
Estradiol Regulates the Expression and Secretion of Antimicrobial Peptide S100A7 via the ERK1/2-Signaling Pathway in Goat Mammary Epithelial Cells
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Primary Antibodies and Chemicals
2.2. Milk Collection and Mammary Epithelial Cells Isolation and Culture
2.3. Immunofluorescence
2.4. RNA Reverse Transcription and Polymerase Chain Reaction
2.5. Western Blot
2.6. ELISA
2.7. Statistics
3. Result
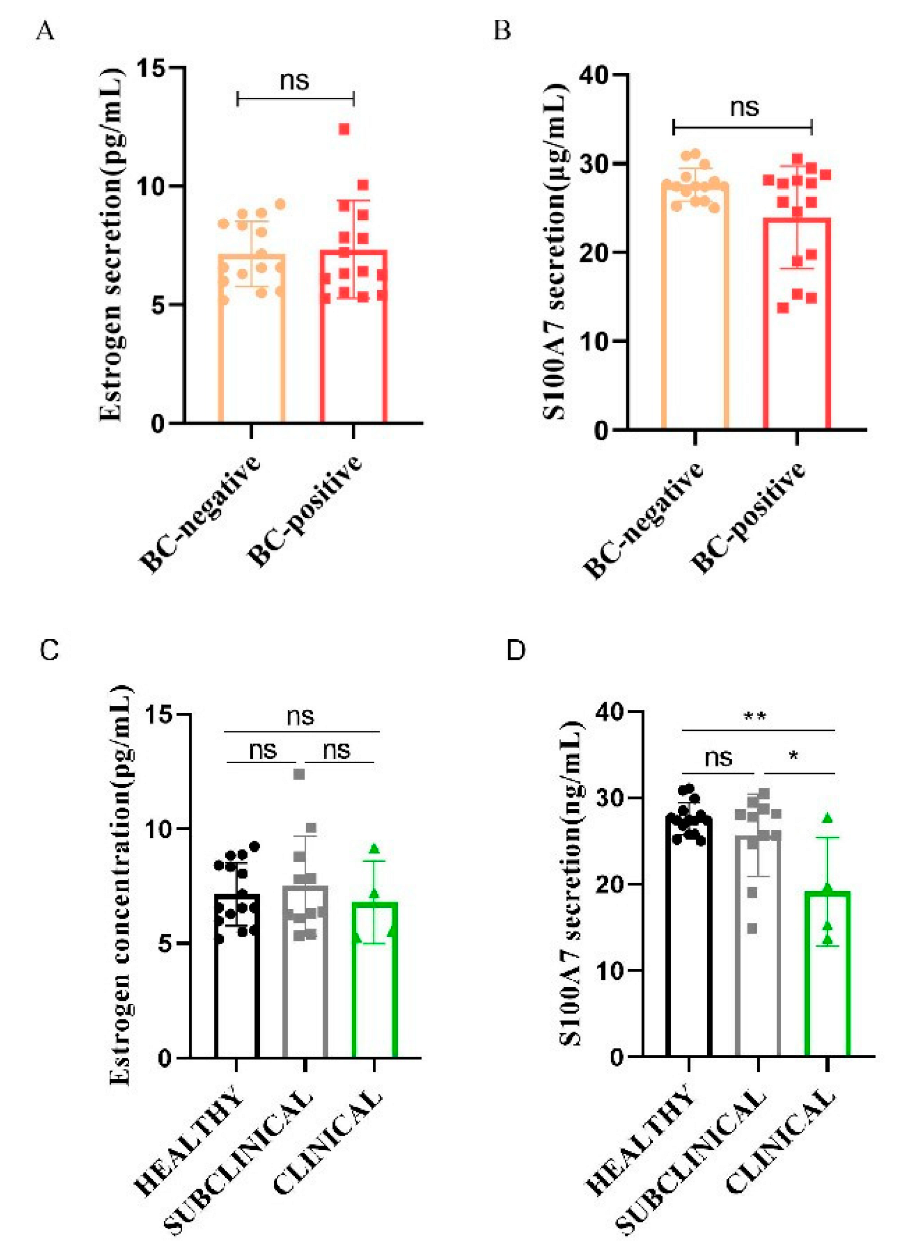
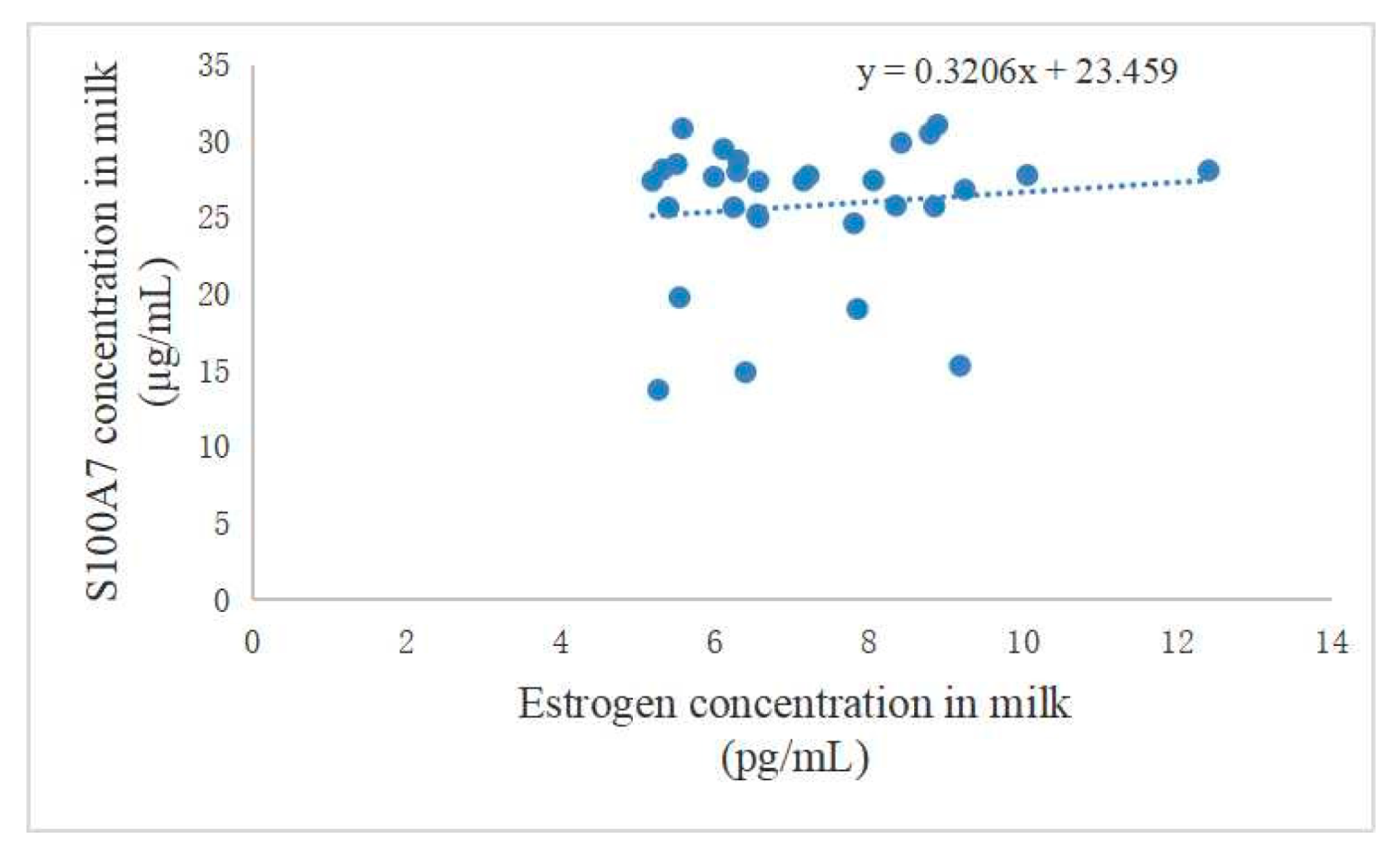
3.1. Difference between Estrogen and S100A7 Abundance in Milk and Their Correlation
3.2. Isolation and Identification of gMECs
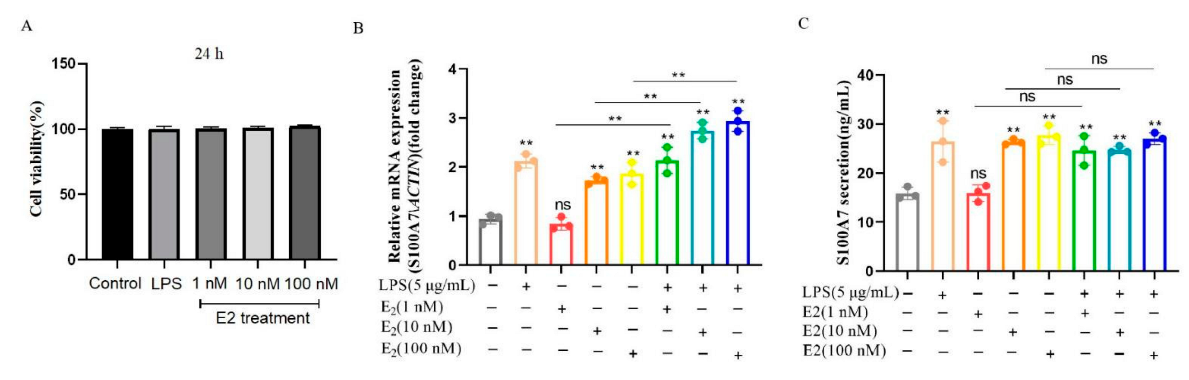
3.3. E2 Induced the mRNA Expression and Secretion of S100A7 in Cultured gMECs
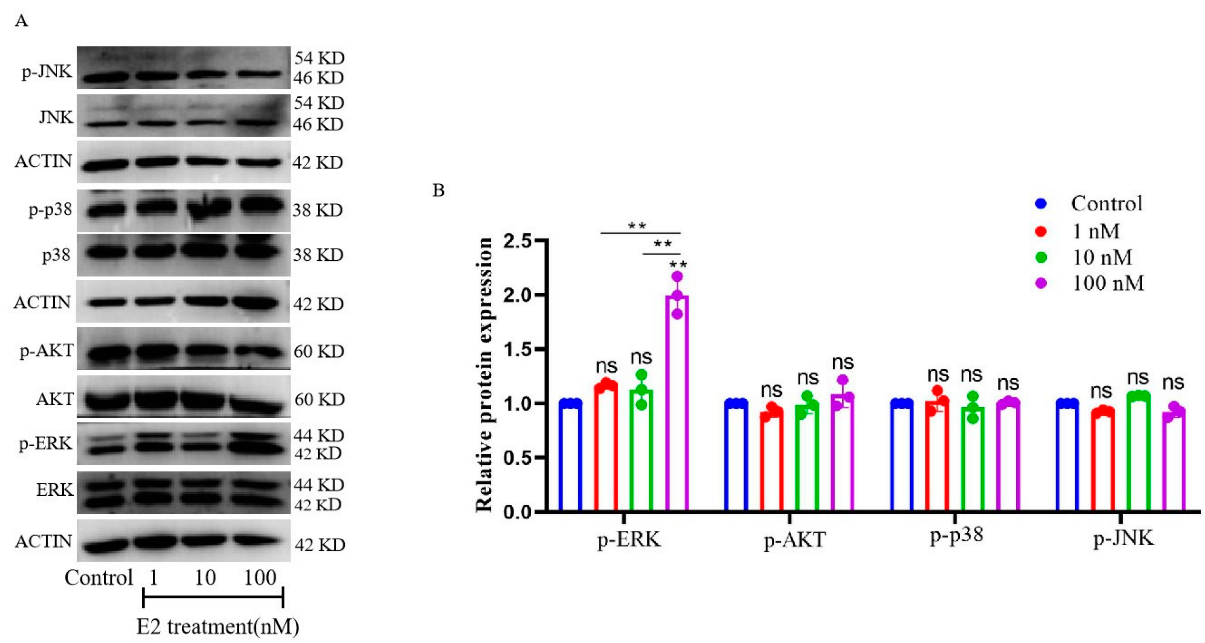
3.4. E2-Activated ERK1/2-Signaling Pathways in Cultured gMECs
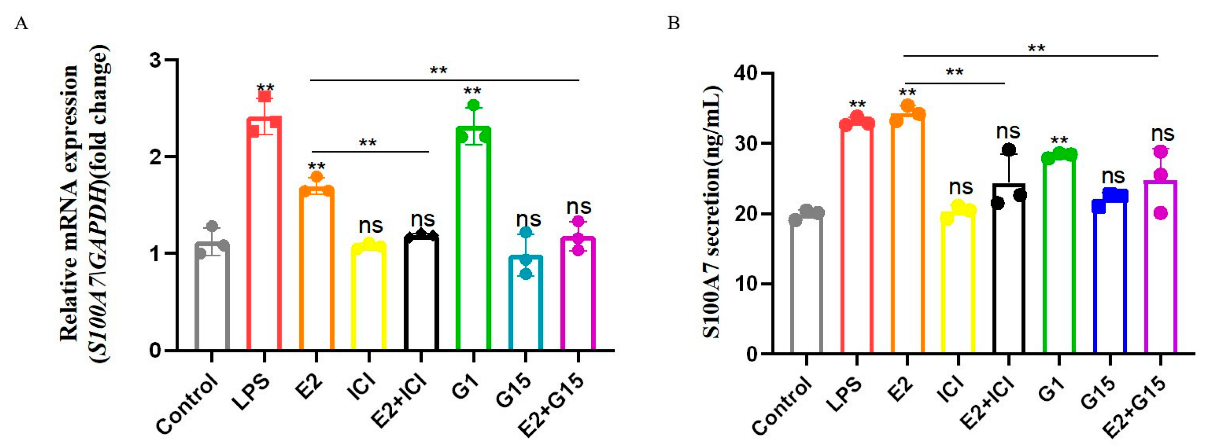
3.5. E2 Binds to Both Nuclear and Membrane Receptors to Induce Expression and Secretion of S100A7
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Degraves, F.J.; Fetrow, J. Economics of mastitis and mastitis control. Vet. Clin. N. Am. Food Anim. Pract. 1993, 9, 421–434. [Google Scholar] [CrossRef]
- Schukken, Y.; Hertl, J.; Bar, D.; Bennett, G.; González, R.; Rauch, B.; Santisteban, C.; Schulte, H.; Tauer, L.; Welcome, F.; et al. Effects of repeated gram-positive and gram-negative clinical mastitis episodes on milk yield loss in Holstein dairy cows. J. Dairy Sci. 2009, 92, 3091–3105. [Google Scholar] [CrossRef] [Green Version]
- Cha, E.; Bar, D.; Hertl, J.; Tauer, L.; Bennett, G.; González, R.; Schukken, Y.; Welcome, F.; Gröhn, Y. The cost and management of different types of clinical mastitis in dairy cows estimated by dynamic programming. J. Dairy Sci. 2011, 94, 4476–4487. [Google Scholar] [CrossRef] [Green Version]
- Seegers, H.; Fourichon, C.; Beaudeau, F. Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 2003, 34, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Erskine, R.J.; Eberhart, R.J.; Hutchinson, L.J.; Spencer, S.B.; Campbell, M.A. Incidence and types of clinical mastitis in dairy herds with high and low somatic cell counts. J. Am. Vet. Med. Assoc. 1988, 192, 761–765. [Google Scholar]
- Burton, J.L.; Madsen, S.A.; Yao, J.; Sipkovsky, S.S.; Coussens, P.M. An immunogenomics approach to understanding periparturient immunosuppression and mastitis susceptibility in dairy cows. Acta Vet. Scand. 2001, 42, 407–424. [Google Scholar] [CrossRef] [Green Version]
- Madsen, S.A.; Weber, P.S.; Burton, J.L. Altered expression of cellular genes in neutrophils of periparturient dairy cows. Vet. Immunol. Immunopathol. 2002, 86, 159–175. [Google Scholar] [CrossRef]
- Burvenich, C.; Paape, M.; Hill, A.; Guidry, A.; Miller, R.; Heyneman, R.; Kremer, W.; Brand, A. Role of the neutrophil leucocyte in the local and systemic reactions during experimentally induced e.coli mastitis in cows immediately after calving. Vet. Q. 1994, 16, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Houdijk, J.G.; Kyriazakis, I.; Jackson, F.; Huntley, J.F.; Coop, R.L. Is the allocation of metabolisable protein prioritised to milk production rather than to immune functions in Teladorsagia circumcincta-infected lactating ewes? Int. J. Parasitol. 2003, 33, 327–338. [Google Scholar] [CrossRef]
- Lamote, I.; Meyer, E.; Massart-Leën, A.M.; Burvenich, C. Sex steroids and growth factors in the regulation of mammary gland proliferation, differentiation, and involution. Steroids 2004, 69, 145–159. [Google Scholar] [CrossRef]
- Burvenich, C.; Bannerman, D.D.; Lippolis, J.D.; Peelman, L.; Nonnecke, B.J.; Kehrli, M.E., Jr.; Paape, M.J. Cumulative physiological events influence the inflammatory response of the bovine udder to Escherichia coli infections during the transition period. J. Dairy Sci. 2007, 90 (Suppl. 1), E39–E54. [Google Scholar] [CrossRef] [PubMed]
- Medina-Estrada, I.; López-Meza, J.E.; Ochoa-Zarzosa, A. Anti-Inflammatory and Antimicrobial Effects of Estradiol in Bovine Mammary Epithelial Cells during Staphylococcus aureus Internalization. Mediat. Inflamm. 2016, 2016, 6120509. [Google Scholar] [CrossRef] [Green Version]
- Lamote, I.; Meyer, E.; De Ketelaere, A.; Duchateau, L.; Burvenich, C. Expression of the estrogen receptor in blood neutrophils of dairy cows during the periparturient period. Theriogenology 2006, 65, 1082–1098. [Google Scholar] [CrossRef]
- Akers, R. Major Advances Associated with Hormone and Growth Factor Regulation of Mammary Growth and Lactation in Dairy Cows. J. Dairy Sci. 2006, 89, 1222–1234. [Google Scholar] [CrossRef]
- Patel, O.; Takenouchi, N.; Takahashi, T.; Hirako, M.; Sasaki, N.; Domeki, I. Plasma oestrone and oestradiol concentrations throughout gestation in cattle: Relationship to stage of gestation and fetal number. Res. Vet. Sci. 1999, 66, 129–133. [Google Scholar] [CrossRef]
- Gläser, R.; Harder, J.; Lange, H.; Bartels, J.; Christophers, E.; Schröder, J.-M. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 2005, 6, 57–64. [Google Scholar] [CrossRef]
- Regenhard, P.; Leippe, M.; Schubert, S.; Podschun, R.; Kalm, E.; Grötzinger, J.; Looft, C. Antimicrobial activity of bovine psoriasin. Vet. Microbiol. 2009, 136, 335–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celis, J.E.; Cruger, D.; Kiil, J.; Lauridsen, J.B.; Ratz, G.; Basse, B.; Celis, A. Identification of a group of proteins that are strongly up-regulated in total epidermal keratinocytes from psoriatic skin. FEBS Lett. 1990, 262, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Madsen, P.; Rasmussen, H.H.; Leffers, H.; Honoré, B.; Dejgaard, K.; Olsen, E.; Kiil, J.; Walbum, E.; Andersen, A.H.; Basse, B.; et al. Molecular cloning, occurrence, and expression of a novel partially secreted protein "psoriasin" that is highly up-regulated in psoriatic skin. J. Investig. Dermatol. 1991, 97, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yuliza Purba, F.; Hayashi, Y.; Nii, T.; Yoshimura, Y.; Isobe, N. Seasonal variations in the concentration of antimicrobial components in milk of dairy cows. Anim. Sci. J. 2020, 91, e13427. [Google Scholar] [CrossRef]
- Kitano, N.; Isobe, N.; Noda, J.; Takahashi, T. Concentration patterns of antibacterial factors and immunoglobulin A antibody in foremilk fractions of healthy cows. Anim. Sci. J. 2020, 91, e13372. [Google Scholar] [CrossRef]
- Elgawish, R.A.; Ogata, Y.; Hidaka, T.; Nii, T.; Yoshimura, Y.; Isobe, N. Changes in plasma concentrations of S100A7 and S100A8 in dairy cows during pregnancy. Reprod. Domest. Anim. 2018, 53, 1013–1015. [Google Scholar] [CrossRef] [PubMed]
- Tetens, J.; Friedrich, J.J.; Hartmann, A.; Schwerin, M.; Kalm, E.; Thaller, G. The spatial expression pattern of antimicrobial peptides across the healthy bovine udder. J. Dairy Sci. 2010, 93, 775–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Zhu, K.; Fan, M.; Wan, W.; Zhao, X.; Pan, M.; Ma, B.; Wei, Q. Immunolocalization of antibacterial peptide S100A7 in mastitis goat mammary gland and lipopolysaccharide induces the expression and secretion of S100A7 in goat mammary gland epithelial cells via TLR4/NF kappa B signal pathway. Anim. Biotechnol. 2022, 1–13. [Google Scholar] [CrossRef]
- Zhang, G.; Lai, S.; Yoshimura, Y.; Isobe, N. Messenger RNA expression and immunolocalization of psoriasin in the goat mammary gland and its milk concentration after an intramammary infusion of lipopolysaccharide. Vet. J. 2014, 202, 89–93. [Google Scholar] [CrossRef]
- Han, J.H.; Kim, M.S.; Lee, M.Y.; Kim, T.H.; Lee, M.K.; Kim, H.R.; Myung, S.C. Modulation of human beta-defensin-2 expression by 17beta-estradiol and progesterone in vaginal epithelial cells. Cytokine 2010, 49, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Cao, G.; Bao, T.; Cheng, L.; Li, H.; Du, C.; Tu, Y.; Li, Q.; Jian, R.; Zhao, P.; et al. Modulation of ovine SBD-1 expression by 17beta-estradiol in ovine oviduct epithelial cells. BMC Vet. Res. 2012, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Miao, Y.; Yan, Y.; Zhu, K.; Zhao, X.; Pan, M.; Ma, B.; Wei, Q. C-Type Natriuretic Peptide Regulates the Expression and Secretion of Antibacterial Peptide S100A7 in Goat Mammary Gland Through PKG/JNK/c-Jun Signaling Pathway. Front. Vet. Sci. 2022, 9, 822165. [Google Scholar] [CrossRef] [PubMed]
- Blosser, T.H. Economic losses from and the national research program on mastitis in the United States. J. Dairy Sci. 1979, 62, 119–127. [Google Scholar] [CrossRef]
- Dodd, F.H. Mastitis—Progress on control. J. Dairy Sci. 1983, 66, 1773–1780. [Google Scholar] [CrossRef]
- Koop, G.; Van Werven, T.; Toft, N.; Nielen, M. Estimating test characteristics of somatic cell count to detect Staphylococcus aureus-infected dairy goats using latent class analysis. J. Dairy Sci. 2011, 94, 2902–2911. [Google Scholar] [CrossRef]
- Boireau, C.; Cazeau, G.; Jarrige, N.; Calavas, D.; Madec, J.Y.; Leblond, A.; Haenni, M.; Gay, É. Antimicrobial resistance in bacteria isolated from mastitis in dairy cattle in France, 2006–2016. J. Dairy Sci. 2018, 101, 9451–9462. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Miramontes, C.; De Haro-Acosta, J.; Aréchiga-Flores, C.; Verdiguel-Fernández, L.; Rivas-Santiago, B. Antimicrobial peptides in domestic animals and their applications in veterinary medicine. Peptides 2021, 142, 170576. [Google Scholar] [CrossRef] [PubMed]
- Gross, S.R.; Sin, C.G.T.; Barraclough, R.; Rudland, P.S. Joining S100 proteins and migration: For better or for worse, in sickness and in health. Cell. Mol. Life Sci. 2013, 71, 1551–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.-Z.; Liu, S.-R. Koebner phenomenon leading to the formation of new psoriatic lesions: Evidences and mechanisms. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, X.; Hu, Z.; Wei, X.; Wang, Z.; Guan, T.; Liu, N.; Liu, X.; Ye, N.; Deng, G.; Luo, C.; et al. IL-37 Ameliorates the Inflammatory Process in Psoriasis by Suppressing Proinflammatory Cytokine Production. J. Immunol. 2014, 192, 1815–1823. [Google Scholar] [CrossRef] [Green Version]
- Fahey, J.V.; Wright, J.A.; Shen, L.; Smith, J.M.; Ghosh, M.; Rossoll, R.M.; Wira, C.R. Estradiol selectively regulates innate immune function by polarized human uterine epithelial cells in culture. Mucosal Immunol. 2008, 1, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Abebe, R.; Hatiya, H.; Abera, M.; Megersa, B.; Asmare, K. Bovine mastitis: Prevalence, risk factors and isolation of Staphylococcus aureus in dairy herds at Hawassa milk shed, South Ethiopia. BMC Vet. Res. 2016, 12, 270. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Wang, J.; Bu, D.; Wei, H.; Zhou, L.; Li, F.; Loor, J.J. In vitro culture and characterization of a mammary epithelial cell line from Chinese Holstein dairy cow. PLoS ONE 2009, 4, e7636. [Google Scholar] [CrossRef] [Green Version]
- Huynh, H.T.; Robitaille, G.; Turner, J.D. Establishment of bovine mammary epithelial cells (MAC-T): An in vitro model for bovine lactation. Exp. Cell Res. 1991, 197, 191–199. [Google Scholar] [CrossRef]
- Krishna, M.; Narang, H. The complexity of mitogen-activated protein kinases (MAPKs) made simple. Cell Mol. Life Sci. 2008, 65, 3525–3544. [Google Scholar] [CrossRef]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef]
- Fresno Vara, J.A.; Casado, E.; De Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.E.; Meyer, M.J.; Li, R.W.; Van Amburgh, M.E.; Boisclair, Y.R.; Capuco, A.V. Regulation of gene expression in the bovine mammary gland by ovarian steroids. J. Dairy Sci. 2007, 90 (Suppl. 1), E55–E65. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yang, Z.; Miao, Y.; Fan, M.; Zhao, X.; Wei, Q.; Ma, B. G protein-coupled estrogen receptor 1 inhibits the epithelial-mesenchymal transition of goat mammary epithelial cells via NF-κB signalling pathway. Reprod. Domest. Anim. 2021, 56, 1137–1144. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Niu, Y.; Ma, Y.; Zhao, X.; Pan, M.; Ma, B.; Wei, Q. Estradiol Regulates the Expression and Secretion of Antimicrobial Peptide S100A7 via the ERK1/2-Signaling Pathway in Goat Mammary Epithelial Cells. Animals 2022, 12, 3077. https://doi.org/10.3390/ani12223077
Yan Y, Niu Y, Ma Y, Zhao X, Pan M, Ma B, Wei Q. Estradiol Regulates the Expression and Secretion of Antimicrobial Peptide S100A7 via the ERK1/2-Signaling Pathway in Goat Mammary Epithelial Cells. Animals. 2022; 12(22):3077. https://doi.org/10.3390/ani12223077
Chicago/Turabian StyleYan, Yutong, Yuwei Niu, Yingwan Ma, Xiaoe Zhao, Menghao Pan, Baohua Ma, and Qiang Wei. 2022. "Estradiol Regulates the Expression and Secretion of Antimicrobial Peptide S100A7 via the ERK1/2-Signaling Pathway in Goat Mammary Epithelial Cells" Animals 12, no. 22: 3077. https://doi.org/10.3390/ani12223077