

Garlic and Its Bioactive Compounds: Implications for Methane Emissions and Ruminant Nutrition


Abstract
:Simple Summary
Abstract
1. Greenhouse Gas Emissions from Ruminants
1.1. Greenhouse Gas Emissions from Ruminants and the Contribution of Methane
1.2. Global Targets for the Mitigation of CH4 Emissions
1.3. The Role of Ruminants’ Diet in Mitigation of CH4 Emissions
2. An Introduction to Rumen CH4 Synthesis
2.1. The Rumen Microbiome and Metabolic Pathways of CH4 Synthesis in the Rumen
2.2. Targeted Manipulation of Ruminant Metabolic Pathways to Reduce CH4 Synthesis
3. Garlic and Ruminant CH4 Emissions
3.1. The Need to Exploit Plant-Derived Bioactive Compounds
3.2. Effect of Garlic on CH4 Emissions: In Vitro Assessments
3.3. Effect of Garlic on CH4 Emissions: In Vivo Assessments

{kind=link}
{kind=link}
{kind=link}
| Form | Garlic Bioactive Compound (Mode of Action) | Antibacterial | Antiprotozoal | Antifungal | Reference |
|---|---|---|---|---|---|
| DAS | |||||
| DAS (purity, 97%) | Diallyl sulphide (binding to thiol-containing proteins/enzymes in bacterial cells) | Cronobacter sakazakii | ND | ND | [115] |
| Garlic extracts | |||||
| Garlic extracts | ND | ND | Taenia taeniaeformis, Hymenolepis microstoma, H. diminuta, Echinostoma caproni, and Fasciola hepatica | ND | [116] |
| Garlic extracts | Thiosulphinates and Allicin (thiol enzyme inhibition and preventing the parasite’s RNA, DNA, and protein synthesis) | ND | Blastocystis spp. | ND | [117] |
| Garlic extracts | DATS (affecting the fungal cell wall and causing irreversible ultrastructural changes in the fungal cells, leading to loss of structural integrity) | ND | ND | Trichophyton verrucosum, T.mentagrophytes, T. rubrum, Botrytis cinerea, Candida species, Epidermophyton floccosum, Aspergillus niger, A. flavus, Rhizopus stolonifera, Microsporum gypseum, M. audouinii, Alternaria alternate, Neofabraea alba, and Penicillium expansum | [118] |
| Garlic extracts | Allicin (oxidative interaction with important thiol-containing enzymes) | Bacillus, Escherichia, Mycobacterium, Pseudomonas, Staphylococcus, and Streptococcus | ND | Aspergillus niger, Penicillium cyclopium, and Fusarium oxysporum | [119] |
| Garlic extracts | Allicin (reacts with cysteine-containing Burkholderia enzymes involved in key biosynthetic pathways) | B. cenocepacia C6433 | ND | ND | [120] |
| Garlic extracts | Allicin (interferes with RNA production and lipid synthesis) | Bacillus subtilis, Staphylococcus aureus, Escherichia coli, and Klebsiella pneumonia | ND | Candida albicans | [121] |
| Garlic extracts | Allicin (interferes with RNA production and lipid synthesis) | S. aureus | ND | ND | [122] |
| Garlic extracts | Spasmolytic effect was most likely mediated through Ca2+-channel inhibition | Salmonella enteritidis, Escherichia coli, Proteus mirabilis, and Enterococcus faecalis | ND | ND | [123] |
| Garlic extracts | Allicin (reduced serum total oxidative status, malondialdehyde, and nitric oxide production, and increased total thiols) | ND | ND | Meyerozyma guilliermondii and Rhodotorula mucilaginosa | [124] |
| Garlic extracts | ND | Bacillus, Enterobacter, Enterococcus, Escherichia, Klebsiella, Listeria, Pseudomonas, Salmonella, and Staphy lococcus | ND | Candida albicans | [125] |
| Garlic oil | |||||
| Garlic oil | DAS (the presence of the allyl group is fundamental for the antimicrobial activity of these sulphide derivatives when they are present in Allium) | Staphylococcus aureus, Pseudomonas aeruginosa, and Escherichia coli | ND | ND | [126] |
| Garlic oil | Ajoene (inhibiting the human glutathione reductase and T. cruzi trypanothione reductase) | ND | Cochlospermum planchonii, Plasmodium, Giardia, Leishmania, and Trypanosoma. | ND | [127] |
| Garlic oil | DAS (the richness in sulphur atoms may have contributed to the effectiveness of the EO activity) | Staphylococus aureus, Salmonella Typhimurium, Listeria monocytogenes, Escherichia coli, Campylobacter jejuni | ND | ND | [128] |
| Garlic oil | Allicin (inactivation of allicin by cysteine groups of mucin or other gastrointestinal bacteria) | Campylobacter jejuni | ND | ND | [129] |
| Type of Study | Garlic Form Supplementation | Level of Supply | Basal Diet | CH4 Yield | Reference |
|---|---|---|---|---|---|
| In Vitro | |||||
| Batch culture | |||||
| Batch culture (sheep rumen fluid) | Garlic and citrus extracts | 0%, 10%, and 20% of DMI | Concentrate and grass at 50:50 ratio | ↓ 11% (from 11.12 mL/g DM to 9.89 mL/g DM) | [130] |
| Batch culture (sheep rumen fluid) | Bulb of garlic | 70 mg | 450 mg DM substrate (a mixture of lucerne hay (500 g/kg), grass hay (200 g/kg), and barley (300 g/kg)) | ↓ 9.8% (from 1.32 mmol/g DM to 1.19 mmol/g DM) | [98] |
| Batch culture (sheep rumen fluid) | ALL and DAD | 0.5, 5, and 10 mg/L | 1:1 alfalfa hay:concentrate either (HF inoculum; 700:300 alfalfa hay:concentrate; 4 sheep) or (HC inoculum, 300:700 alfalfa hay:concentrate; 4 sheep) | ND | [39] |
| Batch culture (sheep rumen fluid) | Garlic oil | 0, 20, 60, 180, or 540 mg/L | 300 mg MC (500:500 alfalfa hay:concentrate), and the other 4 were fed HC (150:850 barley straw:concentrate) | ↓ 12.1% (from 0.262 mmol/L of VFA to 0.257 mmol/L of VFA) | [99] |
| Batch culture (cow rumen fluid) | Garlic extracts | 1% of total volume | 0.3 g of timothy | ↓ 20% (from 40.2 mL/g DM to 32.5 mL/g DM) | [95] |
| Batch culture (buffalo rumen fluid) | Coconut oil and garlic powder | 0:0, 16:0, 8:4, 4:8, and 0:16 mg | 200 mg DM (60:40 roughage (R) and concentrate (C) ratio were used as substrates) | ND | [29] |
| Batch culture (sheep rumen fluid) | Garlic oil and cinnamaldehyde | 0, 20, 60, 180, and 540 mg/L | Forages and concentrates 50:50 alfalfa hay:concentrate diet (MC) and 15:85 barley straw:concentrate diet (HC) | ND | [104] |
| Batch culture and dual flow continuous culture (cow rumen fluid) | Garlic oil | 3, 30, 300, and 3000 mg/L | 50:50 forage:concentrate diet | ↓ 73.6% (from 0.20 mmol/L of VFA to 0.07 mmol/L of VFA) | [37] |
| Batch culture (cow rumen fluid) | Combination of garlic oil, nitrate, and saponin | Garlic oil (0.25 g/L), nitrate (5 mM), and quillaja saponin (0.6 g/L) | 400 mg of ground feed substrate. The feed substrate is a mixture of alfalfa hay and a dairy concentrate feed at a 50:50 ratio | ↓ 65% at day 2 (from 29.1 mL/g DM to 10.3 mL/g DM) and by 40% at day 18 (from 21.4 mL/g DM to 13 mL/g DM) | [48] |
| Batch culture (cow rumen fluid) | Garlic powder | 2–6% of DMI | Concentrate and wheat straw at a 50:50 ratio | ND | [113] |
| CCF | |||||
| CCF (goat rumen fluid) | PTS | 200 μL/L/day | Alfalfa hay and concentrate in a 50:50 ratio | ↓ 48% (from 249 mmol/L of VFA to 129 mmol/L of VFA) | [131] |
| Rusitec | |||||
| Rusitec (cow rumen fluid) | Mootral (garlic and citrus extract) | 1–2 g | 7 g hay and 3 g concentrate | ↓ 96% (from 10.70 mL/g DM to 0.40 mL/g DM) | [96] |
| Rusitec (cow rumen fluid) | Garlic oil | 300 mg/L | A basal diet (15 g DM/d) consisting of ryegrass hay, barley, and soya bean meal (1:0.7:0.3) | ↓ 91% (from 7.96 mL/g DM to 0.73 mL/g DM) | [102] |
| In Vivo | |||||
| Buffalo | |||||
| Buffalo | Coconut oil and garlic powder | 7% coconut oil plus 100 g/d of garlic powder | Rice straw ad libitum, concentrate 0.5% BW | ↓ 9% (from 27.5 mmol/L of VFA to 25 mmol/L of VFA) | [111] |
| Buffalo | Garlic powder | 2% of DMI | Concentrate and roughage diet which comprised of concentrate mixture, berseem, and wheat straw | ↓ 33% (from 40.70 g/kg DMI to 27 g/kg DMI) | [113] |
| Buffalo | A mixture of garlic and soapnut in 2:1 ratio | 2% of DMI | Wheat straw and concentrate mixture at a ratio of 60:40 | ↓ 12.6% (from 36.30 g/kg DMI to 31.72 g/kg DMI) | [132] |
| Buffalo | Mixture of garlic bulb and peppermint oil | 2.5% of DMI | 50% wheat straw and 50% concentrate | ↓ 7.4% (from 29.17 g/kg DMI to 27.01 g/kg DMI) | [133] |
| Cattle | |||||
| Cattle | Mootral (garlic and citrus extract) | 15 g/d | TMR at a ratio of 47% forage and 53% concentrate | ↓ 23.2% (from 19.4 g/kg DMI to 14.9 g/kg DMI) | [106] |
| Cattle Cattle Cattle | Garlic powder A mixture of mangosteen peel, garlic, and urea pellet | 40 g/d 200 g/d 200 g/d | Concentrate at 5 g/kg BW with UTRS fed ad libitum Rice straw ad libitum and concentrate were fed at 0.5% of BW Concentrate at 0.5% of BW while rice straw was fed ad libitum | ↓ 5% (from 29.3 mmol/L of VFA to 27.9 mmol/L of VFA) ↓ 6.5% (from 27.6 mmol/L of VFA to 25.8 mmol/L of VFA) | [87] [134] [134] [135] |
| Goat | |||||
| Goat | Garlic oil | 20–35 g | 600 g/kg DM of concentrate and 400 g/kg DM of cowpea/maize silage in a ratio of 1:3 | ND | [109] |
| Sheep | |||||
| Sheep | ALL | 2 g/head day | TMR | ↓ 7.7% (from 66.1 g/kg DMI to 61 g/kg DMI) | [107] |
| Sheep | FDGL | 2.5 g/(kg BW0.75·d) | Mixed hay plus concentrate at 60:40 ratio | ↓ 10% (from 28.05 g/kg DMI to 25.34 g/kg DMI) | [114] |
| Sheep | Garlic powder | 0.5% concentrate (DM) | Concentrate to rice straw at ratio of 30:70 | ↓ 6.6% (from 42.3 g/kg DMI to 39.5 g/kg DMI) | [136] |
| Sheep | Combined garlic essential oil and linseed oil | Linseed oil (1.6 mL/kg BW) and garlic essential oil (3 μL/kg BW) | Free access to a natural grassland hay 921.1 g DM/kg and concentrate 889.0 g DM/kg | ↓ 19.6% (from 19.68 g/kg DMI to 15.81 g/kg DMI) | [112] |
4. Bioactive Compounds in Garlic That Decrease CH4 Emissions and the Potential Effect on Biochemical Pathways
| Animal | Basal Diet | Garlic form Supplementation | Bioactive Compound | Level of Supply | Effects | Reference |
|---|---|---|---|---|---|---|
| Buffalo | ||||||
| Buffalo | Concentrate was offered at 0.5% of BW, while rice straw was given on ad libitum basis | Coconut oil and garlic powder | ND | 7% coconut oil plus 100 g/d of garlic powder | ↑ BUN22; C3; Total bacteria population; Amylolytic and proteolytic bacteria; rumen ecology ↓ CH4; Total VFA; C2; C2/C3 ratio; protozoal population | [111] |
| Buffalo | Concentrate and roughage diet which comprised of concentrate mixture, berseem, and wheat straw | Garlic powder | ND | 2% of DMI | ↑ Milk production; Digestibility↓ CH4 | [113] |
| Buffalo Buffalo | Wheat straw and concentrate mixture at a ratio of 60:40 50% wheat straw and 50% concentrate mixture | A mixture of (garlic and soapnut in 2:1 ratio A mixture of garlic bulb and peppermint oil | ND ND | 2% of DMI 2.5% of DMI | ↑ urinary nitrogen; feed conversion efficiency ↓ CH4; faecal nitrogen ↓ CH4 | [132] [133] |
| Cattle | ||||||
| Cattle | TMR according to the National Academies of Sciences, Engineering, and Medicine | Mootral (garlic and citrus extract) | ALL and flavonoid | 15 g/d | ↓ CH4
| [106] |
| DMI, average daily gain, and feed efficiency remained similar in control and supplemented steers | ||||||
| Cattle Cattle | Concentrate at 5 g/kg BW UTRS fed ad libitum Rice straw ad libitum and concentrate were fed at 0.5% of BW | Garlic powder | ALL, ajoene, S-allylcysteine, DAD, S-methylcysteine sulphoxide, and S-allylcysteine A mixture of mangosteen peel, garlic, and urea pellet | 40 g/d 200 g/d | ↑ pH; C3; rumen fermentation efficiency ↓ CP digestibility; NH3-N; C2; CH4; Population sizes of bacteria and protozoa; proteolytic bacteria; amylolytic and cellulolytic bacteria ↑ NH3-N; C3; bacterial population; rumen fermentation, microbial protein synthesis ↓ CH4; protozoa population | [87] [134] |
| Cow | ||||||
| Cow | TMR | Garlic essential oil | ALL | 5 g/kg DM | ↑ Feed digestibility ↓ The flow of bypass protein to the small intestine | [149] |
| Cow | TMR | DAD | DAD | DAD was fed at levels of 56 mg/kg DM and 200 mg/kg DM in Exp. 1 and Exp. 2, respectively. This is equivalent to 1.0 or 3.3 g/cow per day | [150] | |
| Cow | Fed with ad libitum with UTRS and concentrate at 0.5 g kg−1 body weight (BW) twice daily | Garlic powder | ND | 80 g d−1 | ↑ C3; N retention and absorption ↓ C2/C3; Protozoa | [151] |
| Goat | ||||||
| Goat | 600 g/kg DM of concentrate and 400 g/kg DM of cowpea/maize silage in a ratio of 1:3, respectively | Garlic oil | ND | 20–35 g | ↑ ADF & lignin digestibility, total VFA, FCR, NH3-N, digestibility ↓ CH4; Protozoa | [109] |
| Goat | Grass hay (Leymus chinensis, 0.38 kg/d dry matter (DM)) and concentrate (0.22 kg/d DM) | Garlic oil | ND | 0.8 g/d | [152] | |
| Sheep | ||||||
| Ewe | TMR | ALL | ALL | 2 g/d | OM; N; NDF; ADF digestibility ↓ CH4; protozoa and methanogens | [107] |
| Ewe | TMR based on barley-based diet | Garlic oil | ALM (26%), allyl trisulphide (18%), ALL (1.5%) | 0.02 g/kg DM | ↑ Methanosphaera stadtmanae, Methanobrevibacter smithii Alter the diversity of rumen methanogens without affecting the methanogenic capacity of the rumen | [108] |
| Lamb | A barley-based concentrate diet ad libitum | Garlic essential oil | ND | 200 mg/kg DM |
| [103] |
| Lamb | Free access to a natural grassland hay [921.1 g dry matter (DM)/kg and concentrate (889.0 g DM/kg)] | Combined garlic essential oil and linseed oil | ND | Linseed oil (1.6 mL/kg BW) and garlic essential oil (3 μL/kg BW) | ↓ CH4; VFA
| [112] |
| Lamb | According to Ministry of Agriculture of P. R. China, 2004 | Garlic skin | ND | 80 g/kg DM | ↑ ADG; VFA; Prevotella, Bulleidia, Howardella, Methanosphaera ↓ Fretibacterium
| [91] |
| Sheep | Control diet (basal total mixed ration with no additive = CTR) | Raw garlic or garlic oil | ND | Dose of raw garlic (75 versus 100 g/kg DM) and garlic oil (500 versus 750 mg/kg DM) | C3; C2/C3 ratio ↓ NDF; ADF by garlic oil supplementation; Protozoa in a dose-independent manner; NH3 | [105] |
| Sheep | Mixed hay (Hay-diet, as control) and hay plus garlic stem and leaf silage diet (GS-diet, at ratio of 9:1) | Garlic stem and leaf silage | ND | 66 g/kg BW 0.75/d DM | ↑ Nitrogen digestibility; C3; C5; Glucose; plasma LeuTR and WBPS NEFA | [101] |
| Sheep | Meadow hay (3rd cut, vented) and concentrate (barley grain and soybean meal; 700:300) offered in a 1:1 ratio | Garlic oil | DAD | 5 g garlic oil or 2 g DAD/kg DM | ↑ digestibility and energy use efficiency ↓ concentrate intake; Low palatability | [110] |
| Sheep | Mixed hay plus concentrate at 60:40 ratio | FDGL | ALL | 2.5 g/(kg BW 0.75·d) | ↑ NH3-N; Glucose ↓ CH4; DM ingested | [114] |
| Sheep | Forage to concentrate ratio of 1:1 | Bulb of garlic | ND | 1% of DM | ↑ Nutrient digestibility (DM, OM, NDF, ADF, and cellulose) | [93] |
| Ram | Concentrate to rice straw was 30:70 (as-fed basis) | Garlic powder | ND | 0.5% concentrate (DM) | ↓ CH4; Serum glutamic oxaloacetic transaminase | [136] |
5. Nutritive Value of Garlic in Ruminants
5.1. Chemical Composition of Garlic
5.2. Effects of Garlic on Rumen Fermentation
5.3. Effects of Garlic on Rumen Microbiota
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization: Rome, Italy, 2013. [Google Scholar]
- Steinfeld, H.; Wassenaar, T.; Jutzi, S. Livestock production systems in developing countries: Status, drivers, trends. Rev. Sci. Tech. 2006, 25, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organisation (FAO). World Livestock 2011: Livestock in Food Security World, Food and Agriculture Organization of the United Nations; Food and Agriculture Organization: Rome, Italy, 2011. [Google Scholar]
- Dragosits, U.; Chadwick, D.R.; Del Prado, A.; Scholefield, D.; Mills, J.A.; Crompton, L.A.; Newbold, C.J. Implications of farm-scale methane mitigation measures for national methane emissions. In Land Management in a Changing Environment; Agriculture and the Environment, VII; Crighton, K., Audsley, R., Eds.; SAC and SEPA: Stirling, UK, 2008; pp. 168–174. [Google Scholar]
- Thorpe, A. Enteric fermentation and ruminant eructation: The role (and control?) of methane in the climate change debate. Clim. Change 2009, 93, 407–431. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, F.-M.D.; Breon, W.; Collins, J.; Fuglestvedt, J.; Huang, D.; Koch, J.-F.; Lamarque, D.; Lee, B.; Mendoza, T.; et al. Anthropogenic and Natural Radiative Forcing. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 659–740. [Google Scholar]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; ISBN 978-1-107-05799-1. [Google Scholar]
- Pachauri, R.K.A.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P. Climate Change: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2014. [Google Scholar]
- Clauss, M.; Dittmann, M.T.; Vendl, C.; Hagen, K.B.; Frei, S.; Ortmann, S.; Muller, D.W.H.; Hammer, S.; Munn, A.J.; Schwarm, A.; et al. Review: Comparative methane production in mammalian herbivores. Animal 2020, 14, s113–s123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Council Directorate-General for Climate Action. Going Climate-Neutral by 2050; Publications Office of the European Union: Luxembourg, 2019. [Google Scholar]
- Tubiello, F.N.; Salvatore, M.; Rossi, S.; Ferrara, A.; Fitton, N.; Smith, P. The FAOSTAT database of greenhouse gas emissions from agriculture. Environ. Res. Lett. 2013, 8, 015009. [Google Scholar] [CrossRef]
- Huppmann, D.; Rogelj, J.; Kriegler, E.; Krey, V.; Riahi, K. A new scenario resource for integrated 1.5 °C research. Nat. Clim. Change 2018, 8, 1027–1030. [Google Scholar] [CrossRef]
- Rogelj, J.; Shindell, D.; Jiang, K.; Fifita, S.; Forster, P.; Ginzburg, V.; Handa, C.; Kheshgi, H.; Kobayashi, S.; Kriegler, E. Global Warming of 1.5 °C; An IPCC Special Report on the impacts of global warming of 1.5 °C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2018; Available online: https://www.ipcc.ch/site/assets/uploads/sites/2/2019/05/SR15_Chapter2_Low_Res.pdf (accessed on 20 October 2022).
- Mbow, C.; Rosenzweig, C.; Barioni, L.; Benton, T.; Herrero, M.; Krishnapillai, M.; Liwenga, E.; Pradhan, P.; Rivera-Ferre, M.; Sapkota, T. Food Security in Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019; Available online: https://www.ipcc.ch/site/assets/uploads/sites/4/2019/11/08_Chapter-5.pdf (accessed on 20 October 2022).
- Hristov, A.N.; Oh, J.; Lee, C.; Meinen, R.; Montes, F.; Ott, T.; Firkins, J.; Rotz, A.; Dell, C.; Adesogan, A.; et al. Mitigation of Greenhouse Gases Emissions in Livestock Production: A Review of Technical Options for Non-CO2 Emissions; FAO Animal Production and Health Paper No. 177; Gerber, P.J., Henderson, B., Makkar, H.P.S., Eds.; Food and Agriculture Organization: Rome, Italy, 2013; pp. 1–226. [Google Scholar]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty years of research on rumen methanogenesis: Lessons learned and future challenges for mitigation. Animal 2020, 14, s2–s16. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.; Frezal, C.; Flynn, E. A Survey of GHG Mitigation Policies for the Agriculture, Forestry and Other Land Use Sector; OECD Publishing: Paris, France, 2020. [Google Scholar] [CrossRef]
- Reisinger, A.; Clark, H.; Cowie, A.L.; Emmet-Booth, J.; Gonzalez Fischer, C.; Herrero, M.; Howden, M.; Leahy, S. How necessary and feasible are reductions of methane emissions from livestock to support stringent temperature goals? Philos. Trans. R. Soc. A 2021, 379, 20200452. [Google Scholar] [CrossRef]
- Golub, A.A.; Henderson, B.B.; Hertel, T.W.; Gerber, P.J.; Rose, S.K.; Sohngen, B. Global climate policy impacts on livestock, land use, livelihoods, and food security. Proc. Natl. Acad. Sci. USA 2013, 110, 20894–20899. [Google Scholar] [CrossRef] [Green Version]
- Eisler, M.C.; Lee, M.R.; Tarlton, J.F.; Martin, G.B.; Beddington, J.; Dungait, J.A.; Greathead, H.; Liu, J.; Mathew, S.; Miller, H.; et al. Agriculture: Steps to sustainable livestock. Nature 2014, 507, 32–34. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Global Warming of 1.5 °C; An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global GHGemission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2018. [Google Scholar]
- Valero, M.; Do Prado, R.; Zawadzki, F.; Eiras, C.; Madrona, G. Propolis and essential oils additives in the diets improved animal performance and feed efficiency of bulls finished in feedlot. Acta Sci. Anim. Sci. 2014, 36, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Broucek, J. Production of Methane Emissions from Ruminant Husbandry: A Review. J. Environ. Prot. 2014, 5, 1482–1493. [Google Scholar] [CrossRef]
- Haque, M.N. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 2018, 60, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckard, R.J.; Clark, H. Potential solutions to the major greenhouse-gas issues facing Australasian dairy farming. Anim. Prod. Sci. 2018, 60, 10–16. [Google Scholar] [CrossRef]
- Honan, M.; Feng, X.; Tricarico, J.; Kebreab, E. Feed additives as a strategic approach to reduce enteric methane production in cattle: Modes of action, effectiveness and safety. Anim. Prod. Sci. 2022, 62, 1303–1317. [Google Scholar] [CrossRef]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Beauchemin, K.; McAllister, T.; McGinn, S. Dietary mitigation of enteric methane from cattle. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2009, 4, 035. [Google Scholar] [CrossRef]
- Kongmun, P.; Wanapat, M.; Pakdee, P.; Navanukraw, C. Effect of coconut oil and garlic powder on in vitro fermentation using gas production technique. Livest. Sci. 2010, 127, 38–44. [Google Scholar] [CrossRef]
- Bayat, A.R.; Kairenius, P.; Stefański, T.; Leskinen, H.; Comtet-Marre, S.; Forano, E.; Chaucheyras-Durand, F.; Shingfield, K.J. Effect of camelina oil or live yeasts (Saccharomyces cerevisiae) on ruminal methane production, rumen fermentation, and milk fatty acid composition in lactating cows fed grass silage diets. J. Dairy Sci. 2015, 98, 3166–3181. [Google Scholar] [CrossRef] [Green Version]
- Ku-Vera, J.C.; Jimenez-Ocampo, R.; Valencia-Salazar, S.S.; Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Gomez-Bravo, C.A.; Aguilar-Perez, C.F.; Solorio-Sanchez, F.J. Role of Secondary Plant Metabolites on Enteric Methane Mitigation in Ruminants. Front. Vet. Sci 2020, 7, 584. [Google Scholar] [CrossRef]
- Patra, A.K.; Yu, Z. Effects of essential oils on methane production and fermentation by, and abundance and diversity of, rumen microbial populations. Appl. Environ. Microbiol. 2012, 78, 4271–4280. [Google Scholar] [CrossRef] [Green Version]
- Krause, D.O.; Denman, S.E.; Mackie, R.I.; Morrison, M.; Rae, A.L.; Attwood, G.T.; McSweeney, C.S. Opportunities to improve fiber degradation in the rumen: Microbiology, ecology, and genomics. FEMS Microbiol. Rev. 2003, 27, 663–693. [Google Scholar] [CrossRef] [Green Version]
- McSweeney, C.S.; Denman, S.E.; Wright, A.D.G.; Yu, Z. Application of Recent DNA/RNA-based Techniques in Rumen Ecology. Asian-Australas. J. Anim. Sci. 2007, 20, 283–294. [Google Scholar] [CrossRef]
- Maron, D.F.; Smith, T.J.S.; Nachman, K.E. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Glob. Health 2013, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Saxena, J. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 2010, 71, 1198–1222. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Carro, M.D.; Kamel, C. Effect of garlic oil and four of its compounds on rumen microbial fermentation. J. Dairy Sci. 2005, 88, 4393–4404. [Google Scholar] [CrossRef] [Green Version]
- Calsamiglia, S.; Busquet, M.; Cardozo, P.W.; Castillejos, L.; Ferret, A. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 2007, 90, 2580–2595. [Google Scholar] [CrossRef] [Green Version]
- Kamel, C.; Greathead, H.M.R.; Tejido, M.L.; Ranilla, M.J.; Carro, M.D. Effects of allicin and diallyl disulfide on in vitro rumen fermentation of a mixed diet. Anim. Feed Sci. Technol. 2008, 145, 351–363. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Church, D.C. The Ruminant Animal: Digestive Physiology and Nutrition; Waveland Press: Prospect Heights, IL, USA, 1993. [Google Scholar]
- Morgavi, D.P.; Kelly, W.J.; Janssen, P.H.; Attwood, G.T. Rumen microbial (meta)genomics and its application to ruminant production. Animal 2013, 7 (Suppl. 1), 184–201. [Google Scholar] [CrossRef] [Green Version]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, E.; Hristov, A.N. Nitrogen and Phosphorus Nutrition of Cattle; CABI: Oxfordshire, UK, 2005. [Google Scholar]
- Sirohi, S.; Singh, N.; Dagar, S.; Puniya, A. Molecular tools for deciphering the microbial community structure and diversity in rumen ecosystem. Appl. Microbiol. Biotechnol. 2012, 95, 1135–1154. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, M.; Ramos-Morales, E.; Wallace, R.J. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, A.K.; Yu, Z. Effects of Adaptation of In Vitro Rumen Culture to Garlic Oil, Nitrate, and Saponin and Their Combinations on Methanogenesis, Fermentation, and Abundances and Diversity of Microbial Populations. Front. Microbiol. 2015, 6, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newbold, C.J.; Lassalas, B.; Jouany, J.P. The importance of methanogens associated with ciliate protozoa in ruminal methane production in vitro. Lett. Appl. Microbiol. 1995, 21, 230–234. [Google Scholar] [CrossRef]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The ruminal microbiome associated with methane emissions from ruminant livestock. J. Anim. Sci. Biotechnol. 2017, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Pinares-Patiño, C.S.; Baumont, R.; Martin, C. Methane emissions by Charolais cows grazing a monospecific pasture of timothy at four stages of maturity. Can. J. Anim. Sci. 2003, 83, 769–777. [Google Scholar] [CrossRef]
- Mizrahi, I. Rumen Symbioses. In The Prokaryotes: Prokaryotic Biology and Symbiotic Associations; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 533–544. [Google Scholar]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef] [Green Version]
- Moss, A.R.; Jouany, J.-P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef] [Green Version]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial ecosystem and methanogenesis in ruminants. Animal 2010, 4, 1024–1036. [Google Scholar] [CrossRef]
- Hopkins, A.; Del Prado, A. Implications of climate change for grassland in Europe: Impacts, adaptations and mitigation options: A review. Grass Forage Sci. 2007, 62, 118–126. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- McAllister, T.A.; Newbold, C.J. Redirecting rumen fermentation to reduce methanogenesis. Aust. J. Exp. Agric. 2008, 48, 7–13. [Google Scholar] [CrossRef]
- Demeyer, D.I. Methanogenesis, and integrated part of carbohydrate fermentation and its control. In Digestion and Metabolism in the Ruminant; McDonald, L.W., Warner, A.C.I., Eds.; The University of New England Publishing Unit: Armidale, Australia, 1975; pp. 366–382. [Google Scholar]
- Saengkerdsub, S.; Ricke, S.C. Ecology and characteristics of methanogenic archaea in animals and humans. Crit. Rev. Microbiol. 2014, 40, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, M.; Schwab, C.; Jensen, B.B.; Engberg, R.M.; Spang, A.; Canibe, N.; Højberg, O.; Milinovich, G.; Fragner, L.; Schleper, C.; et al. Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen. Nat. Commun. 2013, 4, 1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boadi, D.; Benchaar, C.; Chiquette, J.; Massé, D. Mitigation strategies to reduce enteric methane emissions from dairy cows: Update review. Can. J. Anim. Sci. 2004, 84, 319–335. [Google Scholar] [CrossRef] [Green Version]
- Ferry, J.G. Fundamentals of methanogenic pathways that are key to the biomethanation of complex biomass. Curr. Opin. Biotechnol. 2011, 22, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Dridi, B.; Fardeau, M.L.; Ollivier, B.; Raoult, D.; Drancourt, M. Methanomassiliicoccus luminyensis gen. nov., sp. nov., a methanogenic archaeon isolated from human faeces. Int J. Syst. Evol. Microbiol. 2012, 62, 1902–1907. [Google Scholar] [CrossRef] [Green Version]
- Borrel, G.; Harris, H.M.; Tottey, W.; Mihajlovski, A.; Parisot, N.; Peyretaillade, E.; Peyret, P.; Gribaldo, S.; O’Toole, P.W.; Brugère, J.F. Genome sequence of “Candidatus Methanomethylophilus alvus” Mx1201, a methanogenic archaeon from the human gut belonging to a seventh order of methanogens. J. Bacteriol. 2012, 194, 6944–6945. [Google Scholar] [CrossRef] [Green Version]
- Weiland, P. Biogas production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 85, 849–860. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Xie, T.Y.; Janssen, P.H.; Sun, X.Z.; Beauchemin, K.A.; Tan, Z.L.; Gao, M. Shifts in Rumen Fermentation and Microbiota Are Associated with Dissolved Ruminal Hydrogen Concentrations in Lactating Dairy Cows Fed Different Types of Carbohydrates. J. Nutr. 2016, 146, 1714–1721. [Google Scholar] [CrossRef] [Green Version]
- Ugbogu, E.A.; Elghandour, M.M.; Ikpeazu, V.O.; Buendía, G.R.; Molina, O.M.; Arunsi, U.O.; Emmanuel, O.; Salem, A.Z. The potential impacts of dietary plant natural products on the sustainable mitigation of methane emission from livestock farming. J. Clean. Prod. 2019, 213, 915–925. [Google Scholar] [CrossRef]
- McGinn, S.M.; Beauchemin, K.A.; Coates, T.; Colombatto, D. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and fumaric acid. J. Anim. Sci. 2004, 82, 3346–3356. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Nan, X.; Chu, K.; Tong, J.; Yang, L.; Zheng, S.; Zhao, G.; Jiang, L.; Xiong, B. Shifts of Hydrogen Metabolism from Methanogenesis to Propionate Production in Response to Replacement of Forage Fiber with Non-forage Fiber Sources in Diets in vitro. Front. Microbiol. 2018, 9, 2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newbold, C.J.; McIntosh, F.M.; Williams, P.; Losa, R.; Wallace, R.J. Effects of a specific blend of essential oil compounds on rumen fermentation. Anim. Feed Sci. Technol. 2004, 114, 105–112. [Google Scholar] [CrossRef]
- Benchaar, C.; Calsamiglia, S.; Chaves, A.; Fraser, G.; Colombatto, D.; McAllister, T.; Beauchemin, K. A review of plant-derived essential oils in ruminant nutrition and production. Anim. Feed Sci. Technol. 2008, 145, 209–228. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Tamminga, S.; Bannink, A.; Dijkstra, J.; Zom, R.L.G. Feeding Strategies to Reduce Methane Loss in Cattle; Animal Sciences Group: Lelystad, The Netherlands, 2007. [Google Scholar]
- Greathead, H. Plants and plant extracts for improving animal productivity. Proc. Nutr. Soc. 2003, 62, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouany, J.P.; Morgavi, D.P. Use of ‘natural’ products as alternatives to antibiotic feed additives in ruminant production. Animal 2007, 1, 1443–1466. [Google Scholar] [CrossRef] [Green Version]
- Canul-Solis, J.; Campos-Navarrete, M.; Pineiro-Vazquez, A.; Casanova-Lugo, F.; Barros-Rodriguez, M.; Chay-Canul, A.; Cardenas-Medina, J.; Castillo-Sanchez, L. Mitigation of Rumen Methane Emissions with Foliage and Pods of Tropical Trees. Animals 2020, 10, 843. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [Green Version]
- Food and Drug Administration (FDA). Food and Drug Administration of the US; CFR 184; Food and Drug Administration: Montgomery, MD, USA, 2004; Volume 21. [Google Scholar]
- Cieslak, A.; Szumacher-Strabel, M.; Stochmal, A.; Oleszek, W. Plant components with specific activities against rumen methanogens. Animal 2013, 7 (Suppl. 2), 253–265. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K. An Overview of Antimicrobial Properties of Different Classes of Phytochemicals. In Dietary Phytochemicals and Microbes; Springer: Dordrecht, The Netherlands, 2012; pp. 1–32. [Google Scholar] [CrossRef]
- Ahmed, E.; Fukuma, N.; Hanada, M.; Nishida, T. The Efficacy of Plant-Based Bioactives Supplementation to Different Proportion of Concentrate Diets on Methane Production and Rumen Fermentation Characteristics In Vitro. Animals 2021, 11, 1029. [Google Scholar] [CrossRef] [PubMed]
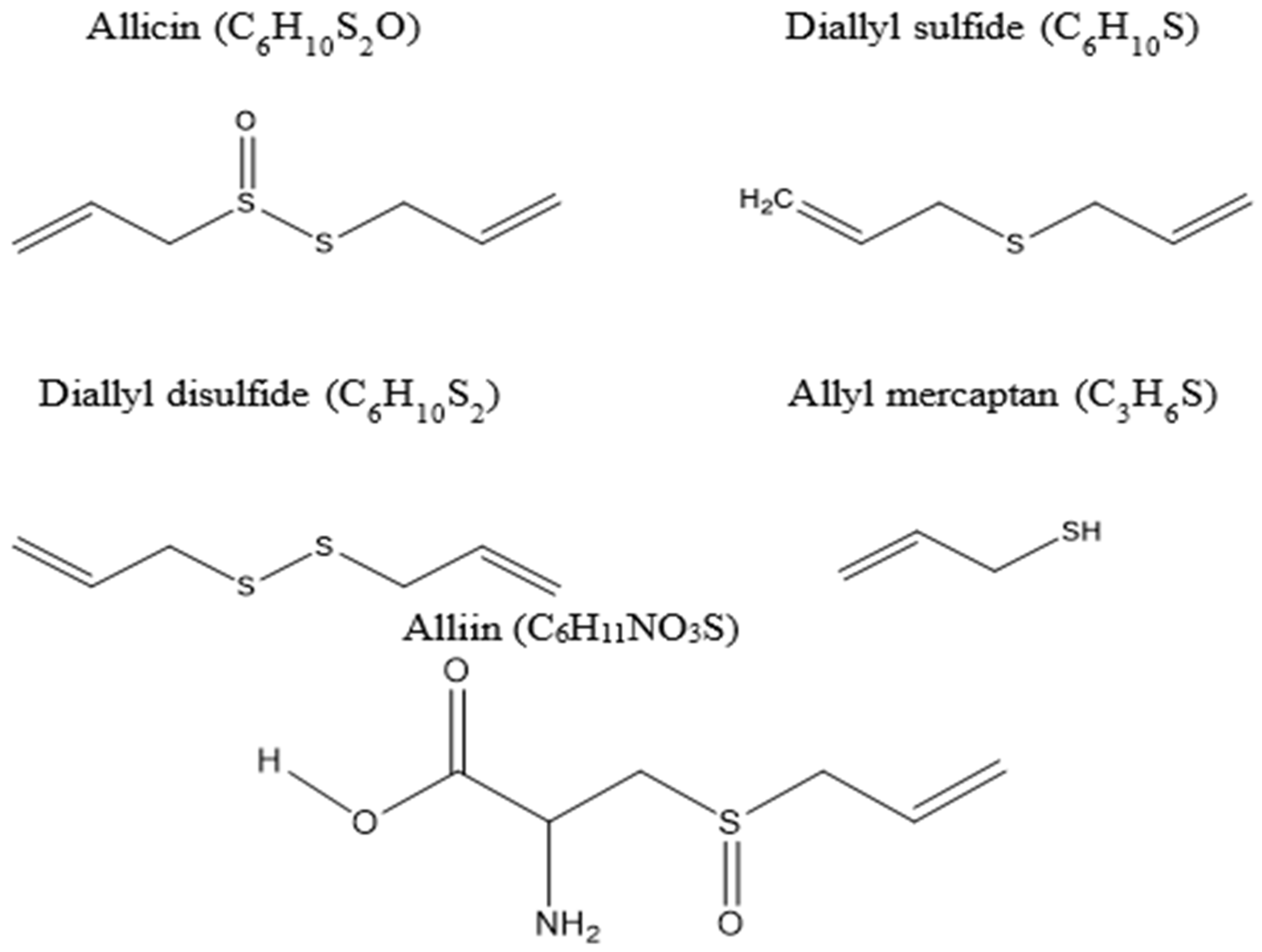
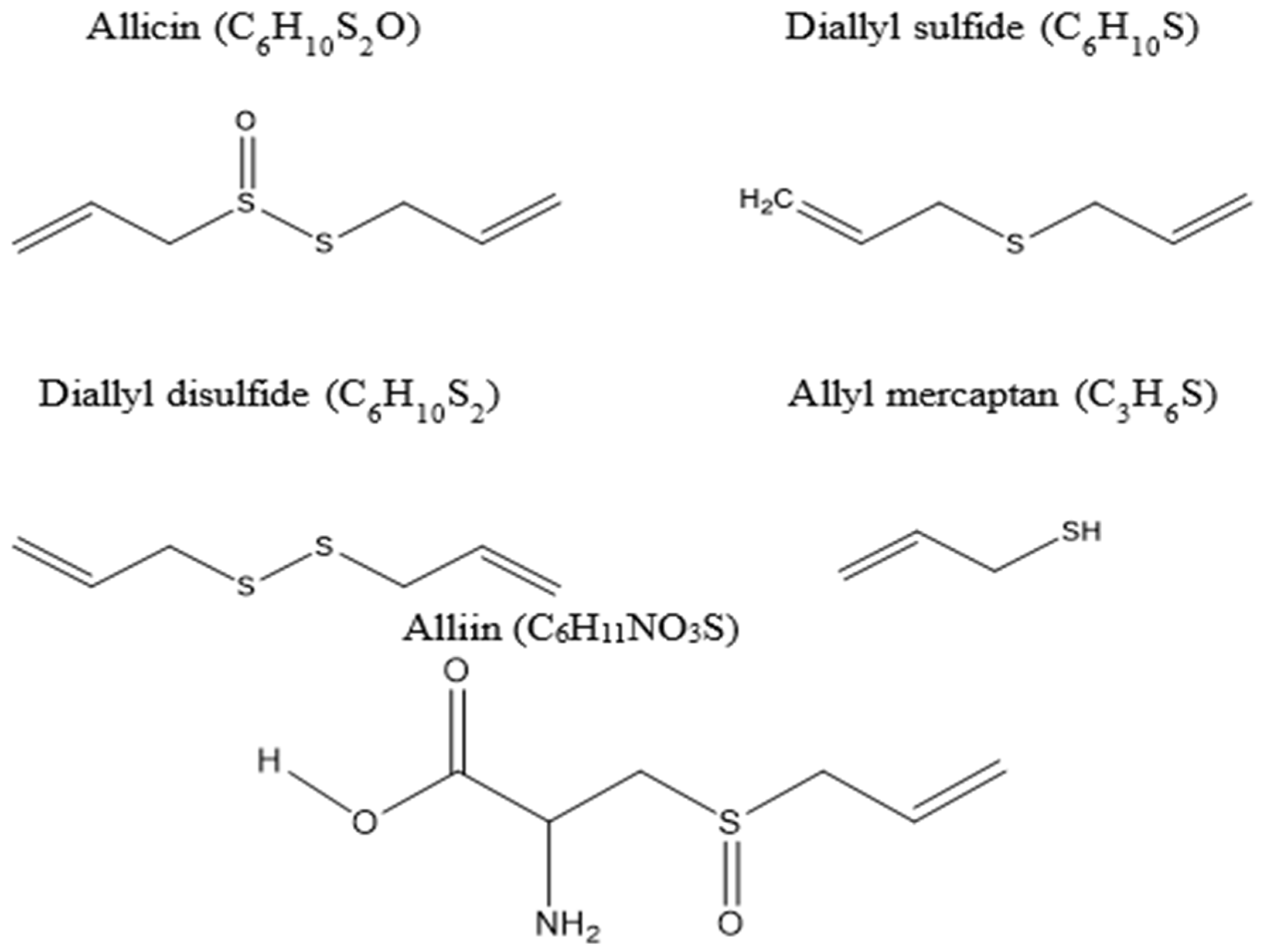
- Ankri, S.; Mirelman, D. Antimicrobial properties of allicin from garlic. Microbes Infect. 1999, 1, 125–129. [Google Scholar] [CrossRef]
- Filocamo, A.; Nueno-Palop, C.; Bisignano, C.; Mandalari, G.; Narbad, A. Effect of garlic powder on the growth of commensal bacteria from the gastrointestinal tract. Phytomedicine 2012, 19, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Ruddock, P.S.; Liao, M.; Foster, B.C.; Lawson, L.; Arnason, J.T.; Dillon, J.A. Garlic natural health products exhibit variable constituent levels and antimicrobial activity against Neisseria gonorrhoeae, Staphylococcus aureus and Enterococcus faecalis. Phytother Res. 2005, 19, 327–334. [Google Scholar] [CrossRef]
- Peinado, M.J.; Ruiz, R.; Echávarri, A.; Rubio, L.A. Garlic derivative propyl propane thiosulfonate is effective against broiler enteropathogens in vivo. Poult. Sci. 2012, 91, 2148–2157. [Google Scholar] [CrossRef]
- Wanapat, M.; Kang, S.; Khejornsart, P.; Wanapat, S. Effects of plant herb combination supplementation on rumen fermentation and nutrient digestibility in beef cattle. Asian-Australas. J. Anim. Sci. 2013, 26, 1127–1136. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Xie, K.; Liu, Z.; Nakasone, Y.; Sakao, K.; Hossain, A.; Hou, D.X. Preventive Effects and Mechanisms of Garlic on Dyslipidemia and Gut Microbiome Dysbiosis. Nutrients 2019, 11, 1225. [Google Scholar] [CrossRef]
- Vogt, L.; Meyer, D.; Pullens, G.; Faas, M.; Smelt, M.; Venema, K.; Ramasamy, U.; Schols, H.A.; De Vos, P. Immunological properties of inulin-type fructans. Crit. Rev. Food Sci. Nutr. 2015, 55, 414–436. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Ma, M.P.; Diao, Q.Y.; Tu, Y. Saponin-Induced Shifts in the Rumen Microbiome and Metabolome of Young Cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Su, Z.; Xu, W.; Sun, H.X.; Gao, J.F.; Tu, D.F.; Ren, C.H.; Zhang, Z.J.; Cao, H.G. Garlic skin induces shifts in the rumen microbiome and metabolome of fattening lambs. Animal 2021, 15, 100216. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei-Aghsaghali, A.; Syadati, S.A.; Fathi, H.; Rasouli, S.; Sadaghian, M.; Tarahomi, M. Garlic in Ruminants Feeding. Asian J. Biol. Sci. 2012, 5, 328–340. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Kamra, D.N.; Bhar, R.; Kumar, R.; Agarwal, N. Effect of Terminalia chebula and Allium sativum on in vivo methane emission by sheep. J. Anim. Physiol. Anim. Nutr. 2011, 95, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Caribu Hadi, P.; Hidayat, N. The Efficacy of Methanol Extract of Garlic (Allium sativum) to Improve Rumen Fermentation Products. Anim. Prod. 2013, 15, 69–75. [Google Scholar]
- Kim, E.T.; Kim, C.H.; Min, K.S.; Lee, S.S. Effects of Plant Extracts on Microbial Population, Methane Emission and Ruminal Fermentation Characteristics in In vitro. Asian-Australas. J. Anim. Sci. 2012, 25, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Eger, M.; Graz, M.; Riede, S.; Breves, G. Application of Mootral(TM) Reduces Methane Production by Altering the Archaea Community in the Rumen Simulation Technique. Front. Microbiol. 2018, 9, 2094. [Google Scholar] [CrossRef]
- Patra, A.; Kamra, D.N.; Agarwal, N. Effect of plants containing secondary metabolites on in vitro methanogenesis, enzyme profile and fermentation of feed with rumen liquor of buffalo. Anim. Nutr. Feed Technol. 2006, 6, 203–213. [Google Scholar]
- García-González, R.; López, S.; Fernández, M.; Bodas, R.; González, J.S. Screening the activity of plants and spices for decreasing ruminal methane production in vitro. Anim. Feed Sci. Technol. 2008, 147, 36–52. [Google Scholar] [CrossRef]
- Kamel, C.; Greathead, H.M.R.; Ranilla, M.J.; Tejido, M.L.; Ramos, S.; Carro, M.D. Effects of Garlic Oil on In Vitro Rumen Fermentation and Methane Production Are Influenced by the Basal Diet. Available online: https://en.engormix.com/dairy-cattle/articles/effects-garlic-oil-vitro-t34553.htm (accessed on 29 August 2022).
- Cardozo, P.W.; Calsamiglia, S.; Ferret, A.; Kamel, C. Screening for the effects of natural plant extracts at different pH on in vitro rumen microbial fermentation of a high-concentrate diet for beef cattle. J. Anim. Sci. 2005, 83, 2572–2579. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; Torita, A.; Sako, Y.; Al-Mamun, M.; Sano, H. Effects of feeding garlic stem and leaf silage on rates of plasma leucine turnover, whole body protein synthesis and degradation in sheep. Small Ruminant Res. 2011, 99, 37–43. [Google Scholar] [CrossRef]
- Soliva, C.R.; Amelchanka, S.L.; Duval, S.M.; Kreuzer, M. Ruminal methane inhibition potential of various pure compounds in comparison with garlic oil as determined with a rumen simulation technique (Rusitec). Br. J. Nutr. 2011, 106, 114–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, A.V.; He, M.L.; Yang, W.Z.; Hristov, A.N.; McAllister, T.A.; Benchaar, C. Effects of essential oils on proteolytic, deaminative and methanogenic activities of mixed ruminal bacteria. Can. J. Anim. Sci. 2008, 88, 117–122. [Google Scholar] [CrossRef]
- Mateos, I.; Ranilla, M.; Tejido, M.; Saro, C.; Kamel, C.; Carro, M. The influence of diet type (dairy versus intensive fattening) on the effectiveness of garlic oil and cinnamaldehyde to manipulate in vitro ruminal fermentation and methane production. Anim. Prod. Sci. 2013, 53, 299. [Google Scholar] [CrossRef] [Green Version]
- Anassori, E.; Dalir-Naghadeh, B.; Pirmohammadi, R.; Taghizadeh, A.; Siamak, A.-R.; Maham, M.; Farahmand-Azar, S.; Farhoomand, P. Garlic: A potential alternative for monensin as a rumen modifier. Livest. Sci. 2011, 142, 276–287. [Google Scholar] [CrossRef]
- Roque, B.M.; Van Lingen, H.J.; Vrancken, H.; Kebreab, E. Effect of Mootral-a garlic- and citrus-extract-based feed additive-on enteric methane emissions in feedlot cattle. Transl. Anim. Sci. 2019, 3, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Chen, D.; Tu, Y.; Zhang, N.; Si, B.; Deng, K.; Diao, Q. Effect of supplementation of allicin on methanogenesis and ruminal microbial flora in Dorper crossbred ewes. J. Anim. Sci. Biotechnol. 2016, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Ohene-Adjei, S.; Chaves, A.V.; McAllister, T.A.; Benchaar, C.; Teather, R.M.; Forster, R.J. Evidence of increased diversity of methanogenic archaea with plant extract supplementation. Microb. Ecol. 2008, 56, 234–242. [Google Scholar] [CrossRef]
- Okoruwa, M.; Edoror, O. Effect of Garlic Oil Supplementation on Intake, Digestibility, Performance and Rumen Function of Goats Fed Silage Based-diet. Eur. J. Nutr. Food Saf. 2019, 10, 104–115. [Google Scholar] [CrossRef]
- Klevenhusen, F.; Zeitz, J.; Duval, S.; Kreuzer, M.; Soliva, C. Garlic oil and its principal component diallyl disulfide fail to mitigate methane, but improve digestibility in sheep. Anim. Feed Sci. Technol. 2011, 166, 356–363. [Google Scholar] [CrossRef]
- Kongmun, P.; Wanapat, M.; Pakdee, P.; Navanukraw, C.; Yu, Z. Manipulation of rumen fermentation and ecology of swamp buffalo by coconut oil and garlic powder supplementation. Livest. Sci. 2011, 135, 84–92. [Google Scholar] [CrossRef]
- Saro, C.; Hohenester, U.M.; Bernard, M.; Lagrée, M.; Martin, C.; Doreau, M.; Boudra, H.; Popova, M.; Morgavi, D.P. Effectiveness of Interventions to Modulate the Rumen Microbiota Composition and Function in Pre-ruminant and Ruminant Lambs. Front. Microbiol. 2018, 9, 1273. [Google Scholar] [CrossRef] [PubMed]
- Zafarian, R.; Manafi, M. Effect of Garlic Powder on Methane Production, Rumen Fermentation and Milk Production of Buffaloes. Annu. Res. Rev. Biol. 2013, 3, 1013–1019. [Google Scholar]
- Panthee, A.; Matsuno, A.; Al-Mamun, M.; Sano, H. Effect of feeding garlic leaves on rumen fermentation, methane emission, plasma glucose kinetics, and nitrogen utilization in sheep. J. Anim. Sci. Technol. 2017, 59, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Eucker, T.P.; Holly, M.K.; Konkel, M.E.; Lu, X.; Wang, S. Investigating the responses of Cronobacter sakazakii to garlic-drived organosulfur compounds: A systematic study of pathogenic-bacterium injury by use of high-throughput whole-transcriptome sequencing and confocal micro-raman spectroscopy. Appl. Environ. Microbiol. 2014, 80, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Ghaffar, F.; Semmler, M.; Al-Rasheid, K.A.; Strassen, B.; Fischer, K.; Aksu, G.; Klimpel, S.; Mehlhorn, H. The effects of different plant extracts on intestinal cestodes and on trematodes. Parasitol. Res. 2011, 108, 979–984. [Google Scholar] [CrossRef]
- Abdel-Hafeez, E.H.; Ahmad, A.K.; Kamal, A.M.; Abdellatif, M.Z.; Abdelgelil, N.H. In vivo antiprotozoan effects of garlic (Allium sativum) and ginger (Zingiber officinale) extracts on experimentally infected mice with Blastocystis spp. Parasitol. Res. 2015, 114, 3439–3444. [Google Scholar] [CrossRef]
- Fufa, B. Anti-bacterial and Anti-fungal Properties of Garlic Extract (Allium sativum): A Review. Microbiol. Res. J. Int. 2019, 28, 1–5. [Google Scholar] [CrossRef]
- Kamra, D.N.; Agarwal, N.; Sakthivel, P.C.; Chaudhary, L.C. Garlic as a rumen modifier for eco-friendly and economic livestock production. J. Appl. Anim. Res. 2012, 40, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Wallock-Richards, D.; Doherty, C.J.; Doherty, L.; Clarke, D.J.; Place, M.; Govan, J.R.; Campopiano, D.J. Garlic revisited: Antimicrobial activity of allicin-containing garlic extracts against Burkholderia cepacia complex. PLoS ONE 2014, 9, e112726. [Google Scholar] [CrossRef] [Green Version]
- Meriga, B.; Mopuri, R.; MuraliKrishna, T. Insecticidal, antimicrobial and antioxidant activities of bulb extracts of Allium sativum. Asian Pac. J. Trop. Med. 2012, 5, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Mozaffari Nejad, A.S.; Shabani, S.; Bayat, M.; Hosseini, S.E. Antibacterial Effect of Garlic Aqueous Extract on Staphylococcus aureus in Hamburger. Jundishapur J. Microbiol. 2014, 7, e13134. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, D.R.; Veljković, M.; Stojanović, N.M.; Gočmanac-Ignjatović, M.; Mihailov-Krstev, T.; Branković, S.; Sokolović, D.; Marčetić, M.; Radulović, N.; Radenković, M. Influence of different wild-garlic (Allium ursinum) extracts on the gastrointestinal system: Spasmolytic, antimicrobial and antioxidant properties. J. Pharm. Pharmacol. 2017, 69, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Pârvu, M.; Moţ, C.A.; Pârvu, A.E.; Mircea, C.; Stoeber, L.; Roşca-Casian, O.; Ţigu, A.B. Allium sativum Extract Chemical Composition, Antioxidant Activity and Antifungal Effect against Meyerozyma guilliermondii and Rhodotorula mucilaginosa Causing Onychomycosis. Molecules 2019, 24, 3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yetgin, A.; Canlı, K.; Altuner, E.M. Comparison of Antimicrobial Activity of Allium sativum Cloves from China and Taşköprü, Turkey. Adv. Pharmacol. Sci. 2018, 2018, 9302840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casella, S.; Leonardi, M.; Melai, B.; Fratini, F.; Pistelli, L. The role of diallyl sulfides and dipropyl sulfides in the in vitro antimicrobial activity of the essential oil of garlic, Allium sativum L., and leek, Allium porrum L. Phytother Res. 2013, 27, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Hazaa, I.K.; Al-Taai, N.A.; Khalil, N.K.; Zakri, A.M. Efficacy of garlic and onion oils on murin experimental Cryptosporidium parvum infection. Al-Anbar J. Vet. Sci. 2016, 9, 69–74. [Google Scholar]
- Mnayer, D.; Fabiano-Tixier, A.S.; Petitcolas, E.; Hamieh, T.; Nehme, N.; Ferrant, C.; Fernandez, X.; Chemat, F. Chemical composition, antibacterial and antioxidant activities of six essentials oils from the Alliaceae family. Molecules 2014, 19, 20034–20053. [Google Scholar] [CrossRef] [Green Version]
- Robyn, J.; Rasschaert, G.; Hermans, D.; Pasmans, F.; Heyndrickx, M. Is allicin able to reduce Campylobacter jejuni colonization in broilers when added to drinking water? Poult. Sci. 2013, 92, 1408–1418. [Google Scholar] [CrossRef]
- Ahmed, E.; Yano, R.; Fujimori, M.; Kand, D.; Hanada, M.; Nishida, T.; Fukuma, N. Impacts of Mootral on Methane Production, Rumen Fermentation, and Microbial Community in an in vitro Study. Front. Vet. Sci. 2021, 7, 623817. [Google Scholar] [CrossRef]
- Martínez-Fernández, G.; Abecia, L.; Martín-García, A.I.; Ramos-Morales, E.; Denman, S.E.; Newbold, C.J.; Molina-Alcaide, E.; Yáñez-Ruiz, D.R. Response of the rumen archaeal and bacterial populations to anti-methanogenic organosulphur compounds in continuous-culture fermenters. FEMS Microbiol. Ecol. 2015, 91, fiv079. [Google Scholar] [CrossRef] [Green Version]
- Samal, L.; Chaudhary, L.; Agarwal, N.; Kamra, D. Impact of phytogenic feed additives on growth performance, nutrient digestion and methanogenesis in growing buffaloes. Anim. Prod. Sci. 2016, 58, 1056–1063. [Google Scholar] [CrossRef]
- Verma, V.; Chaudhary, L.; Agarwal, N.; Bhar, R.; Kamra, D. Effect of feeding mixture of garlic bulb and peppermint oil on methane emission, rumen fermentation and microbial profile in buffaloes. Anim. Nutr. Feed Technol. 2012, 12, 157–164. [Google Scholar]
- Trinh, T.H.N.; Wanapat, M. Effect of mangosteen peel, garlic and urea pellet supplementation on rumen fermentation and microbial protein synthesis of beef cattle. Agric. J. 2012, 7, 95–100. [Google Scholar]
- Manasri, N.; Wanapat, M.; Navanukraw, C. Improving rumen fermentation and feed digestibility in cattle by mangosteen peel and garlic pellet supplementation. Livest. Sci. 2012, 148, 291–295. [Google Scholar] [CrossRef]
- Kim, J.Y.; Ghassemi Nejad, J.; Park, J.Y.; Lee, B.H.; Hanada, M.; Kim, B.W.; Sung, K.I. In vivo evaluation of garlic (Allium sativum) supplementation to rice straw-based diet on mitigation of CH4 and CO2 emissions and blood profiles using crossbreed rams. J. Sci. Food Agric. 2018, 98, 5197–5204. [Google Scholar] [CrossRef]
- Lawson, L. The composition and chemistry of garlic cloves and processed garlic. In The Science and Therapeutic Application of Allium sativum L. and Related Species; Koch, H.P., Lawson, L.D., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1996; pp. 37–107. [Google Scholar]
- Rivlin, R.S. Is garlic alternative medicine? J. Nutr. 2006, 136, 713s–715s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Jin, S.K.; Yang, H.S. Effect of dietary garlic bulb and husk on the physicochemical properties of chicken meat. Poult. Sci. 2009, 88, 398–405. [Google Scholar] [CrossRef]
- Cruz-Martins, N.; Petropoulos, S.; Ferreira, I. Chemical composition and bioactive compounds of garlic (Allium sativum L.) as affected by pre- and post-harvest conditions: A review. Food Chem. 2016, 211, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Kim, D.B.; Jin, W.; Park, J.; Yoon, W.; Lee, Y.; Kim, S.; Lee, S.; Kim, S.; Lee, O.H.; et al. Comparative studies of bioactive organosulphur compounds and antioxidant activities in garlic (Allium sativum L.), elephant garlic (Allium ampeloprasum L.) and onion (Allium cepa L.). Nat. Prod. Res. 2018, 32, 1193–1197. [Google Scholar] [CrossRef]
- Gebhardt, R.; Beck, H. Differential inhibitory effects of garlic-derived organosulfur compounds on cholesterol biosynthesis in primary rat hepatocyte cultures. Lipids 1996, 31, 1269–1276. [Google Scholar] [CrossRef]
- Reuter, H.D.; Koch, H.P.; Lawson, D. Therapeutic Effects and Applications of Garlic and Its Preparations. In Garlic: The Science and Therapeutic Applications of Allium sativum L. and Related Species, 2nd ed.; Lawson, L.D., Koch, H.P., Eds.; William & Wilkins: Baltimore, MA, USA, 1996. [Google Scholar]
- O’Gara, E.A.; Hill, D.J.; Maslin, D.J. Activities of garlic oil, garlic powder, and their diallyl constituents against Helicobacter pylori. Appl. Environ. Microbiol. 2000, 66, 2269–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, J.E.; Andrés, S.; López-Ferreras, L.; Snelling, T.J.; Yáñez-Ruíz, D.R.; García-Estrada, C.; López, S. Dietary supplemental plant oils reduce methanogenesis from anaerobic microbial fermentation in the rumen. Sci. Rep. 2020, 10, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungerfeld, E.M. Shifts in metabolic hydrogen sinks in the methanogenesis-inhibited ruminal fermentation: A meta-analysis. Front. Microbiol. 2015, 6, 37. [Google Scholar] [CrossRef]
- Denman, S.E.; Martinez Fernandez, G.; Shinkai, T.; Mitsumori, M.; McSweeney, C.S. Metagenomic analysis of the rumen microbial community following inhibition of methane formation by a halogenated methane analog. Front. Microbiol. 2015, 6, 1087. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Morgavi, D.P.; Doreau, M. Methane mitigation in ruminants: From microbe to the farm scale. Animal 2010, 4, 351–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.Z.; Benchaar, C.; Ametaj, B.N.; Chaves, A.V.; He, M.L.; McAllister, T.A. Effects of garlic and juniper berry essential oils on ruminal fermentation and on the site and extent of digestion in lactating cows. J. Dairy Sci. 2007, 90, 5671–5681. [Google Scholar] [CrossRef] [Green Version]
- van Zijderveld, S.M.; Dijkstra, J.; Perdok, H.B.; Newbold, J.R.; Gerrits, W.J. Dietary inclusion of diallyl disulfide, yucca powder, calcium fumarate, an extruded linseed product, or medium-chain fatty acids does not affect methane production in lactating dairy cows. J. Dairy Sci. 2011, 94, 3094–3104. [Google Scholar] [CrossRef] [Green Version]
- Wanapat, M.; Khejornsart, P.; Pakdee, P.; Wanapat, S. Effect of supplementation of garlic powder on rumen ecology and digestibility of nutrients in ruminants. J. Sci. Food Agric. 2008, 88, 2231–2237. [Google Scholar] [CrossRef]
- Zhu, Z.; Mao, S.; Zhu, W. Effects of ruminal infusion of garlic oil on fermentation dynamics, Fatty Acid profile and abundance of bacteria involved in biohydrogenation in rumen of goats. Asian-Australas. J. Anim. Sci. 2012, 25, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Amagase, H. Clarifying the real bioactive constituents of garlic. J. Nutr. 2006, 136, 716s–725s. [Google Scholar] [CrossRef] [Green Version]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Cardozo, P.W.; Kamel, C. Effects of cinnamaldehyde and garlic oil on rumen microbial fermentation in a dual flow continuous culture. J. Dairy Sci. 2005, 88, 2508–2516. [Google Scholar] [CrossRef]
- Ferme, D.; Banjac, M.; Calsamiglia, S.; Busquet, M.; Kamel, C.; Avgustin, G. The effects of plant extracts on microbial community structure in a rumen-simulating continuous-culture system as revealed by molecular profiling. Folia Microbiol. 2004, 49, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Hino, T.; Russell, J.B. Effect of reducing-equivalent disposal and NADH/NAD on deamination of amino acids by intact rumen microorganisms and their cell extracts. Appl. Environ. Microbiol. 1985, 50, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, A.; Itabashi, H. Changes in the population of some functional groups of rumen bacteria including methanogenic bacteria by changing the rumen ciliates in calves. J. Gen. Appl. Microbiol. 1995, 41, 377–387. [Google Scholar] [CrossRef]

| In Vitro Studies | Basal Diet (Forage and Concentrate Ratio) | Garlic Form | Level of Supply | Effects | Reference |
|---|---|---|---|---|---|
| Batch culture | |||||
| Batch culture | 1000 g grass/kg ration + 0 g concentrate/kg ration (100:0), 80:20, 60:40, 40:60, and 20:80 | Mixture of garlic and citrus extracts | 200 g/kg of the feed | ↑ Gas and CO2; NH3-N; Total VFA:C3 and C5 ; pH; C2
| [130] |
| Batch culture | 0.5 g DM of a 10:90 forage:concentrate | Garlic extract | 0, 0.3, 3, 30, and 300 mg/L | ↓ C2/C3 ratio; pH; C3 ↓ Total VFA; NH3-N; C2 | [100] |
| Batch culture | Grass and concentrate mixture (50:50) | Sapindus rarak extract with or without garlic extract | 1.8 g/kg Sapindus rarak extract + 0.25 ppm garlic extract | ↑ C3; ruminal fermentation based on feed digestibility, fermentation products, and rumen bacterial population ↓ Crude digestibility; C2; Protozoa | [94] |
| Batch culture | 450 mg DM substrate (a mixture of lucerne hay (500 g/kg), grass hay (200 g/kg), and barley (300 g/kg)) | Bulb of garlic | 70 mg | DM digestibility ↓ CH4; C2/C3 | [98] |
| Batch culture | 1:1 alfalfa hay:concentrate either (HF inoculum; 700:300 alfalfa hay: concentrate; 4 sheep) or (HC inoculum, 300:700 alfalfa hay:concentrate; 4 sheep) | ALL and DAD | 0.5, 5, and 10 mg/L | ↑ C2/C3 ratio at HC ↓ pH; CH4/VFA | [39] |
| Batch culture | 300 mg (MC; 500:500 alfalfa hay:concentrate), and the other 4 were fed (HC; 150:850 barley straw:concentrate) | Garlic oil | 0, 20, 60, 180, or 540 mg/L | C2/C3 ratio; C5 by garlic oil at 60, 180, and 540 mg/L with diet MC ↓ Total VFA by garlic oil 540 for MC diet; C2 by increasing doses of garlic oil; CH4 | [99] |
| Batch culture | 0.3 g of timothy | Garlic extracts | 1% of total volume | ↑ Total VFA; fibrolytic bacteria; F. succinogens C2/C3 ratio; ciliate-associated methanogen; R. flavefaciens | [95] |
| Batch culture | 200 mg DM (60:40 roughage (R) and concentrate (C) ratio were used as substrates) | Coconut oil and garlic powder | 0:0, 16:0, 8:4, 4:8, and 0:16 mg | ↑ C3; Ruminococcus albus at 8:4 mg; at 8:4 and 0:16 mg could improve ruminal fluid fermentation in terms of VFA profile ↓ Gas production; NH3-N; Total VFA; C2/C3 ratio; CH4; Protozoa | [29] |
| Batch culture | Forages and concentrates 50: 50 alfalfa hay:concentrate diet (MC), and the other four received a 15:85 barley straw:concentrate diet (HC) | Garlic oil and cinnamaldehyde | 0, 20, 60, 180, and 540 mg/L | ↑ VFA ↓ CH4/ VFA ratio the effectiveness of garlic oil and cinnamaldehyde in manipulating ruminal fermentation may depend on the characteristics of the diet fed to the animals, which highlights the importance of testing these additives with different diet types | [104] |
| Batch culture and dual flow continuous culture | 50:50 forage:concentrate diet | Garlic oil | 3, 30, 300, and 3000 mg/L | Batch culture ↑ C3; C5 with supplementation of Garlic oil (30 and 300 mg/L), DAD (30 and 300 mg/L), and ALM (300 mg/L) C2 with supplementation of Garlic oil (30 and 300 mg/L), DAD (30 and 300 mg/L), and ALM (300 mg/L) Dual flow Continuous Culture: ↑ Efficiency of energy use in the rumen ↓ CH4 | [37] |
| Batch culture | 200 mg substrate | Bulb of garlic | 30 mg | ↑ Gas production ↓ CH4 Inhibited methanogenesis without adversely affecting other rumen characteristics | [97] |
| Batch culture | 400 mg of ground feed substrate. The feed substrate is a mixture of alfalfa hay and a dairy concentrate feed at a 50:50 ratio | Combination of garlic oil, nitrate, and saponin | garlic oil (0.25 g/L), nitrate (5 mM), and quillaja saponin (0.6 g/L) | ↑ NH3-N by nitrate at days 10 and 18 ↓ CH4; Feed digestion by the combinations (binary and ternary) of garlic oil with the other inhibitors at days 10 and 18; NH3-N by saponin, alone or in combinations, and garlic oil alone at day 2; Total VFA by garlic oil alone or garlic oil-saponin combination; Methanogens | [48] |
| Batch culture | Concentrate and wheat straw at a 50:50 ratio | Garlic powder | 2–6% of DMI | ↓ CH4; C3; C5 | [113] |
| CCF | |||||
| CCF | Alfalfa hay and concentrate in a 50:50 ratio | PTS | 200 μL/L/day | ↑ Prevotella; Methanobrevibacter and Methanosphaera ↓ CH4; methanogenic archaea; Methanomicrobiales | [131] |
| CCF | 50:50 alfalfa hay:concentrate | Garlic oil | 312 mg/L | C3; C5; Small peptide; NH3-N ↓ C2; VFA | [37] |
| Rusitec | |||||
| Rusitec | 7 g hay and 3 g concentrate | Mootral (garlic and citrus extract) | 1–2 g | SCFA; C5 ↓ CH4; Methanobacteriacea | [96] |
| Rusitec | A basal diet (15 g DM/d) consisting of ryegrass hay, barley, and soya bean meal (1:0·7:0·3) | Garlic oil | 300 mg/L | ↑ Bacterial population ↓ CH4; Protozoa; NDF | [102] |
| In Vivo Studies | Basal Diet (Forage and Concentrate Ratio) | Garlic Form Supplementation | Level of Supply | Effects in Ruminant Productivity | References |
|---|---|---|---|---|---|
| Buffalo | |||||
| Buffalo | Concentrate was offered at 0.5% of BW, while rice straw was given on ad libitum basis | Coconut oil and garlic powder | 7% coconut oil plus 100 g/d of garlic powder | ↑ BUN; C3; Total bacteria population; Amylolytic and proteolytic bacteria; rumen ecology ↓ CH4; Total VFA; C2; C2/C3 ratio; protozoal population | [111] |
| Buffalo | Concentrate and roughage diet which comprised of concentrate mixture, berseem, and wheat straw | Garlic powder | 2% of DMI | ↑ Milk production; Digestibility ↓ CH4 | [113] |
| Cattle | |||||
| Cattle | TMR according to the National Academies of Sciences, Engineering, and Medicine | Mootral (garlic and citrus extract) | 15 g/d | ↓ CH4
| [106] |
| Cattle | Concentrate at 5 g/kg BW with UTRS fed ad libitum | Garlic powder | 40 g/d | ↑ pH; C3; rumen fermentation efficiency ↓ CP digestibility; NH3-N; C2; CH4; Population sizes of bacteria and protozoa; proteolytic bacteria; amylolytic and cellulolytic bacteria | [87] |
| Cow | |||||
| Cow | TMR | DAD | DAD was fed at levels of 56 mg/kg DM and 200 mg/kg DM in Exp. 1 and Exp. 2, respectively. This is equivalent to 1.0 or 3.3 g/cow per day | [150] | |
| Cow | Fed with ad libitum with urea-treated rice straw and concentrate at 0.5 g kg−1 body weight (BW) twice daily | Garlic powder | 80 g d−1 | C3; N retention and absorption C2/C3; Protozoa | [151] |
| Cow | TMR | Garlic essential oil | 5 g/kg DM | ↑ Feed digestibility ↓ The flow of bypass protein to the small intestine | [149] |
| Goat | |||||
| Goat | 600 g/kg DM of concentrate and 400 g/kg DM of cowpea/maize silage in a ratio of 1:3, respectively | Garlic oil | 20–35 g | ↑ ADF & lignin digestibility, total VFA, FCR, NH3-N, digestibility ↓ CH4, protozoa | [109] |
| Goat | Grass hay (Leymus chinensis, 0.38 kg/d DM) and concentrate (0.22 kg/d DM) | Garlic oil | 0.8 g/d | [152] | |
| Sheep | |||||
| Ewe | TMR based on barley-based diet | Garlic oil | 0.02 g/kg DM | ↑ Methanosphaera stadtmanae, Methanobrevibacter smithii Alter the diversity of rumen methanogens without affecting the methanogenic capacity of the rumen | [108] |
| Ewe | TMR | ALL | 2 g/head day | OM; N; NDF; ADF digestibility ↓ CH4; protozoa and methanogens | [107] |
| Lamb | A barley-based concentrate diet ad libitum | Garlic essential oil | 200 mg/kg DM |
| [103] |
| Lamb | Free access to a natural grassland hay [921.1 g dry matter (DM)/kg and concentrate (889.0 g DM/kg)] | Combined garlic essential oil and linseed oil | Linseed oil (1.6 mL/kg BW) and garlic essential oil (3 μL/kg BW) | ↓ CH4; VFA
| [112] |
| Lamb | According to Ministry of Agriculture of P. R. China, 2004 | Garlic skin | 80 g/kg DM | ADG; VFA; Prevotella, Bulleidia, Howardella, Methanosphaera ↓ Fretibacterium
| [91] |
| Sheep | |||||
| Sheep | Control diet (basal total mixed ration with no additive = CTR) | Raw garlic or garlic oil | Dose of raw garlic (75 versus 100 g/kg DM) and garlic oil (500 versus 750 mg/kg DM) | C3; C2/C3 ratio
| [105] |
| Sheep | Mixed hay (Hay-diet, as control) and hay plus garlic stem and leaf silage diet (GS-diet, at ratio of 9:1) | Garlic stem and leaf silage | 66 g/kg BW 0.75/d DM | ↑ Nitrogen digestibility; C3; C5; Glucose; plasma LeuTR and WBPS ↓ Plasma non-esterified fatty acids (NEFA) | [101] |
| Sheep | Meadow hay (3rd cut, vented) and concentrate (barley grain and soybean meal; 700:300) offered in a 1:1 ratio | Garlic oil | 5 g garlic oil or 2 g DAD/kg dietary DM | ↑ Digestibility and energy use efficiency ↓ Concentrate intake; Low palatability | [110] |
| Sheep | Mixed hay plus concentrate at 60:40 ratio | FDGL | 2.5 g/(kg BW 0.75·d) | ↑ NH3-N; Glucose ↓ CH4; DM ingested | [114] |
| Sheep | Forage to concentrate ratio of 1:1 | Bulb of garlic | 1% of DM | ↑ Nutrient digestibility (DM, OM, NDF, ADF, and cellulose) | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sari, N.F.; Ray, P.; Rymer, C.; Kliem, K.E.; Stergiadis, S. Garlic and Its Bioactive Compounds: Implications for Methane Emissions and Ruminant Nutrition. Animals 2022, 12, 2998. https://doi.org/10.3390/ani12212998
Sari NF, Ray P, Rymer C, Kliem KE, Stergiadis S. Garlic and Its Bioactive Compounds: Implications for Methane Emissions and Ruminant Nutrition. Animals. 2022; 12(21):2998. https://doi.org/10.3390/ani12212998
Chicago/Turabian StyleSari, Nurul Fitri, Partha Ray, Caroline Rymer, Kirsty E. Kliem, and Sokratis Stergiadis. 2022. "Garlic and Its Bioactive Compounds: Implications for Methane Emissions and Ruminant Nutrition" Animals 12, no. 21: 2998. https://doi.org/10.3390/ani12212998