
Response of Pasture Nitrogen Fertilization on Greenhouse Gas Emission and Net Protein Contribution of Nellore Young Bulls
 , , ,
, , ,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. System Boundaries
2.2. Field Data Acquisition (Primary Data)
2.2.1. Experimental Design and Treatments
2.2.2. Forage Mass and Morphological Composition
2.2.3. Animal Performance
2.2.4. Intake, Digestibility, Nitrogen Balance and Enteric Methane
2.2.5. Chemical Analyses
2.2.6. Slaughter and Carcass Evaluation
2.3. Greenhouse Gas Emission

{kind=link}
{kind=link}
{kind=link}
| Source | Unit | Value | Reference |
|---|---|---|---|
| Diesel fuel use | kg·CO2e·L−1 | 3.53 | [8] |
| Electricity | kg·CO2e·MWh−1 | 115 | EPE |
| Lime | kg·CO2e·kg−1 | 0.48 | [8] |
| Fertilizer | |||
| Nitrogen (ammonium nitrate) | kg·CO2e·kg−1 | 5.50 | [26] |
| Purchased feed | |||
| Mixed mineral | kg·CO2e·kg−1 | 0.16 | [8] |
| Cottonseed meal | kg·CO2e·kg−1 | 0.91 | Feedprint 1 |
| Defatted corn germ meal | kg·CO2e·kg−1 | 0.27 | Feedprint 1 |
2.4. Beef’s Contribution to Meeting Human Protein Requirements
2.4.1. Conversion Efficiency of Beef Cattle
2.4.2. Assessing Protein Quality Using Digestible Indispensable Amino Acid Score
2.4.3. Net Protein Contribution
2.5. Statistical Analysis
3. Results
3.1. Individual Performance
3.2. GHG Emission Intensity
3.3. Beef’s Contribution to Meeting Human Protein Requirements
4. Discussion
4.1. Performance and GHG Emission
4.2. Beef’s Contribution to Meeting Human Protein Requirements
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conant, R.T. Challenges and Opportunities for Carbon Sequestration in Grassland Systems. A Technical Report on Grassland Management and Climate Change Mitigation; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2010; Volume 9, ISBN 9789251064948. [Google Scholar]
- Boval, M.; Dixon, R.M. The Importance of Grasslands for Animal Production and Other Functions: A Review on Management and Methodological Progress in the Tropics. Animal 2012, 6, 748–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. The refinement to the 2006 IPCC guidelines for national greenhouse gas inventories. Fundam. Appl. Climatol. 2019, 2, 5–13. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Muir, J.P.; Riley, D.G.; Fox, D.G. The Role of Ruminant Animals in Sustainable Livestock Intensification Programs. Int. J. Sustain. Dev. World Ecol. 2015, 22, 452–465. [Google Scholar] [CrossRef]
- Berndt, A.; Tomkins, N.W. Measurement and Mitigation of Methane Emissions from Beef Cattle in Tropical Grazing Systems: A Perspective from Australia and Brazil. Animal 2013, 7 (Suppl. 2), 363–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva Cardoso, A.; Barbero, R.P.; Romanzini, E.P.; Teobaldo, R.W.; Ongaratto, F.; da Rocha Fernandes, M.H.M.; Ruggieri, A.C.; Reis, R.A. Intensification: A Key Strategy to Achieve Great Animal and Environmental Beef Cattle Production Sustainability in Brachiaria Grasslands. Sustainability 2020, 12, 6656. [Google Scholar] [CrossRef]
- Ongaratto, F.; da Rocha Fernandes, M.H.M.; Dallantonia, E.E.; de Oliveira Lima, L.; Do Val, G.A.; da Silva Cardoso, A.; Rigobello, I.L.; Campos, J.A.A.; Reis, R.A.; Ruggieri, A.C.; et al. Intensive Production and Management of Marandu Palisadegrass (Urochloa Brizantha ‘Marandu’) Accelerates Leaf Turnover but Does Not Change Herbage Mass. Agronomy 2021, 11, 1846. [Google Scholar] [CrossRef]
- Cardoso, A.S.; Berndt, A.; Leytem, A.; Alves, B.J.R.; de Carvalho, I.d.N.O.; de Barros Soares, L.H.; Urquiaga, S.; Boddey, R.M. Impact of the Intensification of Beef Production in Brazil on Greenhouse Gas Emissions and Land Use. Agric. Syst. 2016, 143, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Dick, M.; Da Silva, M.A.; Dewes, H. Life Cycle Assessment of Beef Cattle Production in Two Typical Grassland Systems of Southern Brazil. J. Clean. Prod. 2015, 96, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Young, V.R.; Pellett, P.L. Plant Proteins in Relation to Human Protein and Amino Acid Nutrition. Am. J. Clin. Nutr. 1994, 59, 1203S–1212S. [Google Scholar] [CrossRef] [Green Version]
- Baber, J.R.; Sawyer, J.E.; Wickersham, T.A. Estimation of Human-Edible Protein Conversion Efficiency, Net Protein Contribution, and Enteric Methane Production from Beef Production in the United States. Transl. Anim. Sci. 2018, 2, 439–450. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An International Terminology for Grazing Lands and Grazing Animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Santana, S.S.; Brito, L.F.; Azenha, M.V.; Oliveira, A.A.; Malheiros, E.B.; Ruggieri, A.C.; Reis, R.A. Canopy Characteristics and Tillering Dynamics of Marandu Palisade Grass Pastures in the Rainy–Dry Transition Season. Grass Forage Sci. 2016, 72, 261–270. [Google Scholar] [CrossRef]
- RNC. Protein Requirements for Beef Cattle. In Nutrient Requirements of Zebu Beef Cattle—BR-CORTE; Suprema Gráfica LTDA: Jaú, Brazil, 2016; pp. 185–212. [Google Scholar]
- Bonnet, O.; Hagenah, N.; Hebbelmann, L.; Meuret, M.; Shrader, A.M. Is Hand Plucking an Accurate Method of Estimating Bite Mass and Instantaneous Intake of Grazing Herbivores? Rangel. Ecol. Manag. 2011, 64, 366–374. [Google Scholar] [CrossRef]
- Scarnecchia, D.L. The Animal-Unit and Animal-Unit-Equivalent Concepts in Range Science. J. Range Manag. 1985, 38, 346. [Google Scholar] [CrossRef]
- Valente, T.; Neves, P.; Detmann, E.; César De Queiroz, A.; De Campos, S.; Filho, V.; Gomes, D.I.; Figueiras, J.F. Evaluation of Ruminal Degradation Profiles of Forages Using Bags Made from Different Textiles. Rev. Bras. De Zootec. 2011, 40, 2565–2573. [Google Scholar] [CrossRef] [Green Version]
- San Vito, E.; Duarte Messana, J.; Frighetto, R.; Andrade Reis, R. Performance and Methane Emissions of Grazing Nellore Bulls Supplemented with Crude Glycerin. J. Anim. Sci. 2016, 94, 4728–4737. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.; Rotta, P.P.; Costa, E.; Silva, L.F.; Menezes, A.C.B.; Pina, D.S.; Valadares Filho, S.C. Protein Ruminal Degradation of Feeds and Microbial Protein Synthesis; UFV: Viçosa, Brazil, 2016; ISBN 978-85-8179-111-1. [Google Scholar]
- Berndt, A.; Boland, T.M.; Deighton, M.H.; Gere, J.I.; Grainger, C.; Hegarty, R.S.; Iwaasa, A.D.; Koolaard, J.P.; Lassey, K.R.; Luo, D.; et al. Guidelines for Use of Sulphur Hexafluoride (SF6) Tracer Technique to Measure Enteric Methane Emissions from Ruminants; Ministry for Primary Industries: Wellington, New Zealand, 2014; ISBN 9780478432107. [Google Scholar]
- Berça, A.S.; da Silva Cardoso, A.; Longhini, V.Z.; Tedeschi, L.O.; Boddey, R.M.; Berndt, A.; Reis, R.A.; Ruggieri, A.C. Methane Production and Nitrogen Balance of Dairy Heifers Grazing Palisade Grass Cv. Marandu Alone or with Forage Peanut. J. Anim. Sci. 2019, 97, 4625–4634. [Google Scholar] [CrossRef]
- Townshend, A. Official Methods of Analysis of the Association of Official Analytical Chemists, 14th Edn. Anal. Chim. Acta 1987, 193, 400–401. [Google Scholar] [CrossRef]
- da Silva Cardoso, A.; Oliveira, S.C.; Janusckiewicz, E.R.; Brito, L.F.; da Silva Morgado, E.; Reis, R.A.; Ruggieri, A.C. Seasonal Effects on Ammonia, Nitrous Oxide, and Methane Emissions for Beef Cattle Excreta and Urea Fertilizer Applied to a Tropical Pasture. Soil Tillage Res. 2019, 194, 104341. [Google Scholar] [CrossRef]
- Vellinga, T.V.; Blonk, H.; Marinussen, M.; Van Zeist, W.J.; De Boer, I.J.M.; Starmans, D. Methodology Used in Feedprint: A Tool Quantifying Greenhouse Gas Emissions of Feed Production and Utilization; Wageningen UR Livestock Research: Wageningen, The Netherlands, 2013. [Google Scholar]
- Intergovernmental Panel on Climate Change. In Encyclopedia of Corporate Social Responsibility; Springer: Berlin/Heidelberg, Germany, 2013; p. 1492.
- Ledgard, S.F.; Boyes, M.; Brentrup, F. Life Cycle Assessment of Local and Imported Fertilisers Used on New Zealand Farms; Ministry of Agriculture and Forestry: Wellington, New Zealand, 2011. [Google Scholar]
- Gionbelli, M.P. Adjusting Cattle Body Weight to Physiological and Feeding Conditions Mateus Pies Gionbelli, Sebastião de Campos Valadares Filho, Edenio Detmann; Suprema Gráfica Ltd.: Viçosa, Brazil, 2016. [Google Scholar]
- Apple, J.K.; Davis, J.C.; Stephenson, J. Influence of Body Condition Score on By-Product Yield and Value from Cull Beef Cows. J. Anim. Sci. 1999, 77, 2670–2679. [Google Scholar] [CrossRef]
- Wilkinson, J.M. Re-Defining Efficiency of Feed Use by Livestock. Animal 2011, 5, 1014–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertl, P.; Steinwidder, A.; Schönauer, M.; Krimberger, K.; Knaus, W.; Zollitsch, W. Net Food Production of Different Livestock: A National Analysis for Austria Including Relative Occupation of Different Land Categories. Bodenkultur 2016, 67, 91–103. [Google Scholar] [CrossRef] [Green Version]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition Report of an FAO Expert Consultation; FAO: Rome, Italy, 2011. [Google Scholar]
- Blok, M.; Spek, J. CVB Veevoedertabel 2019: Chemische Samenstelling en Nutritionele Waarden van Voedermiddelen; Federatie Nederlandse Diervoederketen: Rotterdam, The Netherlands, 2019; pp. 1–614. [Google Scholar]
- Delevatti, L.M.; Romanzini, E.P.; Koscheck, J.F.W.; da Ross de Araujo, T.L.; Renesto, D.M.; Ferrari, A.C.; Barbero, R.P.; Mulliniks, J.T.; Reis, R.A. Forage Management Intensification and Supplementation Strategy: Intake and Metabolic Parameters on Beef Cattle Production. Anim. Feed Sci. Technol. 2019, 247, 74–82. [Google Scholar] [CrossRef]
- Dumortier, J.; Hayes, D.J.; Carriquiry, M.; Dong, F.; Du, X.; Elobeid, A.; Fabiosa, J.F.; Mulik, K. Modeling the Effects of Pasture Expansion on Emissions from Land-Use Change. In Proceedings of the 2010: Climate Change in World Agriculture: Mitigation, Adaptation, Trade and Food Security, Stuttgart-Hohenheim, Germany, 27–29 June 2010. [Google Scholar]
- Ruggieri, A.C.; da Silva Cardoso, A.; Ongaratto, F.; Casagrande, D.R.; Barbero, R.P.; Brito, L.d.F.; Azenha, M.V.; Oliveira, A.A.; Koscheck, J.F.W.; Reis, R.A. Grazing Intensity Impacts on Herbage Mass, Sward Structure, Greenhouse Gas Emissions, and Animal Performance: Analysis of Brachiaria Pastureland. Agronomy 2020, 10, 1750. [Google Scholar] [CrossRef]
- Uzun, P.; Masucci, F.; Serrapica, F.; Varricchio, M.L.; Pacelli, C.; Claps, S.; Di Francia, A. Use of Mycorrhizal Inoculum under Low Fertilizer Application: Effects on Forage Yield, Milk Production, and Energetic and Economic Efficiency. J. Agric. Sci. 2018, 156, 127–135. [Google Scholar] [CrossRef]
- Weiss, F.; Leip, A. Greenhouse Gas Emissions from the EU Livestock Sector: A Life Cycle Assessment Carried out with the CAPRI Model. Agric. Ecosyst. Environ. 2012, 149, 124–134. [Google Scholar] [CrossRef]
- Molossi, L.; Hoshide, A.K.; Pedrosa, L.M.; de Oliveira, A.S.; de Abreu, D.C. Improve Pasture or Feed Grain? Greenhouse Gas Emissions, Profitability, and Resource Use for Nelore Beef Cattle in Brazil’s Cerrado and Amazon Biomes. Animals 2020, 10, 1386. [Google Scholar] [CrossRef]
- ABIEC. Beef Report 2020—ABIEC. Available online: https://www.abiec.com.br/publicacoes/beef-report-2020/ (accessed on 13 September 2022).
- de Figueiredo, E.B.; Jayasundara, S.; de Oliveira Bordonal, R.; Berchielli, T.T.; Reis, R.A.; Wagner-Riddle, C.; La Scala, N. Greenhouse Gas Balance and Carbon Footprint of Beef Cattle in Three Contrasting Pasture-Management Systems in Brazil. J. Clean. Prod. 2017, 142, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Smith, P.; Haberl, H.; Montzka, S.A.; McAlpine, C.; Boucher, D.H. Ruminants, Climate Change and Climate Policy. Nat. Clim. Chang. 2014, 4, 2–5. [Google Scholar] [CrossRef]
- Mc Geough, E.J.; Little, S.M.; Janzen, H.H.; McAllister, T.A.; McGinn, S.M.; Beauchemin, K.A. Life-Cycle Assessment of Greenhouse Gas Emissions from Dairy Production in Eastern Canada: A Case Study. J. Dairy Sci. 2012, 95, 5164–5175. [Google Scholar] [CrossRef]
- da Cruz Corrêa, D.C.; da Silva Cardoso, A.; Ferreira, M.R.; Siniscalchi, D.; De Almeida Gonçalves, P.H.; Lumasini, R.N.; Reis, R.A.; Ruggieri, A.C. Ammonia Volatilization, Forage Accumulation, and Nutritive Value of Marandu Palisade Grass Pastures in Different n Sources and Doses. Atmosphere 2021, 12, 1179. [Google Scholar] [CrossRef]
- Bustamante, M.M.C.; Nobre, C.A.; Smeraldi, R.; Aguiar, A.P.D.; Barioni, L.G.; Ferreira, L.G.; Longo, K.; May, P.; Pinto, A.S.; Ometto, J.P.H.B. Estimating Greenhouse Gas Emissions from Cattle Raising in Brazil. Clim. Chang. 2012, 115, 559–577. [Google Scholar] [CrossRef]
- Assad, E.D.; Pinto, H.S.; Martins, S.C.; Groppo, J.D.; Salgado, P.R.; Evangelista, B.; Vasconcellos, E.; Sano, E.E.; Pavão, E.; Luna, R.; et al. Changes in Soil Carbon Stocks in Brazil Due to Land Use: Paired Site Comparisons and a Regional Pasture Soil Survey. Biogeosciences 2013, 10, 6141–6160. [Google Scholar] [CrossRef] [Green Version]
- Picasso, V.D.; Modernel, P.D.; Becoña, G.; Salvo, L.; Gutiérrez, L.; Astigarraga, L. Sustainability of Meat Production beyond Carbon Footprint: A Synthesis of Case Studies from Grazing Systems in Uruguay. Meat Sci. 2014, 98, 346–354. [Google Scholar] [CrossRef]
- Johnston, A.E.; Poulton, P.R.; Coleman, K. Chapter 1 Soil Organic Matter: Its Importance in Sustainable Agriculture and Carbon Dioxide Fluxes. Adv. Agron. 2009, 101, 1–57. [Google Scholar] [CrossRef]
- Ertl, P.; Zebeli, Q.; Zollitsch, W.; Knaus, W. Feeding of By-Products Completely Replaced Cereals and Pulses in Dairy Cows and Enhanced Edible Feed Conversion Ratio. J. Dairy Sci. 2015, 98, 1225–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottet, A.; de Haan, C.; Falcucci, A.; Tempio, G.; Opio, C.; Gerber, P. Livestock: On Our Plates or Eating at Our Table? A New Analysis of the Feed/Food Debate. Glob. Food Secur. 2017, 14, 1–8. [Google Scholar] [CrossRef]
| Ingredient | Composition (%DM) | ||||
|---|---|---|---|---|---|
| Supplement Backgrounding 1 | Supplement Finishing 2 | TMR 3 | HEF 4 | DIASS 5 | |
| Sorghum silage | - | - | 20.0 | 0 | |
| Defatted corn germ | 21.9 | 94.4 | 75.5 | 0.5 | 34.8 |
| Cotton seed meal | 37.0 | - | - | 0 | |
| Urea | 4.6 | 1.8 | 1.5 | 0 | |
| Mineral mix | 36.5 | 3.8 | 3.0 | 0 | |
| Production Systems 1 | |||||
|---|---|---|---|---|---|
| Variable 2 | P0N + PS | P75N + F | P150N + F | SEM | p |
| Backgrounding | |||||
| Days, d | 172 | 172 | 172 | - | - |
| Avg. initial SBW, kg | 273 | 282 | 294 | 15.60 | 0.650 |
| Avg. SBW at end, kg | 385 | 423 | 437 | 20.00 | 0.210 |
| Avg. daily gain, kg d−1 | 0.62 b | 0.81 a | 0.84 a | 0.030 | 0.001 |
| Dry matter intake, %BW | 2.4 | 2.5 | 2.5 | 0.230 | 0.970 |
| Dry matter digestibility, % | 50.5 | 55.3 | 50.1 | 2.910 | 0.410 |
| Crude protein digestibility, % | 45.2 b | 57.3 a | 55.9 a | 2.870 | 0.028 |
| Individual enteric methane, g d−1 | 141.2 | 173.3 | 179.6 | 16.360 | 0.260 |
| Finishing | |||||
| Days, d | |||||
| Avg. daily gain, kg d−1 | 0.92 | 1.01 | 1 | 0.074 | 0.640 |
| Dry matter intake, %BW | 2.8 a | 2.4 a,b | 2.3 b | 0.130 | 0.046 |
| Dry matter digestibility, % | 58.7 | 61.9 | 60.2 | 1.820 | 0.450 |
| Protein digestibility, % | 61 | 63.8 | 60.7 | 4.250 | 0.820 |
| Individual enteric methane, g d−1 | 135.0 b | 218.6 a | 224.5 a | 22.20 | 0.033 |
| Avg. slaughter SBW, kg | 460 | 504 | 510 | 17.10 | 0.130 |
| Carcass weight, kg | 265 b | 300 a | 304 a | 10.80 | 0.055 |
| Dressing percentage, % | 59.9 | 59.3 | 59.4 | 0.600 | 0.750 |
| Production Systems 1 | |||||
|---|---|---|---|---|---|
| Variable 2 | P0N + PS | P75N + F | P150N + F | SEM | p |
| Backgrounding | |||||
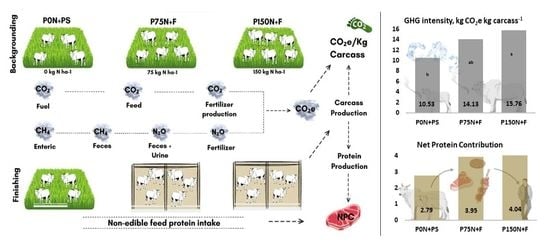
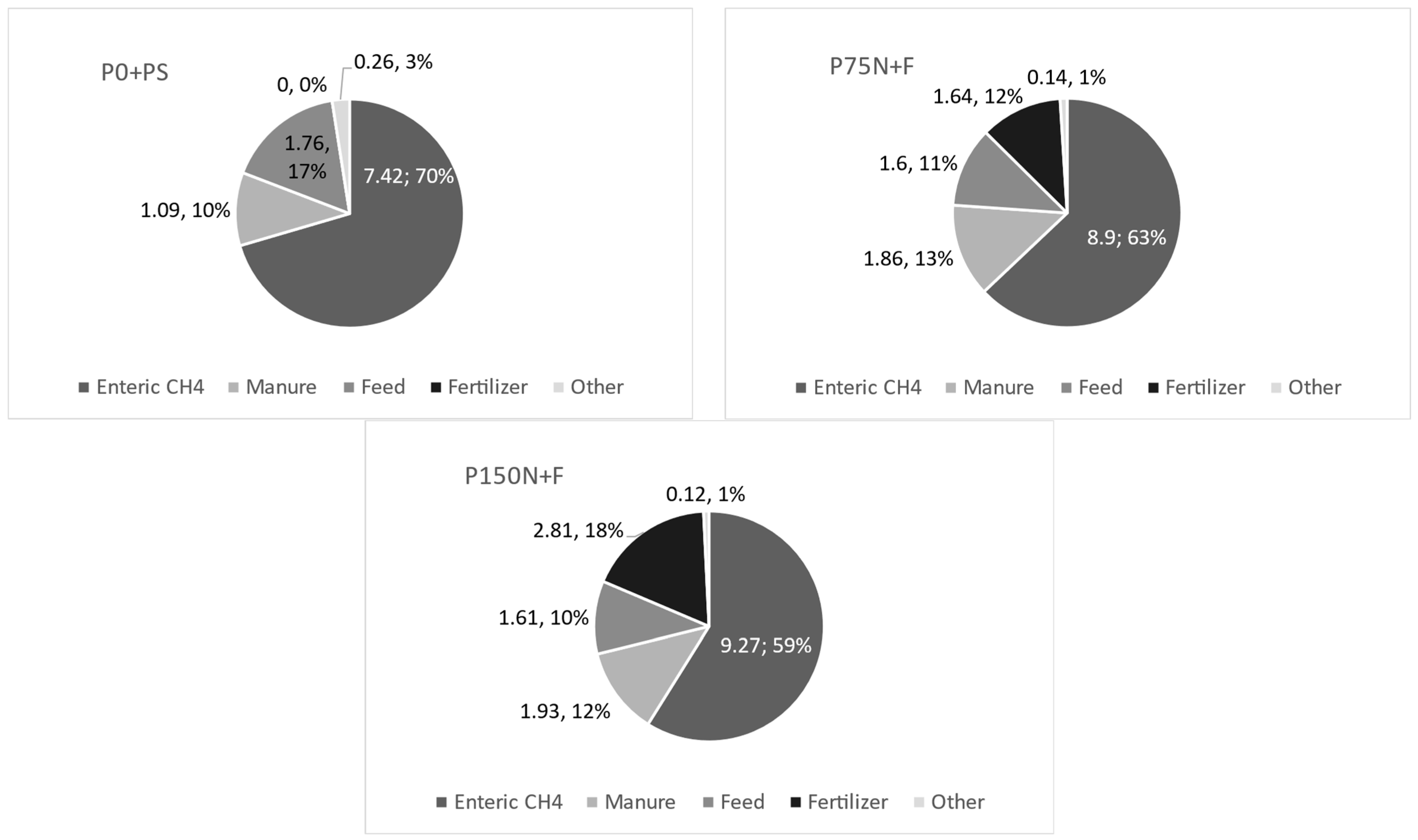
| GHG total, kg CO2e ha−1 | 1994 b | 4634 a | 6134 a | 397 | 0.001 |
| Carcass production, kg ha−1 | 134 b | 272 a | 311 a | 21.1 | 0.001 |
| GHG intensity, kg CO2e kg carcass−1 | 5.87 b | 7.32 a,b | 8.35 a | 0.522 | 0.025 |
| Finishing | |||||
| GHG total, kg CO2e ha−1 | 1580 b | 4429 a | 5363 a | 582 | 0.0034 |
| Carcass production, kg ha−1 | 203 b | 366 a | 423 a | 35.8 | 0.005 |
| GHG intensity, kg CO2e kg carcass−1 | 4.65 b | 6.80 a,b | 7.40 a | 0.652 | 0.036 |
| Entire period | |||||
| GHG total, kg CO2e ha−1 | 3574 b | 9063 a | 11,498 a | 838 | 0.0003 |
| Carcass production, kg ha−1 | 337 b | 637 a | 734 a | 45.8 | 0.0004 |
| GHG intensity, kg CO2e kg carcass−1 | 10.53 b | 14.13 a,b | 15.76 a | 0.915 | 0.008 |
| Production Systems 1 | |||||
|---|---|---|---|---|---|
| Variable 2 | P0N + PS | P75N + F | P150N + F | SEM | p |
| Backgrounding | |||||
| Diet DIAAS | 34.8 | 34.8 | 34.8 | ||
| PQR | 3.2 | 3.22 | 3.22 | ||
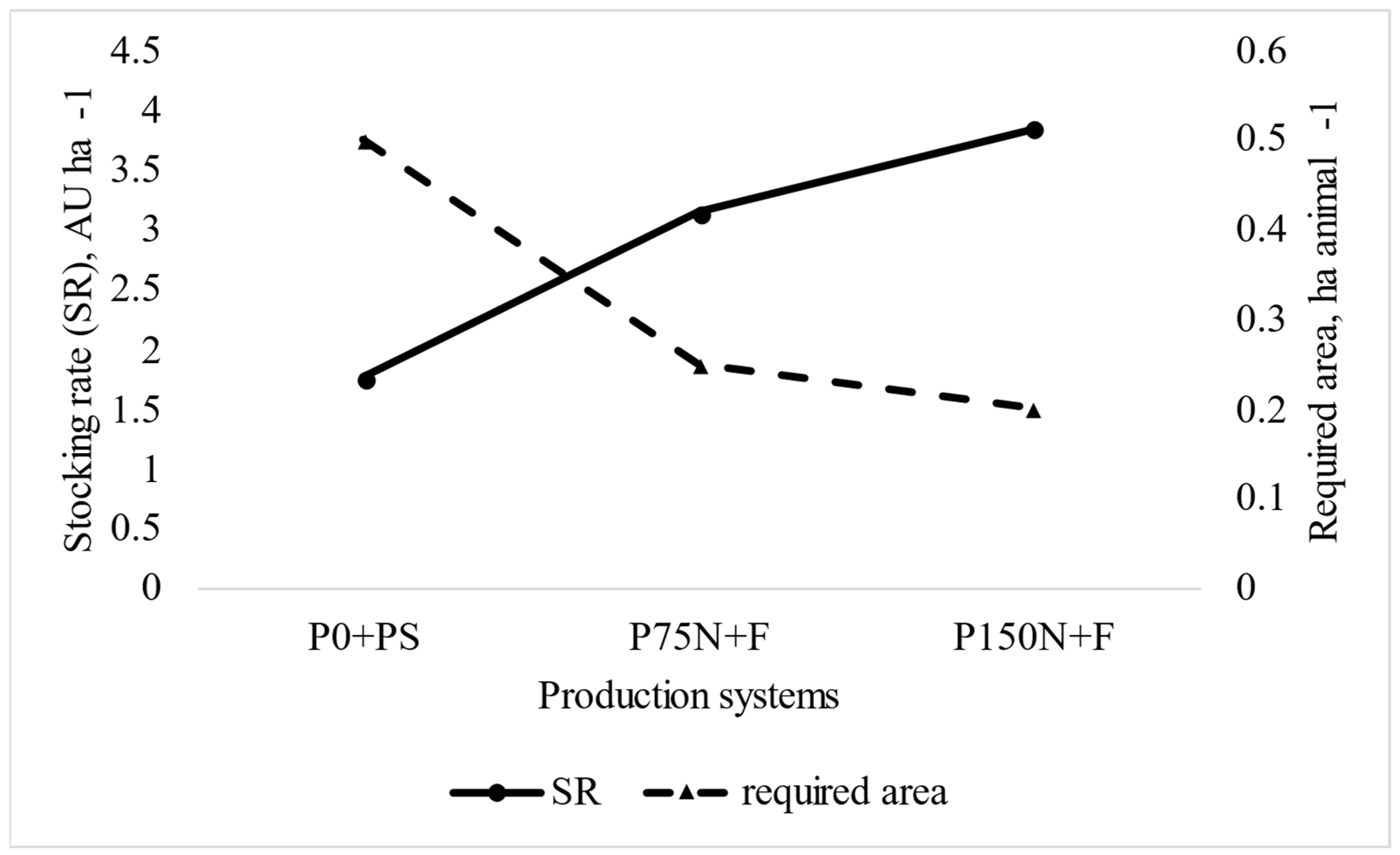
| Total HePf, kg paddock−1 | 0.6 b | 1.16 a | 1.31 a | 0.11 | 0.004 |
| Total HePp, kg paddock−1 | 55.2 b | 118.3 a | 129.4 a | 8.53 | <0.001 |
| HePCE | 90.8 | 103.7 | 99.4 | 5.06 | 0.24 |
| NPC | 212.7 | 242.7 | 232.7 | 11.84 | 0.24 |
| Finishing | |||||
| Diet DIAAS | 34.8 | 34.8 | 34.8 | ||
| PQR | 3.2 | 3.22 | 3.22 | ||
| Total HePf, kg paddock−1 | 98.9 | 162.1 | 162.9 | 23.58 | 0.14 |
| Total HePp, kg paddock−1 | 36.7 b | 68.6 a | 70.2 a | 7.47 | 0.019 |
| HePCE | 0.4 | 0.5 | 0.5 | 0.059 | 0.51 |
| NPC | 1.1 | 1.4 | 1.4 | 0.18 | 0.51 |
| Entire period | |||||
| Diet DIAAS | 34.8 | 34.8 | 34.8 | ||
| PQR | 3.22 | 3.22 | 3.22 | ||
| Total HePf, kg paddock−1 | 99.5 | 163.2 | 164.2 | 23.6 | 0.14 |
| Total HePp, kg paddock−1 | 91.9 b | 186.9 a | 199.6 a | 12.8 | <0.001 |
| HePCE | 0.92 | 1.31 | 1.34 | 0.24 | 0.45 |
| NPC | 2.79 | 3.95 | 4.04 | 0.74 | 0.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, L.; Ongaratto, F.; Fernandes, M.; Cardoso, A.; Lage, J.; Silva, L.; Reis, R.; Malheiros, E. Response of Pasture Nitrogen Fertilization on Greenhouse Gas Emission and Net Protein Contribution of Nellore Young Bulls. Animals 2022, 12, 3173. https://doi.org/10.3390/ani12223173
Lima L, Ongaratto F, Fernandes M, Cardoso A, Lage J, Silva L, Reis R, Malheiros E. Response of Pasture Nitrogen Fertilization on Greenhouse Gas Emission and Net Protein Contribution of Nellore Young Bulls. Animals. 2022; 12(22):3173. https://doi.org/10.3390/ani12223173
Chicago/Turabian StyleLima, Lais, Fernando Ongaratto, Marcia Fernandes, Abmael Cardoso, Josiane Lage, Luis Silva, Ricardo Reis, and Euclides Malheiros. 2022. "Response of Pasture Nitrogen Fertilization on Greenhouse Gas Emission and Net Protein Contribution of Nellore Young Bulls" Animals 12, no. 22: 3173. https://doi.org/10.3390/ani12223173
APA StyleLima, L., Ongaratto, F., Fernandes, M., Cardoso, A., Lage, J., Silva, L., Reis, R., & Malheiros, E. (2022). Response of Pasture Nitrogen Fertilization on Greenhouse Gas Emission and Net Protein Contribution of Nellore Young Bulls. Animals, 12(22), 3173. https://doi.org/10.3390/ani12223173