

Locomotory Profiles in Thoroughbreds: Peak Stride Length and Frequency in Training and Association with Race Outcomes

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Databases
- (1)
- racehorse training sessions: collected by means of a fitness tracker (the ‘Equimetre’™) from a single racing yard (Ciaron Maher Racing) in Victoria, Australia
- (2)
- racehorse pedigree information: publicly available and downloaded from https://www.pedigreequery.com (accessed 4 August 2022)
- (3)
- race results: available upon subscription in Australia, with race data recorded and downloaded from http://www.racing.com (accessed 7 July 2022)
2.2. Designation of Racehorses according to Race Distance
2.3. Training Data
2.4. Pedigree Data
2.5. Statistical Analysis
3. Results
3.1. Racing Data and Performance
3.2. Training Data and Locomotory Performance
3.3. Training Data, Type of Racehorse and Predicting Race Outcome
3.4. Heritability of Stride Parameters and Peak Heart Rate
4. Discussion
4.1. Racing Data and Performance
4.2. Training Data and Locomotory Performance
4.3. Training Data, Type of Racehorse and Predicting Race Outcome
4.4. Heritability of Stride Parameters and Peak Heart Rate
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tozaki, T.; Sato, F.; Hill, E.W.; Miyake, T.; Endo, Y.; Kakoi, H.; Gawahara, H.; Hirota, K.-I.; Nakano, Y.; Nambo, Y. Sequence variants at the myostatin gene locus influence the body composition of Thoroughbred horses. J. Vet. Med. Sci. 2011, 73, 1108030597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellander, J.; Fredricson, I.; Hertén, G.; Drevemo, S.; Dalin, G. Galloppaktion I—Basala gangartsvariabler i relation till hästens hastighet. Sven. Vet. 1983, 35, 75–82. [Google Scholar]
- Schrurs, C.; Dubois, G.; Patarin, F.; Cobb, M.; Gardner, D.S.; Van Erck-Westergren, E. Cardiovascular and locomotory parameters during training in Thoroughbred racehorses: A multi-national study. Comp. Exerc. Physiol. 2022, 18, 185–199. [Google Scholar] [CrossRef]
- Hill, E.W.; Gu, J.; Eivers, S.S.; Fonseca, R.G.; McGivney, B.A.; Govindarajan, P.; Orr, N.; Katz, L.M.; MacHugh, D. A sequence polymorphism in MSTN predicts sprinting ability and racing stamina in thoroughbred horses. PLoS ONE 2010, 5, e8645. [Google Scholar] [CrossRef]
- Kusy, K.; Zielinski, J. Sprinters versus long-distance runners: How to grow old healthy. Exerc. Sport Sci. Rev. 2015, 43, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M. Physiological and biomechanical mechanisms of distance specific human running performance. Integr. Comp. Biol. 2017, 57, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, A.S.; Morrice-West, A.V.; Whitton, R.C.; Hitchens, P.L. Changes in Thoroughbred speed and stride characteristics over successive race starts and their association with musculoskeletal injury. Equine Vet. J. 2022, in press. [CrossRef]
- Binns, M.; Boehler, D.; Lambert, D. Identification of the myostatin locus (MSTN) as having a major effect on optimum racing distance in the Thoroughbred horse in the USA. Anim. Genet. 2010, 41, 154–158. [Google Scholar] [CrossRef]
- Tozaki, T.; Miyake, T.; Kakoi, H.; Gawahara, H.; Sugita, S.; Hasegawa, T.; Ishida, N.; Hirota, K.; Nakano, Y. A genome-wide association study for racing performances in Thoroughbreds clarifies a candidate region near the MSTN gene. Anim. Genet. 2010, 41, 28–35. [Google Scholar] [CrossRef]
- Clayton, H.M. Horse species symposium: Biomechanics of the exercising horse. J. Anim. Sci. 2016, 94, 4076–4086. [Google Scholar] [CrossRef]
- Deuel, N.R.; Park, J.-J. The gait patterns of Olympic dressage horses. J. Appl. Biomech. 1990, 6, 198–226. [Google Scholar] [CrossRef]
- Saastamoinen, M.T.; Barrey, E. Physiological Traits. In The Genetics of the Horse; CAB International: Cambridge, MA, USA, 2000; p. 439. [Google Scholar]
- Schrurs, C.; Dubois, G.; Van Erck-Westergren, E.; Gardner, D.S. Does sex of the jockey influence racehorse physiology and performance. PLoS ONE 2022, 17, e0273310. [Google Scholar] [CrossRef] [PubMed]
- Ter Woort, F.; Dubois, G.; Didier, M.; Van Erck-Westergren, E. Validation of an equine fitness tracker: Heart rate and heart rate variability. Comp. Exerc. Physiol. 2021, 17, 189–198. [Google Scholar] [CrossRef]
- Velie, B.; Wade, C.; Hamilton, N. Profiling the careers of Thoroughbred horses racing in Australia between 2000 and 2010. Equine Vet. J. 2013, 45, 182–186. [Google Scholar] [CrossRef]
- Huggins, M. Flat Racing and British Society 1790–1914: A Social and Economic History; Routledge: London, UK, 2014. [Google Scholar]
- Farries, G.; McGettigan, P.; Gough, K.; McGivney, B.; MacHugh, D.; Katz, L.; Hill, E. Genetic contributions to precocity traits in racing Thoroughbreds. Anim. Genet. 2018, 49, 193–204. [Google Scholar] [CrossRef]
- Hill, E.W.; Ryan, D.P.; MacHugh, D.E. Horses for courses: A DNA-based test for race distance aptitude in thoroughbred racehorses. Recent Pat. DNA Gene Seq. Discontin. 2012, 6, 203–208. [Google Scholar] [CrossRef]
- Butler, J.A.; Colles, C.M.; Dyson, S.J.; Kold, S.E.; Poulos, P.W. Clinical Radiology of the Horse; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]
- Racing Australia. Australian Group Races. Available online: http://www.racingaustralia.com.au (accessed on 1 August 2022).
- Horse Racing Ireland. Irish Pattern Races, Listed Races and Premier Handicaps. 2017. Available online: https://www.goracing.ie/ (accessed on 3 August 2022).
- British Horseracing Authority. UK Flat Pattern. 2017. Available online: https://www.britishhorseracing.com (accessed on 1 September 2022).
- Rooney, M.F.; Hill, E.W.; Kelly, V.P.; Porter, R.K. The “speed gene” effect of myostatin arises in Thoroughbred horses due to a promoter proximal SINE insertion. PLoS ONE 2018, 13, e0205664. [Google Scholar] [CrossRef] [Green Version]
- Webb-Carter, C. A Study into British Stayers and Staying Races; The Thoroughbred Breeders’ Association: Newmarket, UK, 2015. [Google Scholar]
- Sobczynska, M. Genetic correlations between racing performance at different racing distances in Thoroughbreds and Arab horses. Czech J. Anim. Sci. 2006, 51, 523. [Google Scholar] [CrossRef] [Green Version]
- Gramm, M.; Marksteiner, R. The effect of age on thoroughbred racing performance. J. Equine Sci. 2010, 21, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Tranquille, C.; Walker, V.; Hodgins, D.; McEwen, J.; Roberts, C.; Harris, P.; Cnockaert, R.; Guire, R.; Murray, R. Quantification of warm-up patterns in elite showjumping horses over three consecutive days: A descriptive study. Comp. Exerc. Physiol. 2017, 13, 53–61. [Google Scholar] [CrossRef]
- Johnston, C.; Gottlieb-Vedi, M.; Drevemo, S.; Roepstorff, L. The kinematics of loading and fatigue in the Standardbred trotter. Equine Vet. J. 1999, 31, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Wickler, S.; Greene, H.; Egan, K.; Astudillo, A.; Dutto, D.; Hoyt, D. Stride parameters and hindlimb length in horses fatigued on a treadmill and at an endurance ride. Equine Vet. J. 2006, 38, 60–64. [Google Scholar] [CrossRef] [PubMed]
- The Rules of Racing of Racing Victoria. Available online: https://www.racingvictoria.com.au/the-sport/racing/~/media/1c929fd13ebf4ea998c579b5333fd438.ashx (accessed on 1 August 2022).
- Gaffney, B.; Cunningham, E. Estimation of genetic trend in racing performance of thoroughbred horses. Nature 1988, 332, 722–724. [Google Scholar] [CrossRef]
- Hintz, H.F. Nutrition and equine performance. J. Nutr. 1994, 124, 2723S–2729S. [Google Scholar] [PubMed]
- Morrice-West, A.V.; Hitchens, P.L.; Walmsley, E.A.; Wong, A.S.; Whitton, R.C. Association of Thoroughbred Racehorse Workloads and Rest Practices with Trainer Success. Animals 2021, 11, 3130. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.; Vamplew, W. Encyclopedia of British Horse Racing; Routledge: London, UK, 2012. [Google Scholar]
- Hintz, R.L. Genetics of performance in the horse. J. Anim. Sci. 1980, 51, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Kearns, C.; McKeever, K.H.; Abe, T. Overview of horse body composition and muscle architecture: Implications for performance. Vet. J. 2002, 164, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Rivero, J.L. A scientific background for skeletal muscle conditioning in equine practice. J. Vet. Med. Ser. A 2007, 54, 321–332. [Google Scholar] [CrossRef]
- Dolvik, N.; Klemetsdal, G. Conformational traits of Norwegian cold-blooded trotters: Heritability and the relationship with performance. Acta Agric. Scand. Sect. A-Anim. Sci. 1999, 49, 156–162. [Google Scholar] [CrossRef]
- Galisteo, A.; Cano, M.; Morales, J.; Vivo, J.; Miró, F. The influence of speed and height at the withers on the kinematics of sound horses at the hand-led trot. Vet. Res. Commun. 1998, 22, 415–424. [Google Scholar] [CrossRef]
- Leach, D.; Cymbaluk, N.F. Relationships between stride length, stride frequency, velocity, and morphometrics of foals. Am. J. Vet. Res. 1986, 47, 2090–2097. [Google Scholar] [PubMed]
- Drevemo, S.; Dalin, G.; Fredricson, I.; Björne, K. Equine locomotion: 3. The reproducibility of gait in Standardbred trotters. Equine Vet. J. 1980, 12, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Miró, F.; Santos, R.; Garrido-Castro, J.; Galisteo, A.; Medina-Carnicer, R. 2D versus 3D in the kinematic analysis of the horse at the trot. Vet. Res. Commun. 2009, 33, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Barrey, E.; Auvinet, B.; Couroucé, A. Gait evaluation of race trotters using an accelerometric device. Equine Vet. J. 1995, 27, 156–160. [Google Scholar] [CrossRef]
- Weatherby, S. An Introduction to a General Stud Book; Weatherby and Sons: London, UK, 1791. [Google Scholar]
- Cunningham, E.; Dooley, J.; Splan, R.; Bradley, D. Microsatellite diversity, pedigree relatedness and the contributions of founder lineages to thoroughbred horses. Anim. Genet. 2001, 32, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Bower, M.A.; Campana, M.; Whitten, M.; Edwards, C.J.; Jones, H.; Barrett, E.; Cassidy, R.; Nisbet, R.; Hill, E.; Howe, C. The cosmopolitan maternal heritage of the Thoroughbred racehorse breed shows a significant contribution from British and Irish native mares. Biol. Lett. 2011, 7, 316–320. [Google Scholar] [CrossRef] [Green Version]
- Wallner, B.; Palmieri, N.; Vogl, C.; Rigler, D.; Bozlak, E.; Druml, T.; Jagannathan, V.; Leeb, T.; Fries, R.; Tetens, J. Y chromosome uncovers the recent oriental origin of modern stallions. Curr. Biol. 2017, 27, 2029–2035.e25. [Google Scholar] [CrossRef] [Green Version]
- Langlois, B. Heritability of racing ability in Thoroughbreds—A review. Livest. Prod. Sci. 1980, 7, 591–605. [Google Scholar] [CrossRef]
- Barrey, E.; Desliens, F.; Blouin, C.; Langlois, B. Heritabilities of gait characteristics: Application for breeding in dressage. In Proceedings of the Conference on Equine Sports Medicine and Science 2002, the Elite Dressage and Three-Day-Event Horse, Saumur, France, 19–21 October 2002. [Google Scholar]
- Couroucé, A.; Chrétien, M.; Valette, J. Physiological variables measured under field conditions according to age and state of training in French Trotters. Equine Vet. J. 2002, 34, 91–97. [Google Scholar] [CrossRef]
- Betros, C.; McKeever, K.; Kearns, C.; Malinowski, K. Effects of ageing and training on maximal heart rate and VO2max. Equine Vet. J. 2002, 34, 100–105. [Google Scholar] [CrossRef]
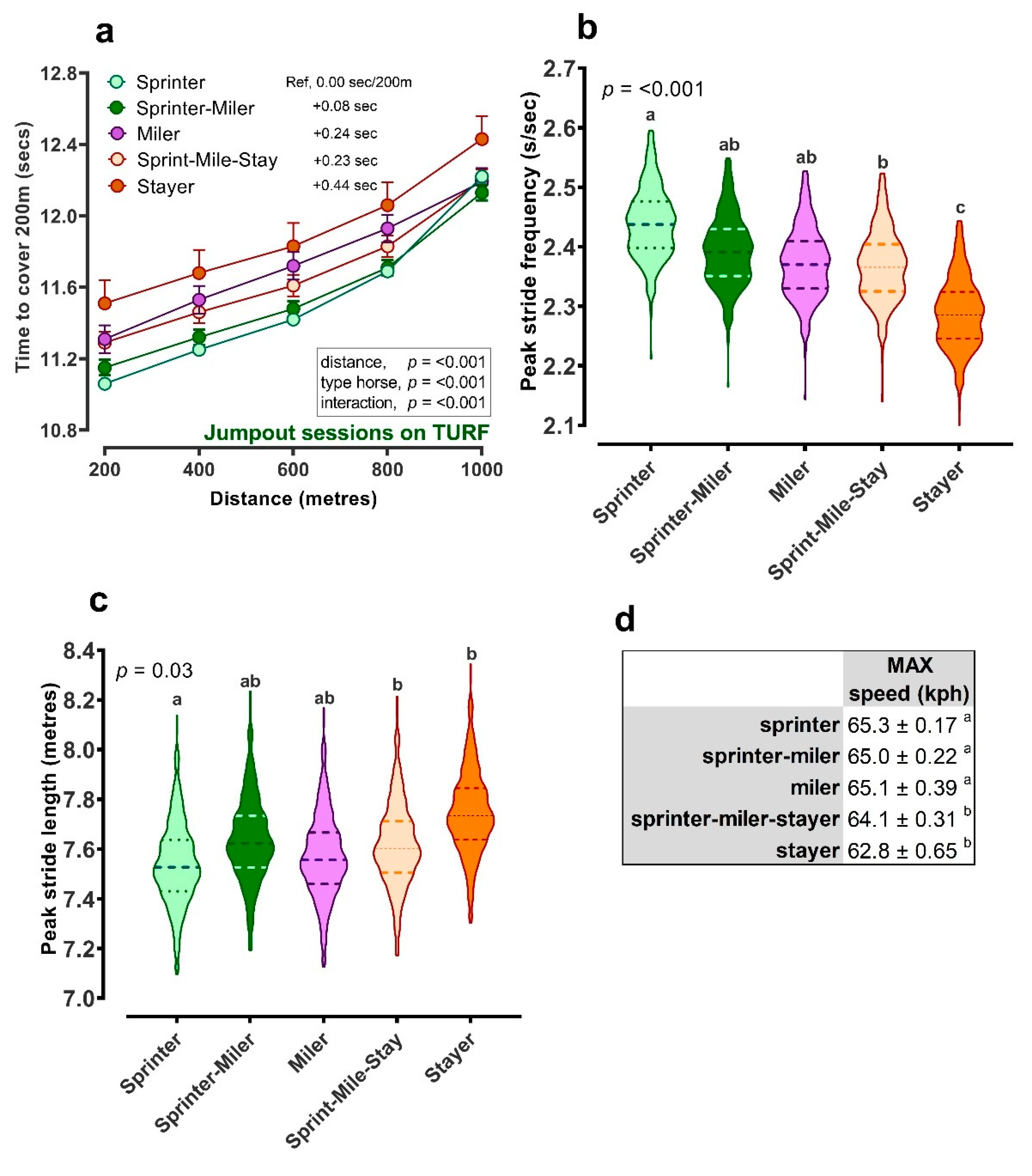
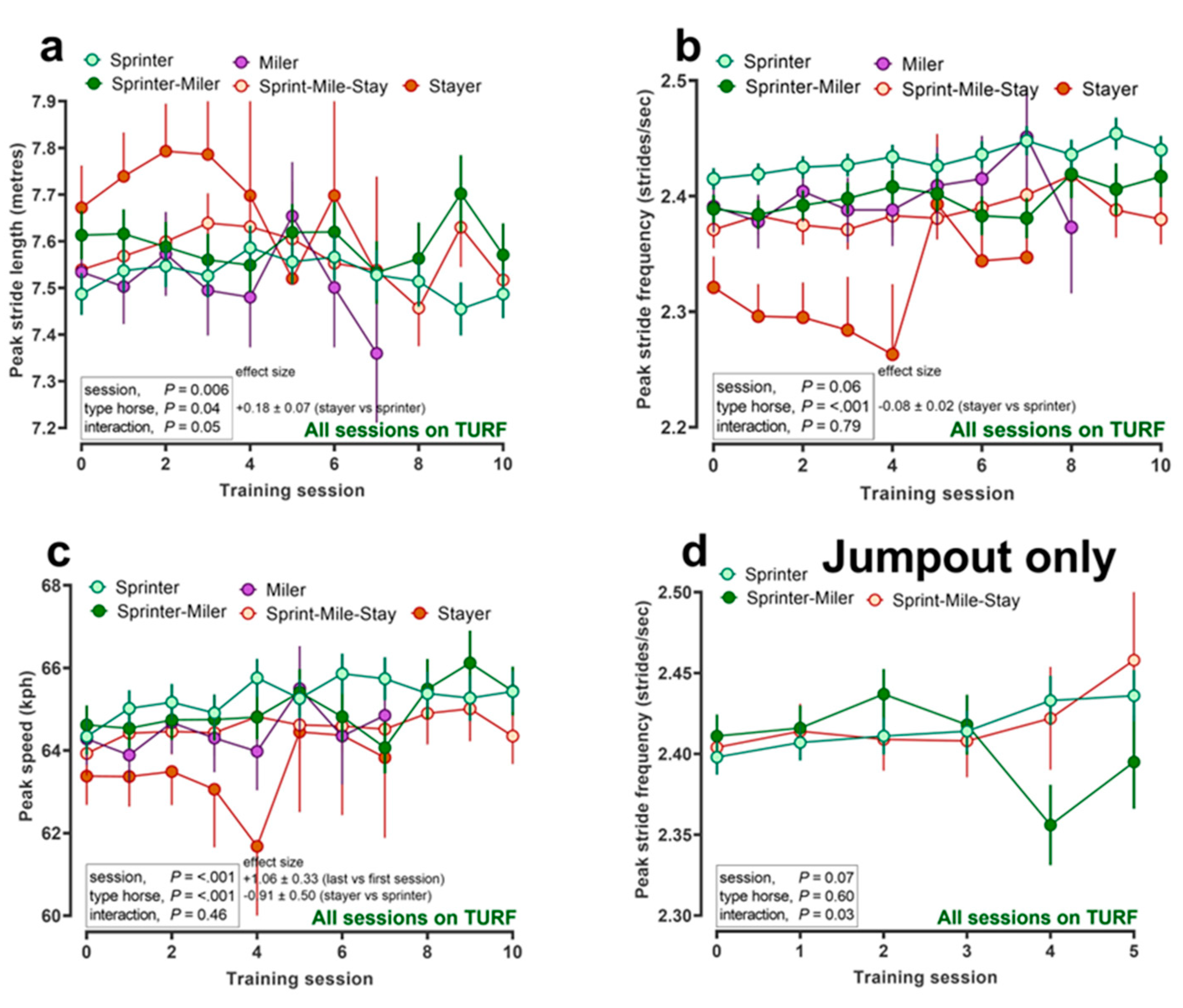
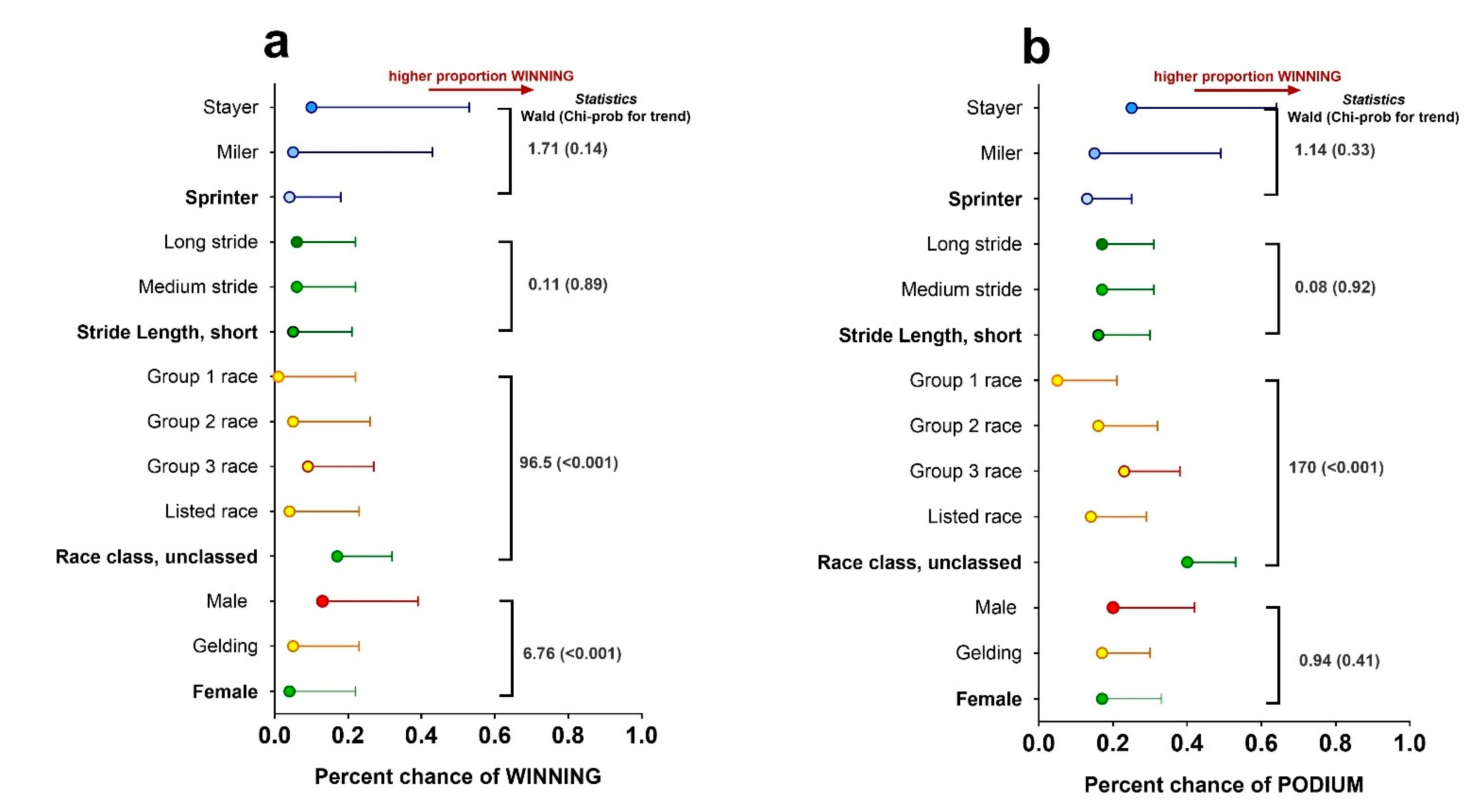

{kind=link}
{kind=link}
{kind=link}
| All (n = 3269) | Sprinter (n = 1671) | Sprinter-Miler (n = 874) | Miler (n = 167) | Sprinter-Miler-Stayer (n = 408) | Stayer (n = 149) | * p-Value | |
|---|---|---|---|---|---|---|---|
| Wins (%) | 508 (15.5) | 234 (14.0) | 134 (15.3) | 27 (16.2) | 74 (18.1) | 39 (26.2) | <0.001 |
| Top3 (%) | 1190 (36.4) | 593 (35.5) | 299 (34.2) | 60 (35.9) | 166 (40.7) | 72 (48.3) | <0.001 |
| Top5 (%) | 1668 (51.0) | 823 (49.3) | 422 (48.3) | 89 (53.3) | 239 (58.6) | 95 (63.8) | <0.001 |
| Soft Gallop (n = 635) | Medium Gallop (n = 579) | Hard Gallop (n = 585) | p-Value | |
|---|---|---|---|---|
| Best 0–200 m (s) | 11.6 ± 0.1 | 11.3 ± 0.1 | 10.8 ± 0.4 | <0.001 |
| Max speed (kph) | 62.5 ± 0.9 | 64.2 ± 0.8 | 66.8 ± 1.4 | <0.001 |
| Peak stride frequency (stride/s) | 2.37 ± 0.08 | 2.40 ± 0.00 | 2.45 ± 0.08 | <0.001 |
| Peak stride length (ms) | 7.35 ± 0.24 | 7.48 ± 0.24 | 7.63 ± 0.27 | <0.001 |
| Sprinter | Sprinter-Miler | Miler | Sprinter-Miler-Stayer | Stayer | * p-Value | |
|---|---|---|---|---|---|---|
| Training distance (m) | 4924 ± 1146 a | 4987 ± 1232 ab | 4993 ± 1192 ab | 5167 ± 1240 b | 5333 ± 1148 ab | 0.018 |
| ‘Work’ distance (m) | 1802 ± 380 a | 1912 ± 479 b | 1968 ± 479 b | 2124 ± 565 c | 2292 ± 650 c | <0.001 |
| Peak stride frequency (strides/sec) | ||||||
| Soft Gallop | 2.39 ± 0.07 a | 2.36 ± 0.07 b | 2.38 ± 0.08 bc | 2.35 ± 0.08 bc | 2.28 ± 0.06 c | Type horse, <0.001 Train intensity, <0.001 Interaction, 0.03 |
| Medium Gallop | 2.41 ± 0.07 | 2.39 ± 0.07 | 2.38 ± 0.08 | 2.37 ± 0.06 | 2.29 ± 0.08 | |
| Hard Gallop | 2.46 ± 0.08 * | 2.43 ± 0.08 * | 2.46 ± 0.05 * | 2.41 ± 0.06 * | 2.34 ± 0.05 * | |
| Peak stride length (meters) | ||||||
| Soft Gallop | 7.31 ± 0.23 a | 7.41 ± 0.23 ab | 7.37 ± 0.21 b | 7.41 ± 0.24 bc | 7.60 ± 0.20 c | Type horse, <0.001 Train intensity, <0.001 Interaction, 0.07 |
| Medium Gallop | 7.44 ± 0.24 | 7.50 ± 0.22 | 7.52 ± 0.30 | 7.56 ± 0.22 | 7.77 ± 0.24 | |
| Hard Gallop | 7.61 ± 0.28 * | 7.67 ± 0.27 * | 7.54 ± 0.20 * | 7.67 ± 0.24 * | 7.88 ± 0.08 * | |
| Sprinter | Sprinter-Miler | Miler | Sprinter-Miler-Stayer | Stayer | Heritability | * p-Value | |
|---|---|---|---|---|---|---|---|
| Peak stride frequency (stride/s) | 2.43 ± 0.01 a | 2.39 ± 0.01 b | 2.38 ± 0.01 b | 2.37 ± 0.01 b | 2.31 ± 0.02 c | 0.20 ± 0.15 | <0.001 |
| Peak stride length (meters) | 7.45 ± 0.02 a | 7.54 ± 0.03 ab | 7.49± 0.05 ab | 7.53 ± 0.04 b | 7.62 ± 0.06 ab | 0.18 ± 0.14 | 0.003 |
| Peak heart rate (beats/min) | 217 ± 0.6 a | 216 ± 0.8 a | 214 ± 1.6 a | 215 ± 1.2 a | 215 ± 1.8 a | 0.19 ± 0.15 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schrurs, C.; Blott, S.; Dubois, G.; Van Erck-Westergren, E.; Gardner, D.S. Locomotory Profiles in Thoroughbreds: Peak Stride Length and Frequency in Training and Association with Race Outcomes. Animals 2022, 12, 3269. https://doi.org/10.3390/ani12233269
Schrurs C, Blott S, Dubois G, Van Erck-Westergren E, Gardner DS. Locomotory Profiles in Thoroughbreds: Peak Stride Length and Frequency in Training and Association with Race Outcomes. Animals. 2022; 12(23):3269. https://doi.org/10.3390/ani12233269
Chicago/Turabian StyleSchrurs, Charlotte, Sarah Blott, Guillaume Dubois, Emmanuelle Van Erck-Westergren, and David S. Gardner. 2022. "Locomotory Profiles in Thoroughbreds: Peak Stride Length and Frequency in Training and Association with Race Outcomes" Animals 12, no. 23: 3269. https://doi.org/10.3390/ani12233269
APA StyleSchrurs, C., Blott, S., Dubois, G., Van Erck-Westergren, E., & Gardner, D. S. (2022). Locomotory Profiles in Thoroughbreds: Peak Stride Length and Frequency in Training and Association with Race Outcomes. Animals, 12(23), 3269. https://doi.org/10.3390/ani12233269