
Enriching Zoo-Housed Ring-Tailed Lemurs (Lemur catta): Assessing the Influence of Three Types of Environmental Enrichment on Behavior

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Study Sites
2.2. Enrichment Types
2.3. Data Collection
2.4. Data Analysis
3. Results
3.1. Time Budget
3.2. Generalized Linear Models (GLMs)
4. Discussion
4.1. Time Budget
4.2. Environmental Enrichment Effect
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosey, G.R.; Melfi, V.; Pankhurst, S. Zoo Animals: Behaviour, Management and Welfare, 2nd ed.; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Mellor, D.J.; Hunt, S.; Gusset, M. Caring for Wildlife: The World Zoo and Aquarium Animal Welfare Strategy; WAZA Executive Office: Gland, Switzerland, 2015. [Google Scholar]
- Mellen, J.D.; Shepherdson, D.J.; Hutchins, M. The future of environmental enrichment. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institution: Washington, DC, USA, 1998; pp. 329–336. [Google Scholar]
- Young, R.J. Environmental Enrichment for Captive Animals; Blackwell Publishing: Oxford, UK, 2013. [Google Scholar]
- de Azevedo, C.S.; Cipreste, C.F.; Young, R.J. Environmental enrichment: A GAP analysis. Appl. Anim. Behav. Sci. 2007, 102, 329–343. [Google Scholar] [CrossRef]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 five domains model: Including human–animal interactions in assessments of animal welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Basset, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef] [Green Version]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2006, 102, 262–302. [Google Scholar] [CrossRef]
- Ward, S.J.; Melfi, V. Keeper-animal interactions: Differences between the behaviour of zoo animals affect stockmanship. PLoS ONE 2015, 10, e0140237. [Google Scholar] [CrossRef] [PubMed]
- Mellen, J.; MacPhee, M.S. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Mason, G. Stereotypies: A critical review. Anim. Behav. 1991, 41, 1015–1037. [Google Scholar] [CrossRef] [Green Version]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Clark, F.E. Cognitive enrichment and welfare: Current approaches and future directions. Anim. Behav. Cogn. 2017, 4, 52–71. [Google Scholar] [CrossRef] [Green Version]
- Alligood, C.; Leighty, K. Putting the “E” in SPIDER: Evolving Trends in the Evaluation of Environmental Enrichment Efficacy in Zoological Settings. Anim. Behav. Cogn. 2015, 2, 200–217. [Google Scholar] [CrossRef]
- Dishman, D.L.; Thomson, D.M.; Karnovsky, N.J. Does simple feeding enrichment raise activity levels of captive ring-tailed lemurs (Lemur catta)? Appl. Anim. Behav. Sci. 2009, 116, 88–95. [Google Scholar] [CrossRef]
- LaFleur, M.; Gould, L. Lemur catta . IUCN Red List. Threat. Species 2020, e.T11496A115565760. [Google Scholar] [CrossRef]
- Byers, O.; Lees, C.; Wilcken, J.; Schwitzer, C. The One Plan Approach: The philosophy and implementation of CBSG’s approach to integrated species conservation planning. WAZA Mag. 2013, 14, 2–5. [Google Scholar]
- Goodman, S.M.; Rakotoarisoa, S.V.; Wilmé, L. The distribution and biogeography of the ringtailed lemur (Lemur catta) in Madagascar. In Ringtailed Lemur Biology; Jolly, A., Sussman, R.W., Koyama, N., Rasamimanana, H., Eds.; Springer: Boston, MA, USA, 2006. [Google Scholar] [CrossRef]
- Gould, L.; Gabriel, D.N. Wet and dry season diets of the Endangered Lemur catta (ring-tailed lemur) in two mountainous rocky outcrop forest fragments in south-central Madagascar. Afr. J. Ecol. 2015, 53, 320–330. [Google Scholar] [CrossRef]
- Norscia, I.; Palagi, E. Cambridge Studies in Biological and Evolutionary Anthropology: The Missing Lemur Link: An Ancestral Step in the Evolution of Human Behaviour Series Number 74: An Ancestral Step in the Evolution of Human Behaviour; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar] [CrossRef]
- Sauther, M.L.; Sussman, R.W.; Gould, L. The socioecology of the ringtailed lemur: Thirty-five years of research. Evol. Anthropol. 1999, 8, 120–132. [Google Scholar] [CrossRef]
- Donati, G.; Santini, L.; Razafindramanana, J.; Boitani, L.; Borgognini-Tarli, S. (Un-)Expected nocturnal activity in “Diurnal” Lemur catta supports cathemerality as one of the key adaptations of the lemurid radiation. Am. J. Phys. Anthropol. 2013, 150, 99–106. [Google Scholar] [CrossRef]
- LaFleur, M.; Sauther, M.; Cuozzo, F.; Yamashita, N.; Youssouf, I.A.J.; Bender, R. Cathemerality in wild ring-tailed lemurs (Lemur catta) in the spiny forest of Tsimanampetsotsa National Park: Camera trap data and preliminary behavioral observations. Primates 2014, 55, 207–217. [Google Scholar] [CrossRef]
- Jolly, A. Lemur Behaviour: A Madagascar Field Study; University of Chicago Press: Chicago, IL, USA, 1966. [Google Scholar]
- Ganzhorn, J.U. Feeding behavior of Lemur catta and Lemur fulvus. Int. J. Primatol. 1986, 7, 17–30. [Google Scholar] [CrossRef]
- Sauther, M.L. The Effect of Reproductive State, Social Rank and Group Size on Resource Use among Free-Ranging Ringtailed Lemurs (Lemur catta) of Madagascar. Ph.D. Thesis, Washington University, St. Louis, MO, USA, 1992. [Google Scholar]
- Gould, L.; Power, M.L.; Ellwanger, N.; Rambeloarivony, H. Feeding behavior and nutrient intake in spiny forest-dwelling ring-tailed lemurs (Lemur catta) during early gestation and early to mid-lactation periods: Compensating in a harsh environment. Am. J. Phys. Anthropol. 2011, 145, 469–479. [Google Scholar] [CrossRef]
- LaFleur, M.; Sauther, M.L. Seasonal feeding ecology of ring-tailed lemurs: A comparison of spiny and gallery forest habitats. Folia Primatol. 2015, 86, 25–34. [Google Scholar] [CrossRef]
- Britt, A. Encouraging natural feeding behavior in captive-bred black and white ruffed Lemurs (Varecia variegata variegata). Zoo Biol. 1998, 17, 379–392. [Google Scholar] [CrossRef]
- Kerridge, F.J. Environmental enrichment to address behavioral differences between wild and captive black-and-white ruffed lemurs (Varecia variegata). Am. J. Primatol. 2005, 66, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Maloney, M.A.; Meiers, S.T.; White, J.; Romano, M.A. Effects of three Food Enrichment Items on the behavior of black lemurs (Eulemur macaco macaco) and ringtail lemurs (Lemur catta) at the Henson Robinson Zoo, Springfield, Illinois. J. Appl. Anim. Welf. Sci. 2006, 9, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.E.; Shapiro, H.G.; Ehmke, E.E. Behavioral responses of three lemur species to different food enrichment devices. Zoo Boil. 2018, 37, 146–155. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Selecting and Testing Environmental Enrichment in Lemurs. Front. Psychol. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.; Taylor, S.; Montrose, V.T. The effects of olfactory stimulation on the behavior of captive ring-tailed lemurs (Lemur catta). Zoo Biol. 2018, 37, 16–22. [Google Scholar] [CrossRef]
- Vaglio, S.; Kaburu, S.S.K.; Young, C.; Pearce, R.; Smith, S.; Tompkins, B.; Elwell, E.; Fontani, S.; Marliani, G.; Accorsi, P.A. Effects of scent enrichment on behavioural and physiological indicators of stress in zoo primates—A preliminary study on red-ruffed lemurs and Lar gibbons. Am. J. Primatol. 2020, 83, e23247. [Google Scholar] [CrossRef]
- Laméris, D.W.; Verspeek, J.; Depoortere, A.; Plessers, L.; Salas, M. Effects of Enclosure and Environmental Enrichment on the Behaviour of Ring-Tailed Lemurs (Lemur catta). J. Zool. Bot. Gard. 2021, 2, 12. [Google Scholar] [CrossRef]
- Vining, A.Q.; Nunn, C.L.; Samson, D.R. Enriched sleep environments lengthen lemur sleep duration. PLoS ONE 2021, 16, e0253251. [Google Scholar] [CrossRef]
- Lutz, C.K. Stereotypic behavior in nonhuman primates as a model for the human condition. ILAR J. 2014, 55, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, S.L.; Ross, S.R.; Bloomsmith, M.A. Characterizing abnormal behavior in a large population of zoo-housed chimpanzees: Prevalence and potential influencing factors. PeerJ 2016, 4, e2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sclafani, V.; Norscia, I.; Antonacci, D.; Palagi, E. Scratching around mating: Factors affecting anxiety in wild Lemur catta. Primates 2012, 53, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Zannella, A.; Norscia, I.; Stanyon, R.; Palagi, E. Testing yawning hypotheses in wild populations of two strepsirrhine species: Propithecus verreauxi and Lemur catta. Am. J. Primatol. 2015, 77, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Troisi, A. Displacement activities as a behavioral measure of stress in nonhuman primates and human subjects. Stress 2002, 5, 47–54. [Google Scholar] [CrossRef]
- Ward, S.; Sherwen, S. Zoo animals. In Anthrozoology. Human-Animal Interactions in Domesticated and Wild Animals; Hosey, G., Melf, V., Eds.; Oxford University Press: Oxford, UK, 2019; pp. 81–103. [Google Scholar]
- Goodenough, A.E.; McDonald, K.; Moody, K.; Wheeler, C. Are “visitor fefects” overestmated? Behaviour in captive lemurs is mainly driven by co-variaton with time and weather. J. Zoo Aquar. Res. 2019, 7, 59–66. [Google Scholar]
- Hones, P.E.; Marin, C.M. Enrichment and aggression in primates. Neurosci. Biobehav. Rev. 2006, 30, 413–436. [Google Scholar] [CrossRef]
- Pereira, M.E.; Kappeler, P.M. Divergent systems of agonistic behaviour in lemurid primates. Behaviour 1997, 134, 225–274. [Google Scholar] [CrossRef]
- Lehner, P.N. Handbook of Ethological Methods, 2nd ed.; Cambridge University Press: Cambridge, NY, USA, 1998. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [Green Version]
- de Vries, H.; Stevens, J.M.; Vervaecke, H. Measuring and testing the steepness of dominance hierarchies. Anim. Behav. 2006, 71, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Desmarais, B.A.; Harden, J.J. Testing for zero inflation in count models: Bias correction for the Vuong test. Stata J. 2013, 13, 810–835. [Google Scholar] [CrossRef]
- Dobson, A.J. An Introduction to Generalized Linear Models, 2nd ed.; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Nagelkerke, N. A note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Cook, R.D. Testing predictor contributions in sufficient dimension reduction. Ann. Stat. 2004, 32, 1062–1092. [Google Scholar] [CrossRef] [Green Version]
- Barr, D.J.; Levy, R.; Scheepers, C.; Tily, H.J. Random effects structure for confirmatory hypothesis testing: Keep it maximal. J. Mem. Lang. 2013, 68, 255–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretz, F.; Hothorn, T.; Westfall, P. Multiple Comparisons Using R; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.r-project.org/ (accessed on 13 June 2022).
- Sussman, R.W. An Ecological Study of Two Madagascan Primates: Lemur fulvus rufus (Audebert) and Lemur catta (Linnaeus). Ph.D. Thesis, Duke University, Durham, NC, USA, 1972. [Google Scholar]
- Sussman, R.W. Feeding Behavior of Lemur catta and Lemur fulvus. In Primate Ecology: Studies of Feeding and Ranging Behavior in Lemurs, Monkey and Apes; Clutton-Brock, T.H., Ed.; Accademic Press Inc.: New York, NY, USA, 1977; pp. 1–36. [Google Scholar]
- Keith-Lucas, T.; White, F.J.; Keith-Lucas, L.; Vick, L.G. Changes in behavior in free-ranging Lemur catta following release in a natural habitat. Am. J. Primatol. 1999, 47, 15–28. [Google Scholar] [CrossRef]
- Millette, J.B.; Sauther, M.L.; Cuozzo, F.P. Behavioral responses to tooth loss in wild ring-tailed lemurs (Lemur catta) at the Beza Mahafaly Special Reserve, Madagascar. Am. J. Phys. Anthropol. 2009, 140, 120–134. [Google Scholar] [CrossRef]
- Ellwanger, N.; Gould, L. Variations in behavioural patterns between Lemur catta groups living in different forest types: Implications for conservation. Endanger. Species Res. 2011, 14, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Savage, J.A. Home Range and Provisioning Use of Semi-Free -Ranging Ringtailed Lemurs (Lemur catta). Master’s Thesis, Georgia Southern University, Statesboro, GA, USA, 2005. [Google Scholar]
- Carlstead, K. Effects of captivity on the behavior of wild mammals. In Wild Mammals in Captivity: Principles and Techniques; University of Chicago Press: Chicago, IL, USA, 1996; pp. 317–333. [Google Scholar]
- Shepherdson, D. The animal’s perspective: Developing strategies for successful enrichment. In Proceedings of the AZA Annual Conference, Albuquerque, New Mexico, 14–18 September 1997; pp. 486–489. [Google Scholar]
- Gabriel, D.N. Ecological Flexibility in a Disturbed Landscape: An Assessment of the Behavioural and Health Ecology of Ring-Tailed Lemurs (Lemur catta) in Relation to Forest Fragmentation. Ph.D. Thesis, University of Victoria, Victoria, BC, Canada, 2013. [Google Scholar]
- Sauther, M.L. Resource competition in wild populations of ringtailed lemurs (Lemur catta): Implications for female dominance. In Lemur Social Systems and Their Ecological Basis; Kappeler, P.M., Ganzhorn, J.U., Eds.; Springer: Boston, MA, USA, 1993; pp. 135–152. [Google Scholar] [CrossRef]
- Pereira, M. Obsession with agonistic power. In Ringtailed Lemur Biology; Jolly, A., Sussman, R.W., Koyama, N., Rasamimanana, H., Eds.; Springer: Boston, MA, USA, 2006; pp. 245–270. [Google Scholar]
- Taylor, L.; Sussman, R.W. A preliminary study of kinship and social organization in a semi-free-ranging group of Lemur catta. Int. J. Primatol. 1985, 6, 601–614. [Google Scholar] [CrossRef]
- Sommerfeld, R.; Bauert, M.; Hillmann, E.; Stauffacher, M. Feeding enrichment by self-operated food boxes for white-fronted lemurs (Eulemur fulvus albifrons) in the Masoala exhibit of the Zurich Zoo. Zoo Biol. 2006, 25, 145–154. [Google Scholar] [CrossRef]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef]
- Shepherdson, D.; Bemment, N.; Carman, M.; Reynolds, S. Auditory enrichment for Lar gibbons Hylobates lar at London Zoo. Int. Zoo Yearb. 1989, 28, 256–260. [Google Scholar] [CrossRef]
- Brent, L.; Weaver, O. The physiological and behavioral effects of radio music on singly housed baboons. J. Med. Primatol. 1996, 25, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.; Schwandt, M.; Fritz, J.; Roeder, E.; Nelson, C. A stereo music system as environmental enrichment for captive chimpanzees. Lab. Anim. 2003, 32, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Wells, D.L.; Coleman, D.; Challis, M.G. A note on the effect of auditory stimulation on the behaviour and welfare of zoo-housed gorillas. Appl. Anim. Behav. Sci. 2006, 100, 327–332. [Google Scholar] [CrossRef]
- Robbins, L.; Margulis, S.W. The effects of auditory enrichment on gorillas. Zoo Biol. 2014, 33, 197–203. [Google Scholar] [CrossRef]
- Brent, L.; Belik, M. The response of group-housed baboons to three enrichment toys. Lab. Anim. 1997, 31, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Pizzutto, C.S.; Nichi, M.; Sgai, M.G.F.G.; Corrêa, S.H.R.; Viau, P.; Beresca, A.M.; Alvarenga, C.; Barnabé, R.C.; Guimarães, M.A.B.V. Effect of environmental enrichment on behavioral and endocrine aspects of a captive orangutan (Pongo pygmaeus). LPN 2008, 47, 10–14. [Google Scholar]
- Nakamichi, M.; Koyama, N. Social relationships among ring-tailed lemurs (Lemur catta) in two free-ranging troops at Berenty Reserve, Madagascar. Int. J. Primatol. 1997, 18, 73–93. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Year of Birth | Sex | NDS |
|---|---|---|---|
| Milly | 1993 | Female | 3.667 |
| Bekili | 2000 | Female | 2.500 |
| Andribe | 2000 | Female | 2.417 |
| Mandrare | 2001 | Female | 1.167 |
| Ankarana | 2005 | Male | 1.586 |
| Sakalava | 2006 | Female | 3.667 |
| Category 1: Locomotion | Description |
| Locomotion | An individual ambulates on the ground or on a tree. |
| Category 2: Resting | Description |
| Resting | An individual remains inactive |
| Category 3: Foraging | Description |
| Foraging | An individual searches for and eats food with nose over the ground or terminal branch parts. |
| Feeding | An individual eats food provided by keepers. |
| Category 4: Stress-related behaviors | Description |
| Self-grooming | An individual cleans its own fur. |
| Scratching | An individual repeatedly and rapidly moves its hind limb digits over its own fur. |
| Yawning | An individual stretches mouth wide open without vocalizing in non-feeding, non-resting social context. |
| Category 5: Affiliative behaviors | Description |
| Sit in contact | Two or more individuals are sitting in reciprocal contact. |
| Proximity | Two individuals are far apart for a length not exceeding that of an arm. |
| Grooming | An individual cleans another one’s fur with dental comb and/or tongue. This behavior could be made or received. |
| Reciprocal grooming | Two individuals grooming each other. |
| Play | Two or more individuals play together. The most common forms of social play are the “rough and tumble”, which can include, for example, play slaps, play bites etc., and the “play run”, where one subject chases another one. |
| Category 6: Agonistic behaviors | Description |
| Bite | An individual bites another one. |
| Charge | An individual sprints < 5 m toward partner. |
| Cuff | An individual strikes partner (or attempt). |
| Full cuff | Same as cuff but some fur is pulled out. |
| Chase | An individual chases another one. |
| Muzzle | An individual has physical contact with the antagonist, but still not serious. it is used in minor situations. The animal gives a slap to the other with a quick swipe of the muzzle. |
| Stare | An individual widens eyes during mutual gazing with partner. |
| Go towards | An individual walks in a straight line towards the subject involved in the conflict. Often it assumes a gait with almost exaggerated and bold movements, moves with straight and rigid legs, keeps its head and tail very high and straight and continues to stare at the subject towards which it is heading. |
| Jump fight | Two individuals face each other on the ground in a race of jumping on two legs, holding their arms up and out, jumping around each other and trying to scratch, hit and bite the opponent. |
| Dismiss | An individual causes a lower-ranking subject to move from a certain point by a short vocalization. |
| Avoid | When an individual avoids interacting with another one, or when it changes its moving direction or goes far away from the latter. |
| Check scared | An individual who is being pursued stops and turns to look at his pursuer. The subject is in an alert position with his ears straight forward and with his eyes wide open and fixed on his pursuer. |
| Jump away | An individual leaps away from nearby partner (2 m) |
| Run away | An individual who is being chased runs away by running or jumping on branches trying to get as far away from his pursuer as possible. |
| Be displaced | An individual walks away after gazing at approaching conspecific. |
| Glance | An individual gazes rapidly toward and away from the partner. |
| Category 7: Explorative and scent-marking behaviors | Description |
| Skin lick | An individual licks a partner’s palms, soles, eyelids or nostrils. |
| Wrist mark | An individual scars arboreal substrate with carpal spur near antebrachial gland. |
| Wrist to pit | An individual rubs antebrachial gland against axillary gland. |
| Genital mark | An individual holds/rubs genitalia against arboreal substrate. |
| Urine mark | An individual urinates on the substrate with the hindquarters down and the tail raised like a question mark. |
| Anoint tail | An individual passes the ventral surface of the wrists and arms along the tail held between the legs and held erect in front of the back. |
| Wave tail | An individual arches his tail over his head and waves it in the direction of a conspecific to expand the smell, usually made after the anoint tail. |
| Sniff body | An individual places the nose less than 3 cm from the partner’s body, excluding the nose and the genital area. |
| Sniff genital | An individual places his nose less than 3 cm from the genital area and licks it. |
| Category 8: Self-maintenance | Description |
| Sun bathing | An individual sits upright in an area with sunlight, with the belly-side directed towards the sun and arms open. |
| Urinate | An individual eliminates urine without a specific posture but with a lowered tail. |
| Defecate | An individual eliminates faces without a specific posture. |
| Category 9: Abnormal repetitive behaviors | Description |
| Locomotory pacing | An individual walks/runs repeatedly along the same route. |
| Head trough-back | An individual repeatedly tosses its head in a circular motion. |
| Overgroom | An individual cleans itself or another individual excessively, resulting in bald patches of fur. |
| Self-injurious | An individual uses teeth, claws or nails to harm to itself. |
| Category 10: Vigilance | Description |
| Vigilance | An individual observes the surrounding environment while is sitting or standing. |
| Category 11: Inter-specific interactions | Description |
| Human–lemur interaction | An individual pays attention to, approaches, moves away from, etc., visitors, keepers or the observer. |
| Animal–lemur interaction | An individual pays attention to, approaches, moves away from, etc., other animals, such as peacocks and dogs. |
| Category 12: Out of sight | Description |
| Out of sight | An individual is not visible to the observer. |
| Predictors | Estimates | SEM | C.I. | ꭓ2 | p |
| GLMM1 (locomotion) | |||||
| Intercept a | 3.476 | 0.450 | 2.589, 4.363 | a | a |
| Age | −0.048 | 0.015 | −0.077, −0.019 | −3.217 | 0.001 |
| NDS | 0.009 | 0.066 | −0.122, 0.139 | 0.131 | 0.896 |
| Condition_E1 b | 0.362 | 0.311 | −0.252, 0.975 | 1.163 | 0.245 |
| Condition_E2 b | 0.146 | 0.306 | −0.457, 0.750 | 0.479 | 0.632 |
| Condition_E3 b | 0.137 | 0.318 | −0.489, 0.764 | 0.433 | 0.665 |
| Condition_NE b | −0.023 | 0.315 | −0.644, 0.598 | −0.073 | 0.942 |
| Day of week_weekend b | 0.091 | 0.094 | −0.094, 0.277 | 0.971 | 0.331 |
| Ambient temperature | −0.041 | 0.014 | −0.067, −0.014 | −2.965 | 0.003 |
| Condition_E1 × age b | −0.004 | 0.019 | −0.042, 0.034 | −0.214 | 0.830 |
| Condition_E2 × age b | −0.002 | 0.019 | −0.039, 0.035 | −0.087 | 0.931 |
| Condition_E3 × age b | −0.002 | 0.020 | −0.041, 0.036 | −0.109 | 0.913 |
| Condition_NE × age b | 0.007 | 0.019 | −0.031, 0.045 | 0.379 | 0.705 |
| Condition_E1 × NDS b | 0.064 | 0.086 | −0.106, 0.233 | 0.742 | 0.458 |
| Condition_E2 × NDS b | 0.147 | 0.085 | −0.021, 0.315 | 0.730 | 0.084 |
| Condition_E3 × NDS b | 0.095 | 0.088 | −0.078, 0.269 | 1.080 | 0.280 |
| Condition_NE × NDS b | 0.110 | 0.088 | −0.063, 0.283 | 1.252 | 0.210 |
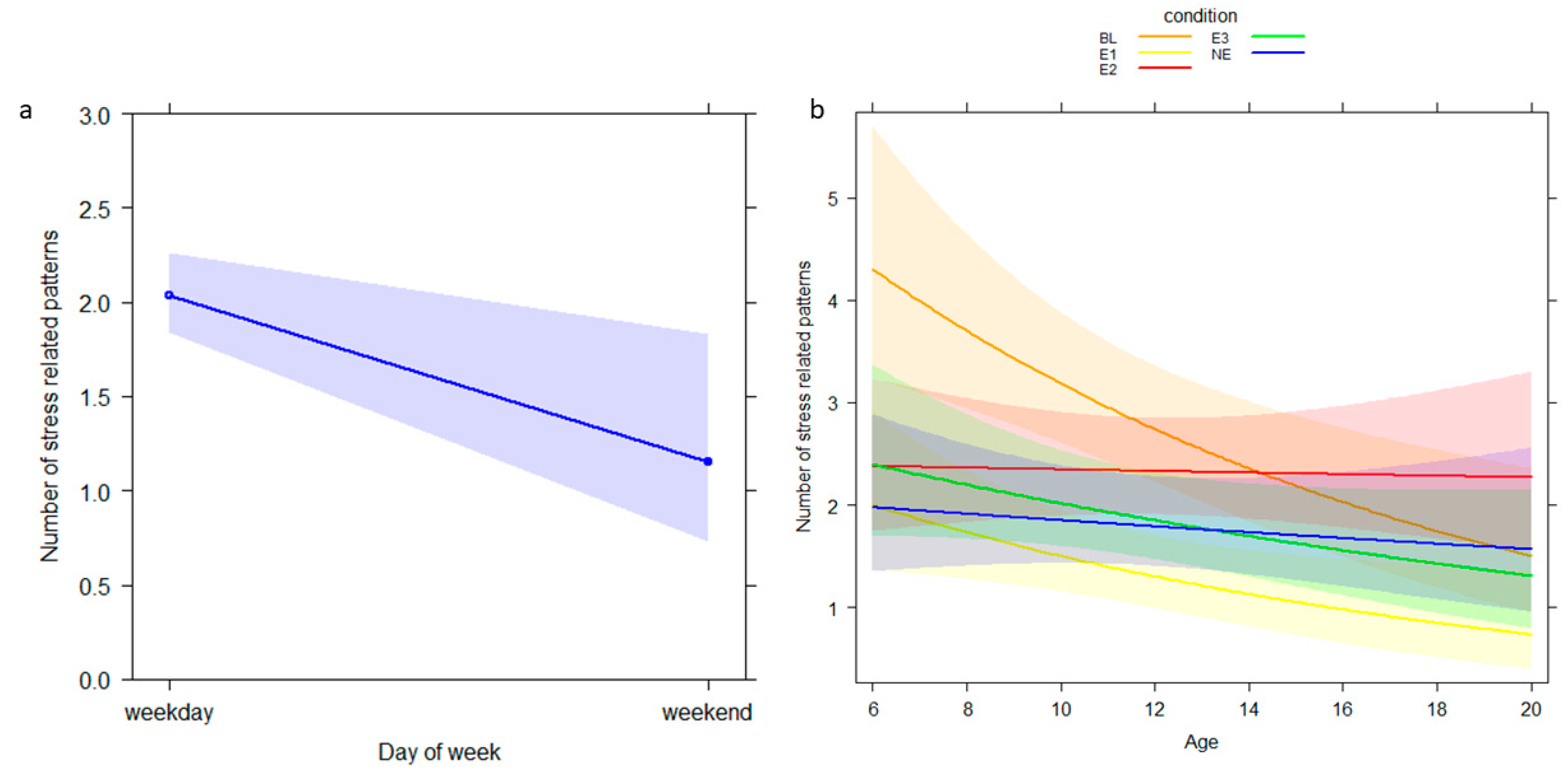
| GLMM2 (stress-related behavior) | |||||
| Intercept a | 0.967 | 0.779 | −0.569, 2.503 | a | a |
| Age | −0.075 | 0.022 | −0.119, −0.031 | −3.387 | 0.001 |
| NDS | 0.037 | 0.095 | −0.151, 0.224 | 0.386 | 0.670 |
| Condition_E1 b | −0.901 | 0.613 | −2.109, 0.308 | −1.470 | 0.142 |
| Condition_E2 b | −1.590 | 0.506 | −2.588, −0.592 | −3.142 | 0.002 |
| Condition_E3 b | −0.904 | 0.553 | −1.994, 0.187 | −1.634 | 0.102 |
| Condition_NE b | −1.369 | 0.566 | −2.485, −0.253 | −2.420 | 0.016 |
| Day of week_weekend b | −0.568 | 0.243 | −1.048, −0.088 | −2.334 | 0.020 |
| Ambient temperature | 0.030 | 0.024 | −0.018, 0.077 | 1.231 | 0.218 |
| Condition_E1 × age b | 0.004 | 0.037 | −0.070, 0.078 | 0.102 | 0.919 |
| Condition_E2 × age b | 0.072 | 0.030 | 0.013, 0.131 | 2.395 | 0.017 |
| Condition_E3 × age b | 0.032 | 0.034 | −0.035, 0.099 | 0.944 | 0.345 |
| Condition_NE × age b | 0.059 | 0.034 | −0.009, 0.126 | 1.704 | 0.088 |
| Condition_E1 × NDS b | 0.045 | 0.163 | −0.277, 0.367 | 0.274 | 0.784 |
| Condition_E2 × NDS b | 0.227 | 0.143 | −0.055, 0.509 | 1.587 | 0.113 |
| Condition_E3 × NDS b | 0.051 | 0.151 | −0.248, 0.350 | 0.337 | 0.736 |
| Condition_NE × NDS b | 0.096 | 0.160 | −0.219, 0.411 | 0.604 | 0.546 |
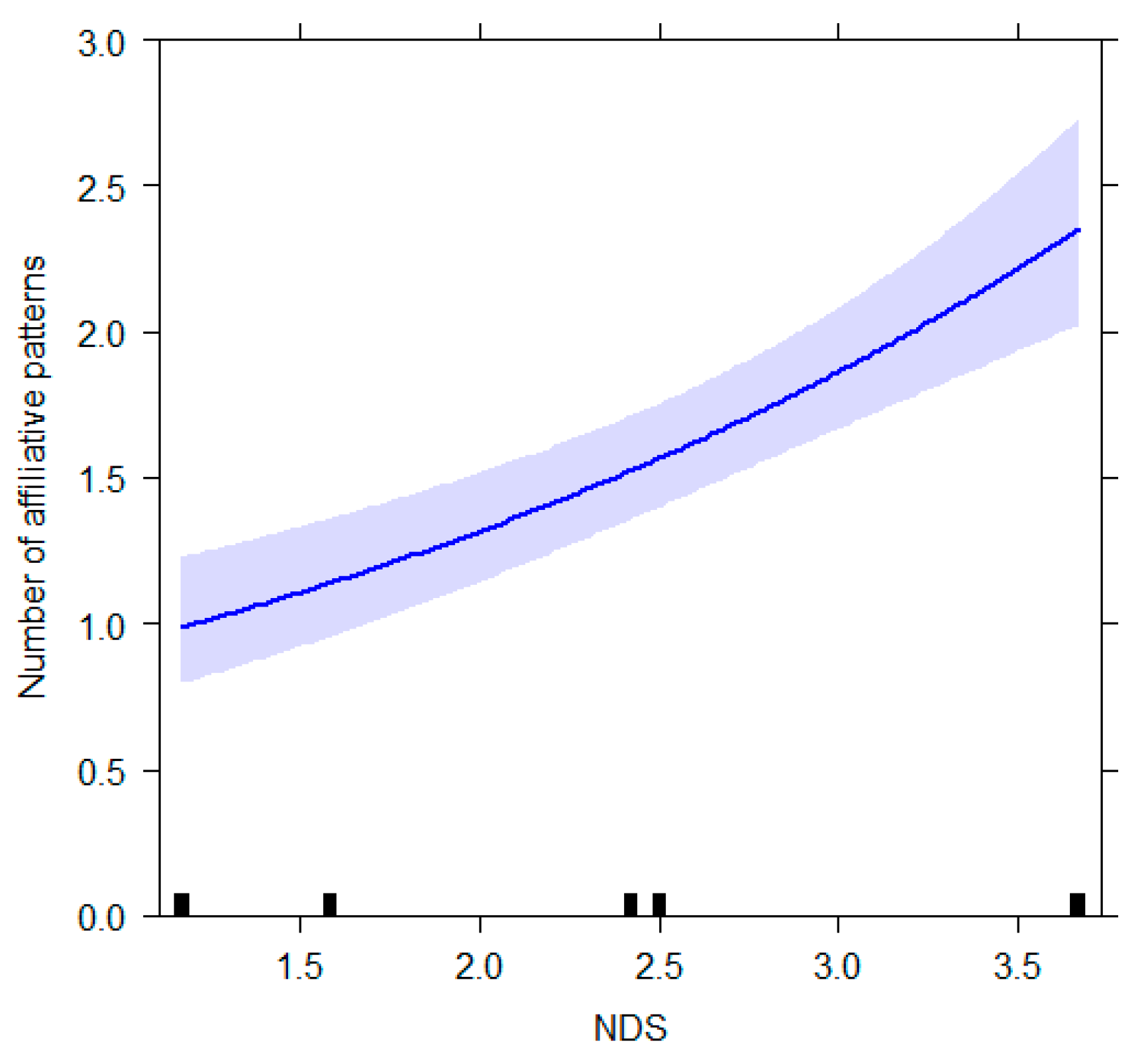
| GLMM4 (affiliative behavior) | |||||
| Intercept a | −1.679 | 0.882 | −3.418, 0.061 | a | a |
| Age | 0.002 | 0.021 | −0.039, 0.042 | 0.089 | 0.929 |
| NDS | 0.358 | 0.116 | 0.130, 0.587 | 3.094 | 0.002 |
| Condition_E1 b | −0.605 | 0.569 | −1.727, 0.517 | −1.063 | 0.288 |
| Condition_E2 b | −1.204 | 0.659 | −2.503, 0.095 | −1.827 | 0.068 |
| Condition_E3 b | −0.493 | 0.587 | −1.651, 0.665 | −0.839 | 0.401 |
| Condition_NE b | 0.162 | 0.615 | −1.051, 1.376 | 0.264 | 0.792 |
| Day of week_weekend b | −0.075 | 0.220 | −0.508, 0.358 | −0.343 | 0.732 |
| Ambient temperature | 0.053 | 0.027 | −0.001, 0.107 | 1.920 | 0.055 |
| Condition_E1 × age b | 0.007 | 0.029 | −0.050, 0.064 | 0.237 | 0.813 |
| Condition_E2 × age b | 0.049 | 0.037 | −0.025, 0.123 | 1.311 | 0.190 |
| Condition_E3 × age b | 0.023 | 0.032 | −0.041, 0.086 | 0.706 | 0.480 |
| Condition_NE × age b | −0.032 | 0.033 | −0.098, 0.033 | −0.978 | 0.328 |
| Condition_E1 × NDS b | 0.115 | 0.169 | −0.218, 0.449 | 0.680 | 0.496 |
| Condition_E2 × NDS b | −0.076 | 0.209 | −0.488, 0.337 | −0.362 | 0.718 |
| Condition_E3 × NDS b | −0.075 | 0.179 | −0.428, 0.278 | −0.417 | 0.677 |
| Condition_NE × NDS b | −0.084 | 0.178 | −0.435, 0.266 | −0.474 | 0.636 |
| GLMM5 (agonistic behavior) | |||||
| Intercept a | 3.712 | 2.550 | −1.327, 8.751 | a | a |
| Age | −0.126 | 0.041 | −0.207, −0.044 | −3.052 | 0.002 |
| NDS | −0.260 | 0.148 | −0.552, 0.032 | −1.758 | 0.079 |
| Condition_E1 b | 2.317 | 0.720 | 0.898, 3.736 | 3.220 | 0.001 |
| Condition_E2 b | 1.759 | 0.697 | 0.384, 3.134 | 2.522 | 0.012 |
| Condition_E3 b | 1.666 | 0.757 | 0.174, 3.159 | 2.201 | 0.028 |
| Condition_NE b | 1.824 | 0.727 | 0.391, 3.257 | 2.509 | 0.012 |
| Day of week_weekend b | −0.421 | 0.630 | −1.664, 0.822 | −0.668 | 0.504 |
| Ambient temperature | −0.158 | 0.090 | −0.336, 0.020 | −1.750 | 0.080 |
| GLMM6 (explorative and scent-marking behavior) | |||||
| Intercept a | 1.879 | 0.932 | 0.040, 3.717 | a | a |
| Age | −0.019 | 0.026 | −0.071, 0.032 | −0.739 | 0.460 |
| NDS | 0.316 | 0.140 | 0.040, 0.592 | 2.257 | 0.024 |
| Condition_E1 b | 2.003 | 0.645 | 0.732, 3.275 | 3.108 | 0.002 |
| Condition_E2 b | 1.339 | 0.655 | 0.048, 2.631 | 2.045 | 0.041 |
| Condition_E3 b | 1.467 | 0.662 | 0.162, 2.772 | 2.217 | 0.027 |
| Condition_NE b | 1.545 | 0.719 | 0.127, 2.964 | 2.149 | 0.032 |
| Day of week_weekend b | 0.102 | 0.216 | −0.325, 0.529 | 0.471 | 0.637 |
| Ambient temperature | −0.075 | 0.029 | −0.132, −0.018 | −2.607 | 0.009 |
| Condition_E1 × age b | −0.052 | 0.037 | −0.125, 0.022 | −1.389 | 0.165 |
| Condition_E2 × age b | −0.033 | 0.037 | −0.107, 0.041 | −0.882 | 0.378 |
| Condition_E3 × age b | −0.032 | 0.038 | −0.107, 0.043 | −0.839 | 0.402 |
| Condition_NE × age b | −0.051 | 0.043 | −0.135, 0.033 | −1.196 | 0.231 |
| Condition_E1 × NDS b | −0.323 | 0.179 | −0.677, 0.031 | −1.802 | 0.072 |
| Condition_E2 × NDS b | −0.226 | 0.185 | −0.592, 0.139 | −1.223 | 0.221 |
| Condition_E3 × NDS b | −0.274 | 0.186 | −0.642, 0.093 | −1.471 | 0.141 |
| Condition_NE × NDS b | −0.357 | 0.199 | −0.750, 0.035 | −1.796 | 0.073 |
| GLMM7 (foraging) | |||||
| Intercept a | 1.871 | 0.832 | 0.230, 3.512 | a | a |
| Age | −0.012 | 0.025 | −0.062, 0.037 | −0.493 | 0.622 |
| NDS | −0.006 | 0.117 | −0.237, 0.224 | −0.054 | 0.957 |
| Condition_E1 b | 0.120 | 0.539 | −0.864, 1.264 | 0.371 | 0.711 |
| Condition_E2 b | −0.430 | 0.536 | −1.488, 0.627 | −0.802 | 0.422 |
| Condition_E3 b | −0.796 | 0.636 | −2.050, 0.457 | −1.253 | 0.210 |
| Condition_NE b | 0.299 | 0.056 | −0.806, 1.404 | 0.534 | 0.593 |
| Day of week_weekend b | −0.559 | 0.250 | −1.053, −0.066 | −2.235 | 0.025 |
| Ambient temperature | −0.035 | 0.026 | −0.087, 0.016 | −1.355 | 0.176 |
| Condition_E1 × age b | 0.016 | 0.035 | −0.054, 0.086 | 0.442 | 0.658 |
| Condition_E2 × age b | 0.056 | 0.036 | −0.016, 0.127 | 1.536 | 0.125 |
| Condition_E3 × age b | 0.008 | 0.039 | −0.070, 0.085 | 0.196 | 0.845 |
| Condition_NE × age b | 0.027 | 0.038 | −0.048, 0.103 | 0.714 | 0.475 |
| Condition_E1 × NDS b | −0.075 | 0.163 | −0.396, 0.246 | −0.461 | 0.645 |
| Condition_E2 × NDS b | −0.055 | 0.169 | −0.388, 0.279 | −0.323 | 0.747 |
| Condition_E3 × NDS b | 0.155 | 0.190 | −0.219, 0.529 | 0.816 | 0.414 |
| Condition_NE × NDS b | −0.206 | 0.173 | −0.546, 0.135 | −1.192 | 0.233 |
| Predictors | Estimates | SEM | ꭓ2 | p |
| E1 vs. BL | 2.317 | 0.720 | 3.220 | 0.010 |
| E2 vs. BL | 1.759 | 0.697 | 2.522 | 0.080 |
| E3 vs. BL | 1.666 | 0.757 | 2.201 | 0.170 |
| NE vs. BL | 1.824 | 0.727 | 2.509 | 0.083 |
| E2 vs. E1 | −0.558 | 0.385 | −1.451 | 0.581 |
| E3 vs. E1 | −0.651 | 0.414 | −1.572 | 0.501 |
| NE vs. E1 | −0.493 | 0.404 | −1.222 | 0.727 |
| E3 vs. E2 | −0.093 | 0.455 | −0.204 | 1.000 |
| NE vs. E2 | 0.065 | 0.434 | 0.149 | 1.000 |
| NE vs. E3 | 0.158 | 0.455 | 0.346 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caselli, M.; Messeri, P.; Dessì-Fulgheri, F.; Bandoli, F. Enriching Zoo-Housed Ring-Tailed Lemurs (Lemur catta): Assessing the Influence of Three Types of Environmental Enrichment on Behavior. Animals 2022, 12, 2836. https://doi.org/10.3390/ani12202836
Caselli M, Messeri P, Dessì-Fulgheri F, Bandoli F. Enriching Zoo-Housed Ring-Tailed Lemurs (Lemur catta): Assessing the Influence of Three Types of Environmental Enrichment on Behavior. Animals. 2022; 12(20):2836. https://doi.org/10.3390/ani12202836
Chicago/Turabian StyleCaselli, Marta, Patrizia Messeri, Francesco Dessì-Fulgheri, and Francesca Bandoli. 2022. "Enriching Zoo-Housed Ring-Tailed Lemurs (Lemur catta): Assessing the Influence of Three Types of Environmental Enrichment on Behavior" Animals 12, no. 20: 2836. https://doi.org/10.3390/ani12202836