
The Effect of Frequency of Fresh Pasture Allocation on the Feeding Behaviour of High Production Dairy Cows


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Grazing Management
2.2. Animal Behaviour
2.3. Statistical Analysis
3. Results
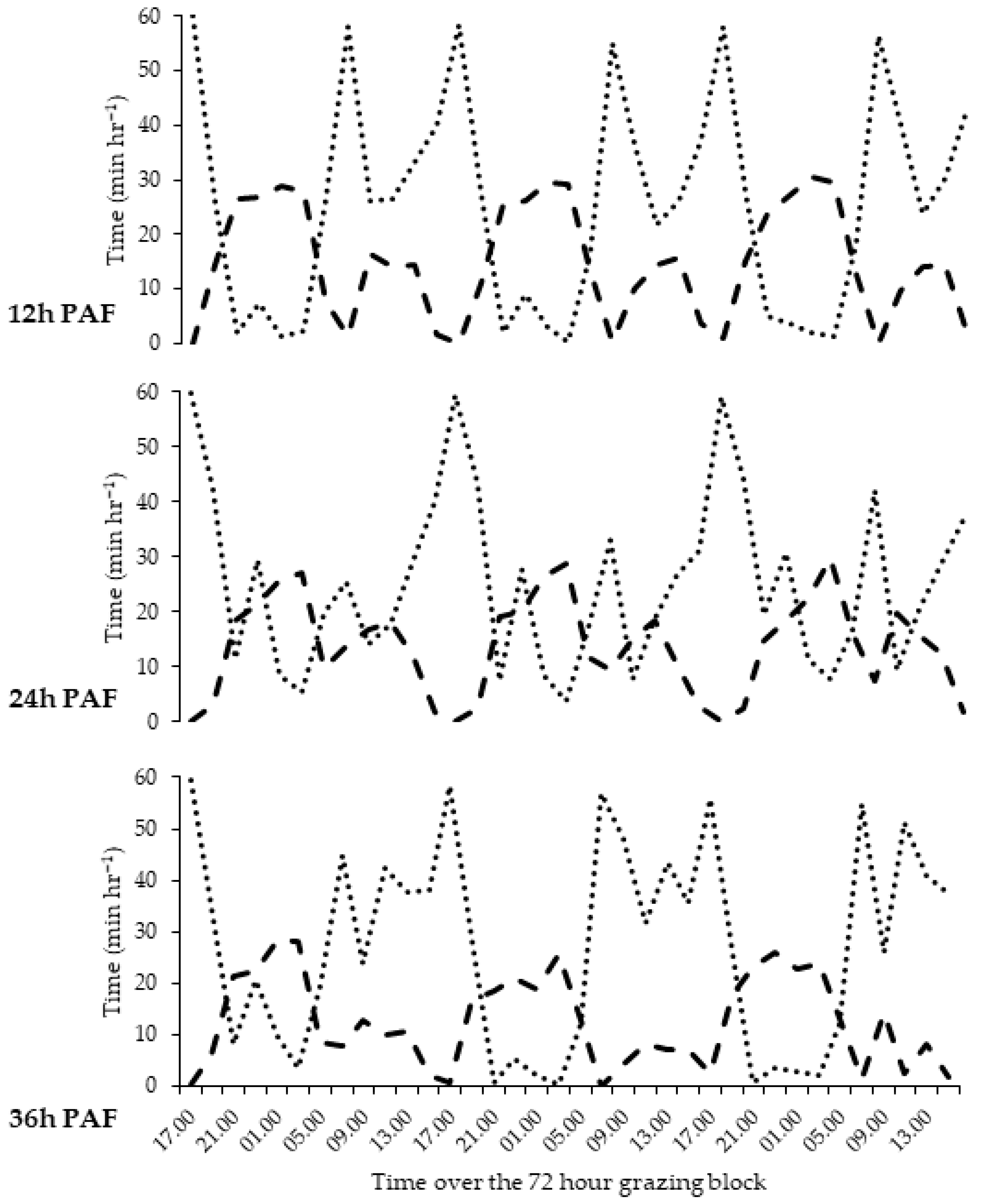
3.1. Feeding Behaviour
3.2. Interaction of Frequency of Pasture Allocation and Animal Parity Group
4. Discussion
4.1. Daily Feeding Patterns
4.2. Influence of PAF on Animal Feeding Behaviour
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kolver, E.; Muller, L. Performance and nutrient intake of high producing Holstein cows consuming pasture or a total mixed ration. J. Dairy Sci. 1998, 81, 1403–1411. [Google Scholar] [CrossRef]
- Rutter, S.; Orr, R.; Yarrow, N.; Champion, R. Dietary preference of dairy cows grazing ryegrass and white clover. J. Dairy Sci. 2004, 87, 1317–1324. [Google Scholar] [CrossRef] [Green Version]
- Prendiville, R.; Lewis, E.; Pierce, K.; Buckley, F. Comparative grazing behavior of lactating Holstein-Friesian, Jersey, and Jersey× Holstein-Friesian dairy cows and its association with intake capacity and production efficiency. J. Dairy Sci. 2010, 93, 764–774. [Google Scholar] [CrossRef] [Green Version]
- McEvoy, M.; Kennedy, E.; Murphy, J.; Boland, T.; Delaby, L.; O’donovan, M. The effect of herbage allowance and concentrate supplementation on milk production performance and dry matter intake of spring-calving dairy cows in early lactation. J. Dairy Sci. 2008, 91, 1258–1269. [Google Scholar] [CrossRef] [Green Version]
- Abrahamse, P.; Dijkstra, J.; Vlaeminck, B.; Tamminga, S. Frequent allocation of rotationally grazed dairy cows changes grazing behavior and improves productivity. J. Dairy Sci. 2008, 91, 2033–2045. [Google Scholar] [CrossRef]
- Vance, E.; Ferris, C.; Elliott, C.; Kilpatrick, D. A comparison of the feeding and grazing behaviour of primiparous Holstein-Friesian and Jersey× Holstein-Friesian dairy cows. Ir. J. Agric. Food Res. 2012, 51, 45–61. [Google Scholar]
- Susenbeth, A.; Mayer, R.; Koehler, B.; Neumann, O. Energy requirement for eating in cattle. J. Anim. Sci. 1998, 76, 2701–2705. [Google Scholar] [CrossRef] [PubMed]
- Osuji, P. The physiology of eating and the energy expenditure of the r uminant at pasture. Rangel. Ecol. Manag. J. Range Manag. Arch. 1974, 27, 437–443. [Google Scholar]
- Dohme-Meier, F.; Kaufmann, L.; Görs, S.; Junghans, P.; Metges, C.; Van Dorland, H.; Bruckmaier, R.; Münger, A. Comparison of energy expenditure, eating pattern and physical activity of grazing and zero-grazing dairy cows at different time points during lactation. Livest. Sci. 2014, 162, 86–96. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Braghieri, A.; Álvarez-Macías, A.; Serrapica, F.; Ramírez-Bribiesca, E.; Cruz-Monterrosa, R.; Masucci, F.; Mora-Medina, P.; Napolitano, F. The use of draught animals in rural labour. Animals 2021, 11, 2683. [Google Scholar] [CrossRef]
- Phillips, C.; Rind, M. The effects of social dominance on the production and behavior of grazing dairy cows offered forage supplements. J. Dairy Sci. 2002, 85, 51–59. [Google Scholar] [CrossRef]
- Hussein, A.N.; Al-Marashdeh, O.; Bryant, R.H.; Edwards, G. Relationship between Social Dominance and Milk Production of Dairy Cows Grazing Pasture; New Zealand Society of Animal Production: Wellington, New Zealand, 2016. [Google Scholar]
- DeVries, T.; Von Keyserlingk, M.; Weary, D. Effect of feeding space on the inter-cow distance, aggression, and feeding behavior of free-stall housed lactating dairy cows. J. Dairy Sci. 2004, 87, 1432–1438. [Google Scholar] [CrossRef]
- Chilibroste, P.; Mattiauda, D.; Bentancur, O.; Soca, P.; Meikle, A. Effect of herbage allowance on grazing behavior and productive performance of early lactation primiparous Holstein cows. Anim. Feed Sci. Technol. 2012, 173, 201–209. [Google Scholar] [CrossRef]
- Dale, A.; Laidlaw, A.; McGettrick, S.; Gordon, A.; Ferris, C. The effect of grazing intensity on the performance of high-yielding dairy cows. Grass Forage Sci. 2018, 73, 798–810. [Google Scholar] [CrossRef]
- Chilibroste, P.; Tamminga, S.; Boer, H. Effects of length of grazing session, rumen fill and starvation time before grazing on dry-matter intake, ingestive behaviour and dry-matter rumen pool sizes of grazing lactating dairy cows. Grass Forage Sci. 1997, 52, 249–257. [Google Scholar] [CrossRef]
- DeVries, T.; Von Keyserlingk, M.; Beauchemin, K. Frequency of feed delivery affects the behavior of lactating dairy cows. J. Dairy Sci. 2005, 88, 3553–3562. [Google Scholar] [CrossRef] [Green Version]
- Schütz, K.; Davison, D.; Matthews, L. Do different levels of moderate feed deprivation in dairy cows affect feeding motivation? Appl. Anim. Behav. Sci. 2006, 101, 253–263. [Google Scholar] [CrossRef]
- Verdon, M.; Rawnsley, R.; Raedts, P.; Freeman, M. The behaviour and productivity of mid-lactation dairy cows provided daily pasture allowance over 2 or 7 intensively grazed strips. Animals 2018, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Pollock, J.G.; Gordon, A.W.; Huson, K.M.; McConnell, D.A. The Effect of Frequency of Fresh Pasture Allocation on Pasture Utilisation and the Performance of High Yielding Dairy Cows. Animals 2020, 10, 2176. [Google Scholar] [CrossRef]
- Werner, J.; Leso, L.; Umstatter, C.; Niederhauser, J.; Kennedy, E.; Geoghegan, A.; Shalloo, L.; Schick, M.; O’Brien, B. Evaluation of the RumiWatchSystem for measuring grazing behaviour of cows. J. Neurosci. Methods 2018, 300, 138–146. [Google Scholar] [CrossRef]
- Werner, J.; Umstatter, C.; Kennedy, E.; Grant, J.; Leso, L.; Geoghegan, A.; Shalloo, L.; Schick, M.; O’Brien, B. Identification of possible cow grazing behaviour indicators for restricted grass availability in a pasture-based spring calving dairy system. Livest. Sci. 2019, 220, 74–82. [Google Scholar] [CrossRef]
- Rind, M.; Phillips, C. The effects of group size on the ingestive and social behaviour of grazing dairy cows. Anim. Sci. 1999, 68, 589–596. [Google Scholar] [CrossRef]
- Bargo, F.; Muller, L.D.; Delahoy, J.; Cassidy, T. Milk response to concentrate supplementation of high producing dairy cows grazing at two pasture allowances. J. Dairy Sci. 2002, 85, 1777–1792. [Google Scholar] [CrossRef] [Green Version]
- Stobbs, T. Automatic measurement of grazing time by dairy cows on tropical grass and legume pastures. Trop. Grassl. 1970, 4, 237–244. [Google Scholar]
- Stockdale, C.; King, K. Effect of stocking rate on the grazing behaviour and faecal output of lactating dairy cows. Grass Forage Sci. 1983, 38, 215–218. [Google Scholar] [CrossRef]
- Linnane, M.; Brereton, A.; Giller, P. Seasonal changes in circadian grazing patterns of Kerry cows (Bos taurus) in semi-feral conditions in Killarney National Park, Co. Kerry, Ireland. Appl. Anim. Behav. Sci. 2001, 71, 277–292. [Google Scholar] [CrossRef]
- Schirmann, K.; Chapinal, N.; Weary, D.M.; Heuwieser, W.; Von Keyserlingk, M.A. Rumination and its relationship to feeding and lying behavior in Holstein dairy cows. J. Dairy Sci. 2012, 95, 3212–3217. [Google Scholar] [CrossRef] [Green Version]
- Gregorini, P.; DelaRue, B.; McLeod, K.; Clark, C.; Glassey, C.; Jago, J. Rumination behavior of grazing dairy cows in response to restricted time at pasture. Livest. Sci. 2012, 146, 95–98. [Google Scholar] [CrossRef]
- Vibart, R.; Tavendale, M.; Otter, D.; Schwendel, B.; Lowe, K.; Gregorini, P.; Pacheco, D. Milk production and composition, nitrogen utilization, and grazing behavior of late-lactation dairy cows as affected by time of allocation of a fresh strip of pasture. J. Dairy Sci. 2017, 100, 5305–5318. [Google Scholar] [CrossRef] [Green Version]
- Dalley, D.; Roche, J.; Moate, P.; Grainger, C. More frequent allocation of herbage does not improve the milk production of dairy cows in early lactation. Aust. J. Exp. Agric. 2001, 41, 593–599. [Google Scholar] [CrossRef]
- DeVries, T.; Von Keyserlingk, M.; Beauchemin, K. Diurnal feeding pattern of lactating dairy cows. J. Dairy Sci. 2003, 86, 4079–4082. [Google Scholar] [CrossRef]
- Orr, R.; Penning, P.; Harvey, A.; Champion, R. Diurnal patterns of intake rate by sheep grazing monocultures of ryegrass or white clover. Appl. Anim. Behav. Sci. 1997, 52, 65–77. [Google Scholar] [CrossRef]
- Gregorini, P.; Eirin, M.; Refi, R.; Ursino, M.; Ansin, O.; Gunter, S. Timing of herbage allocation in strip grazing: Effects on grazing pattern and performance of beef heifers. J. Anim. Sci. 2006, 84, 1943–1950. [Google Scholar] [CrossRef]
- Orr, R.; Rutter, S.; Penning, P.; Rook, A. Matching grass supply to grazing patterns for dairy cows. Grass Forage Sci. 2001, 56, 352–361. [Google Scholar] [CrossRef]
- Gregorini, P.; Soder, K.; Sanderson, M.; Ziegler, G.R. Toughness, particle size and chemical composition of meadow fescue (Festuca pratensis Hud.) herbage as affected by time of day. Anim. Feed Sci. Technol. 2009, 151, 330–336. [Google Scholar] [CrossRef]
- Phillips, C.; Rind, M. The effects of frequency of feeding a total mixed ration on the production and behavior of dairy cows. J. Dairy Sci. 2001, 84, 1979–1987. [Google Scholar] [CrossRef]
- Phillips, C.; Leaver, J. The effect of forage supplementation on the behaviour of grazing dairy cows. Appl. Anim. Behav. Sci. 1986, 16, 233–247. [Google Scholar] [CrossRef]
- Niu, M.; Ying, Y.; Bartell, P.; Harvatine, K. The effects of feeding time on milk production, total-tract digestibility, and daily rhythms of feeding behavior and plasma metabolites and hormones in dairy cows. J. Dairy Sci. 2014, 97, 7764–7776. [Google Scholar] [CrossRef]
- Atkins, N.E.; Cianchi, C.; Rutter, S.; Williams, S.J.; Gauld, C.; Charlton, G.L.; Sinclair, L.A. Performance, milk fatty acid composition and behaviour of high-yielding Holstein dairy cows given a limited grazing period. Grass Forage Sci. 2020, 75, 181–191. [Google Scholar] [CrossRef]
- Bach, A.; Iglesias, C.; Devant, M.; Ràfols, N. Performance and feeding behavior of primiparous cows loose housed alone or together with multiparous cows. J. Dairy Sci. 2006, 89, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Wales, W.; Doyle, P.; Dellow, D. Dry matter intake and nutrient selection by lactating cows grazing irrigated pastures at different pasture allowances in summer and autumn. Aust. J. Exp. Agric. 1998, 38, 451–460. [Google Scholar] [CrossRef]
- Kondo, S.; Sekine, J.; Okubo, M.; Asahida, Y. The effect of group size and space allowance on the agonistic and spacing behavior of cattle. Appl. Anim. Behav. Sci. 1989, 24, 127–135. [Google Scholar] [CrossRef]
- Kennedy, E.; McEvoy, M.; Murphy, J.; O’donovan, M. Effect of restricted access time to pasture on dairy cow milk production, grazing behavior, and dry matter intake. J. Dairy Sci. 2009, 92, 168–176. [Google Scholar] [CrossRef]
- Olofsson, J. Competition for total mixed diets fed for ad libitum intake using one or four cows per feeding station. J. Dairy Sci. 1999, 82, 69–79. [Google Scholar] [CrossRef]
- Grant, R.; Colenbrander, V.; Albright, J. Effect of particle size of forage and rumen cannulation upon chewing activity and laterality in dairy cows. J. Dairy Sci. 1990, 73, 3158–3164. [Google Scholar] [CrossRef]
- Dado, R.; Allen, M. Variation in and relationships among feeding, chewing, and drinking variables for lactating dairy cows. J. Dairy Sci. 1994, 77, 132–144. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Behaviour Variable | Variable in RumiWatch System * | Variable Description |
|---|---|---|
| Grazing time (min cow−1 h−1) | EAT1TIME | Time spent eating (prehension bites and mastication chews in the downward position) |
| Ruminating time (min cow−1 h−1) | RUMINATETIME | Time spent ruminating per hour |
| Number of boli (n. cow−1 h−1) | BOLUS | Number of rumination boluses per hour |
| Chews per bolus (n. bolus−1) | CHEWSPERBOLUS | Number of chews per rumination bolus |
| Primiparous Animals | Multiparous Animals | Significance | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 12 h 1 | 24 h 2 | 36 h 3 | 12 h | 24 h | 36 h | SED 4 | Parity | Treatment | Interaction Effect | |
| Grazing time (min cow−1 h−1) | 24.3 c | 22.4 b,c | 22.3 b,c | 20.3 a | 21.4 a,b | 24.3 d | 0.61 | 0.004 | <0.001 | <0.001 |
| Rumination time (min cow−1 h−1) | 15.8 c | 13.2 a,b | 12.4 a | 14.2 b | 14.2 b | 12.8 a | 0.47 | 0.951 | <0.001 | <0.001 |
| Number of boli (no. cow−1 h−1) | 17.2 c | 14.3 a,b | 13.0 a | 15.7 b,c | 15.4 b | 13.9 a | 0.55 | 0.635 | <0.001 | 0.002 |
| Chews per bolus (no bolus−1) | 37.7 c | 32.0 a,b | 33.4 b | 31.5 a,b | 31.7 a,b | 29.9 a | 1.15 | <0.001 | 0.006 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pollock, J.G.; Gordon, A.W.; Huson, K.M.; McConnell, D.A. The Effect of Frequency of Fresh Pasture Allocation on the Feeding Behaviour of High Production Dairy Cows. Animals 2022, 12, 243. https://doi.org/10.3390/ani12030243
Pollock JG, Gordon AW, Huson KM, McConnell DA. The Effect of Frequency of Fresh Pasture Allocation on the Feeding Behaviour of High Production Dairy Cows. Animals. 2022; 12(3):243. https://doi.org/10.3390/ani12030243
Chicago/Turabian StylePollock, Jessica G., Alan W. Gordon, Kathryn M. Huson, and Deborah A. McConnell. 2022. "The Effect of Frequency of Fresh Pasture Allocation on the Feeding Behaviour of High Production Dairy Cows" Animals 12, no. 3: 243. https://doi.org/10.3390/ani12030243