. Pathol. 2002, 55, 401–407. [Google Scholar] [CrossRef]
- Kelly, R.W.; Lindsay, D.R. Change with length of feeding period in the oestrogenic response to ovariectomized ewes to ingested coumestans. Aust. J. Agric. Res. 1975, 26, 305–311. [Google Scholar] [CrossRef]
- Omede, A.A.; Opara, M.N.; Ezeokeke, C.T.; Okoli, I.C. The significance of phytohormones in animal production. Int. J. Trop. Agric. Food Syst. 2008, 2, 89–104. [Google Scholar] [CrossRef]
- Watson, C.S.; Alyea, R.A.; Jeng, Y.J.; Kochukov, M.Y. Nongenomic actions of low concentration estrogens and xenoestrogens on multiple tissues. Mol. Cell. Endocrinol. 2007, 274, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Matthews, J.; Celius, T.; Halgren, R.; Zacharewski, T. Differential estrogen receptor binding of estrogenic substances: A species comparison. J. Steroid Biochem. Mol. Biol. 2000, 74, 223–234. [Google Scholar] [CrossRef]
- Nehybová, T.; Smarda, J.; Beneš, P. Plant coumestans: Recent advances and future perspectives in cancer therapy. Anti-Cancer Agents Med. Chem. 2014, 14, 1351–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustonen, E.A.; Tuori, M.; Kurki, P.; Isolahti, M.; Taponen, J.; Vanhatalo, A. Variety, time of harvest and conditions during growing season have impact on red clover isoflavone content. Agric. Food Sci. 2018, 27, 102–109. [Google Scholar] [CrossRef]
- Zhao, X.H.; Chen, Z.D.; Zhou, S.; Song, X.Z.; Ouyang, K.H.; Pan, K.; Xu, L.J.; Liu, C.J.; Qu, M.R. Effects of daidzein on performance, serum metabolites, nutrient digestibility, and fecal bacterial community in bull calves. Anim. Feed Sci. Technol. 2017, 225, 87–96. [Google Scholar] [CrossRef]
- Zhao, X.H.; Yang, Z.Q.; Bao, L.B.; Wang, C.Y.; Zhou, S.; Gong, J.M.; Fu, C.B.; Xu, L.J.; Liu, C.J.; Qu, M. Daidzein enhances intramuscular fat deposition and improves meat quality in finishing steers. Exp. Biol. Med. 2015, 240, 1152–1157. [Google Scholar] [CrossRef]
- Liu, D.Y.; He, S.J.; Jin, E.H.; Liu, S.Q.; Tang, Y.G.; Li, S.H.; Zhong, L.T. Effect of daidzein on production performance and serum antioxidative function in late lactation cows under heat stress. Livest. Sci. 2013, 152, 16–20. [Google Scholar] [CrossRef]
- Akula, S.M.; Hurley, D.J.; Wixon, R.L.; Wang, C.; Chase, C.C.L. Effect of genistein on replication of bovine herpesvirus type 1. Am. J. Vet. Res. 2002, 63, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Lecot, S.; Belouzard, S.; Dubuisson, J.; Rouillé, Y. Bovine viral diarrhea virus entry is dependent on clathrin-mediated endocytosis. J. Virol. 2005, 79, 10826–10829. [Google Scholar] [CrossRef] [Green Version]
- Pace, V.; Carbone, K.; Spirito, F.; Iacurto, M.; Terzano, M.G.; Verna, M.; Vincenti, F.; Settineri, D. The effects of subterranean clover phytoestrogens on sheep growth, reproduction and carcass characteristics. Meat Sci. 2006, 74, 616–622. [Google Scholar] [CrossRef]
- Itana, D.D.; Duguma, A. The role and impacts of growth hormones in maximizing animal production-a review. Turk. J. Agric. Food Sci. Technol. 2021, 9, 975–981. [Google Scholar] [CrossRef]
- Dai, S.; Duan, J.; Lu, Y.; Cheng, J.; Ren, J.; Deng, W.; Wu, Y. α-Zearalanol, a phytoestrogen for cardiovascular therapy. Endocrine 2004, 25, 117–119. [Google Scholar] [CrossRef]
- Liang, H.; Xu, L.; Zhao, X.; Bai, J.; Chen, Z.; Zhou, S.; Song, X.; Ouyang, K.; Pan, K.; Lui, C. Effect of daidzein on fermentation parameters and bacterial community of finishing Xianan cattle. Ital. J. Anim. Sci. 2018, 17, 950–958. [Google Scholar] [CrossRef]
- Adams, N.R. Detection of the effects of phyto-oestrogens on sheep and cattle. J. Anim. Sci. 1995, 73, 1509–1515. [Google Scholar] [CrossRef] [PubMed]
- Cederroth, C.R.; Auger, J.; Zimmermann, C.; Eustache, F.; Nef, S. Soy, phyto-oestrogens and male reproductive function: A review. Int. J. Androl. 2010, 33, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic activity, biological effect and bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Shore, L.S.; Rios, C.; Marcus, S.; Bernstein, M.; Shemesh, M. Relationship between peripheral estrogen concentrations at insemination and subsequent fetal loss in cattle. Theriogenology 1998, 50, 101–107. [Google Scholar] [CrossRef]
- Piotrowska, K.K.; Wocławek-Potocka, I.; Bah, M.M.; Piskula, M.K.; Pilawski, W.; Bober, A.; Skarzynski, D.J. Phytoestrogens and their metabolites inhibit the sensitivity of the bovine corpus luteum to luteotropic factors. J. Reprod. Dev. 2005, 52, 33. [Google Scholar]
- Zdunczyk, S.; Piskula, M.; Janowski, T.; Baranski, W.; Ras, M. Concentrations of isoflavones in blood plasma of dairy cows with different incidence of silent heat. Bull. Vet. Inst. Pulawy 2005, 49, 189–191. [Google Scholar]
- Lookhart, G.L. Analysis of coumestrol, a plant estrogen, in animal feeds by high-performance liquid chromatography. J. Agric. Food Chem. 1980, 28, 666–667. [Google Scholar] [CrossRef]
- Puschner, B. Anti-nutritional factors in alfalfa hay. In Proceedings of the National Alfalfa Symposium, Las Vegas, NV, USA, 10–12 December 2000; The California Alfalfa Workgroup and The Alfalfa Council: Las Vegas, NV, USA, 2000; pp. 157–162. [Google Scholar]
- Borzym, E.; Bobowiec, R.; Kosior-Korzecka, U.; Martelli, F.; Burmaczuk, A. Disturbances of cow oocyte maturation by phytoestrogens. Med. Weter 2008, 64, 1107–1111. [Google Scholar]
- Adams, N. Morphological changes in the organs of ewes grazing oestrogenic subterranean clover. Res. Vet. Sci. 1977, 22, 216–221. [Google Scholar] [CrossRef]
- Cantero, A.; Sancha, J.L.; Flores, J.M.; Rodriguez, A.; Gonzalez, T. Histopathological changes in the reproductive organs of Manchego ewes grazing on lucerne. J. Vet. Med. A 1996, 43, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.F.; Jagusch, K.T.; Brunswick, L.F.C.; Kelly, R.W. Coumestans in lucerne and ovulation in ewes. N. Z. J. Agric. Res. 1979, 22, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Nwannenna, A.I.; Lundh, T.; Madej, A.; Fredriksson, G.; Björnhag, G. Clinical changes in ovariectomized ewes exposed to phytoestrogens and 17β-estradiol implants. Proc. Soc. Exp. Biol. Med. 1995, 208, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.L.; Moot, D.J.; Sedcole, J.R.; Barrell, G.K. Recovery of ovulation rate in ewes following their removal from an oestrogenic lucerne forage. Anim. Prod. Sci. 2019, 59, 493–498. [Google Scholar] [CrossRef]
- McDowell, K.; Allman, R.; Henning, J. Phytoestrogens and estrogenic activity in white clover samples from no-loss and high-loss fields during mare reproductive loss syndrome. In Proceedings of the 1st Workshop on Mare Reproductive Loss Syndrome, Lexington, KY, USA, August 2002; Powell, D.G., Troppman, A., Tobin, T., Eds.; Kentucky Agricultural Experiment Station: Lexington, KY, USA, 2002. [Google Scholar]
- Poppenga, R. An overview of fetotoxic agents and their possible role as agents in mare reproductive loss syndrome. In Proceedings of the 1st Workshop on Mare Reproductive Loss Syndrome, Lexington, KY, USA, August 2002; Powell, D.G., Troppman, A., Tobin, T., Eds.; Kentucky Agricultural Experiment Station: Lexington, KY, USA, 2002. [Google Scholar]
- Szóstek, A.Z.; Sadowska, A.; Piotrowska-Tomala, K.K.; Botelho, M.; Fradinho, M.J.; Rebordão, M.R.; Ferreira-Dias, G.M.; Skarzynski, D.J. The effect of coumestrol on progesterone and prostaglandin production in the mare: In vitro and in vivo studies. Biol. Reprod. 2016, 95, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.; Rebordão, M.R.; Galvão, A.M.; Bravo, P.P.; Piotrowska-Tomala, K.; Szóstek, A.Z.; Wiczkowski, W.; Piskula, M.; Skarzynski, D.J.; Fradinho, M.J.; et al. Phytoestrogen coumestrol and its metabolite in mares’ plasma after clover mixed pasture and alfalfa pellets ingestion. In Forages and Grazing in Horse Nutrition, 2nd ed.; Saastamoinen, M., Fradinho, M.J., Santos, A.S., Miraglia, N., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2012; Volume 132, pp. 49–53. [Google Scholar]
- Glover, A.; Assinder, S. Acute exposure of adult male rats to dietary phytoestrogens reduces fecundity and alters epididymal steroid hormone receptor expression. J. Endocrinol. 2006, 189, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pool, K.R.; Kent, T.C.; Blache, D. Oestrogenic metabolite equol negatively impacts the functionality of ram spermatozoa in vitro. Theriogenology 2021, 172, 216–222. [Google Scholar] [CrossRef]
- Miller, H.D. Effects of Isoflavones on the Spermatogenesis of Prepuberal Bovine Bulls. Master’s Thesis, Angelo State University, San Angelo, TX, USA, 2015. [Google Scholar]
- Runyan, C.A.; Yurrita, S.C.; Dickison, J.W.; Salisbury, M.W. Evaluation of age of dam and phytoestrogens on growth and semen quality measures in yearling bulls. J. Anim. Sci. 2018, 96, 62. [Google Scholar] [CrossRef]
- Yurrita, S. WS Effects of dietary phytoestrogens on testicular growth and semen quality characteristics in developing Angus bulls. J. Anim. Sci. 2016, 94, 606. [Google Scholar] [CrossRef]
- Meyer, B.D. Comparison of Serum Steroidal Hormone Concentrations in Buller Steers, Riders, and Uninterested Penmates: Implication for the Etiology of the Buller Steer Syndrome in North American Feedlots. Master’s Thesis, Iowa State University, Ames, IA, USA, 2001. [Google Scholar]
- Menzel, V.A.; Hinsch, E.; Hägele, W.; Hinsch, K.D. Effect of genistein on acrosome reaction and zona pellucida binding independent of protein tyrosine kinase inhibition in bull. Asian J. Androl. 2007, 9, 650–658. [Google Scholar] [CrossRef]
- Hinsch, K.D.; Aires, V.; Hägele, W.; Hinsch, E. In vitro tests for essential sperm functions using the phyto-oestrogen genistein as a test substance. Andrologia 2000, 32, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, M.A.; Vicente-Fiel, S.; Raga, E.; Salvador, I.; Soler, C.; Yániz, J.L. Effect of genistein added to bull semen after thawing on pronuclear and sperm quality. Anim. Reprod. Sci. 2015, 163, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, D.H.; El-Shamy, A.A.; Abdelrazek, H.M.; El-Badry, D.A. Effect of genistein on semen quality, antioxidant capacity, caspase-3 expression and DNA integrity in cryopreserved ram spermatozoa. Small Rumin. Res. 2019, 177, 50–55. [Google Scholar] [CrossRef]
- Macías-García, B.; Guimarães, T.; Lopes, G.; Rocha, A.; González-Fernández, L. Effect of genistein addition to equine sperm freezing extender. J. Hell. Vet. Med. 2015, 66, 241–248. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyse, J.; Latif, S.; Gurusinghe, S.; McCormick, J.; Weston, L.A.; Stephen, C.P. Phytoestrogens: A Review of Their Impacts on Reproductive Physiology and Other Effects upon Grazing Livestock. Animals 2022, 12, 2709. https://doi.org/10.3390/ani12192709
Wyse J, Latif S, Gurusinghe S, McCormick J, Weston LA, Stephen CP. Phytoestrogens: A Review of Their Impacts on Reproductive Physiology and Other Effects upon Grazing Livestock. Animals. 2022; 12(19):2709. https://doi.org/10.3390/ani12192709
Chicago/Turabian StyleWyse, Jessica, Sajid Latif, Saliya Gurusinghe, Jeffrey McCormick, Leslie A. Weston, and Cyril P. Stephen. 2022. "Phytoestrogens: A Review of Their Impacts on Reproductive Physiology and Other Effects upon Grazing Livestock" Animals 12, no. 19: 2709. https://doi.org/10.3390/ani12192709