
Analyses of Genetic Diversity in the Endangered “Berrenda” Spanish Cattle Breeds Using Pedigree Data

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data and Reference Population
2.2. Calculated Parameters
- (i)
- Generation interval: It is defined as the average age of parents (male or female) when its replacement is born [47]; it is calculated considering the four possible paths: sire-daughter, sire-son, dam-daughter and dam-calf;
- (ii)
- Proportion of present ancestors per generation: It was estimated in order to analyze the pedigree completeness level [48];
- (iii)
- Pedigree depth: It is calculated by considering the equivalent number of discreet generations equals to , where n is equal to the number of generations between the individual and its known ancestor; the individuals without known ancestors were assigned to the base generation [49]. The maximum number and complete generations of the pedigree have also been calculated. On the other side, the Inbreeding Coefficient (F) and the Average Relationship Coefficient (AR) have been calculated to analyze the inbreeding and the genetic weight of every individual over the whole population. F is defined as the likelihood that an individual carries two identical genes by descent. AR is defined as the average genetic weight of every individual over the whole population and it is equal to the average coancestry coefficient of every individual related to the other members of the population [42,50].
- (i)
- Effective number of herds that are rearing sires performing as grandparents, great-grandparents and great-great-grandparents: It was calculated according to Robertson [56] and it is defined as the inverse of the probability that two animals randomly chosen could belong to the same herd. Robertson’s statistics also allow identifying the herd they belong, depending on the exchange of breeding animals with other herds (nuclear, multiplier, commercial or isolated herds).
- (ii)
- Effective number of founders (fe) that equally contributes and determines the existing genetic diversity in the population: It was calculated according to , where q1 represents the genetic contribution to the population of founder [57].
- (iii)
- Effective number of ancestors (fa): It is defined as the number of ancestors, founders or not, that are needed to explain the population whole genetic variability [58].
- (iv)
- Marginal contributions of ancestors fa/fe reveals the possible bottlenecks that could have affected the population under study. It also considers the genetic variability provided by an animal that could not be explained by the contribution of its offspring. It was calculated according to Boichard [58].
- (v)
- Number of Founder Genome Equivalents (fg): It can be defined as the number of founders that would lead to a similar genetic diversity in the population under study if the founders were equally represented and with no loss of alleles [59]. The values of this parameter in each “berrenda” cattle breed were calculated as the inverse of twice the average coancestries in the reference populations [60].
- (vi)
- Genetic Conservation Index (GCI): It is calculated based on the genetic contributions of all the individuals considered as founders (pi) [4]. According to that, the highest values would be obtained by those individuals retaining a higher number of alleles from the existing ones in the founders’ population: .
- (vii)
- The amount of genetic diversity (GD) in the reference population, accounting for loss of diversity due to genetic drift and the unequal contribution of founder was calculated as follows [61]:
- (viii)
- The degree of genetic differentiation among herds, considered as the contribution level of the populations to the whole genetic variability, were estimated using Wright’s F statistics (1978) according to Caballero and Toro [62] and were adapted to the subpopulation sizes, as specified by Bartolomé et al. [63].
3. Results and Discussion
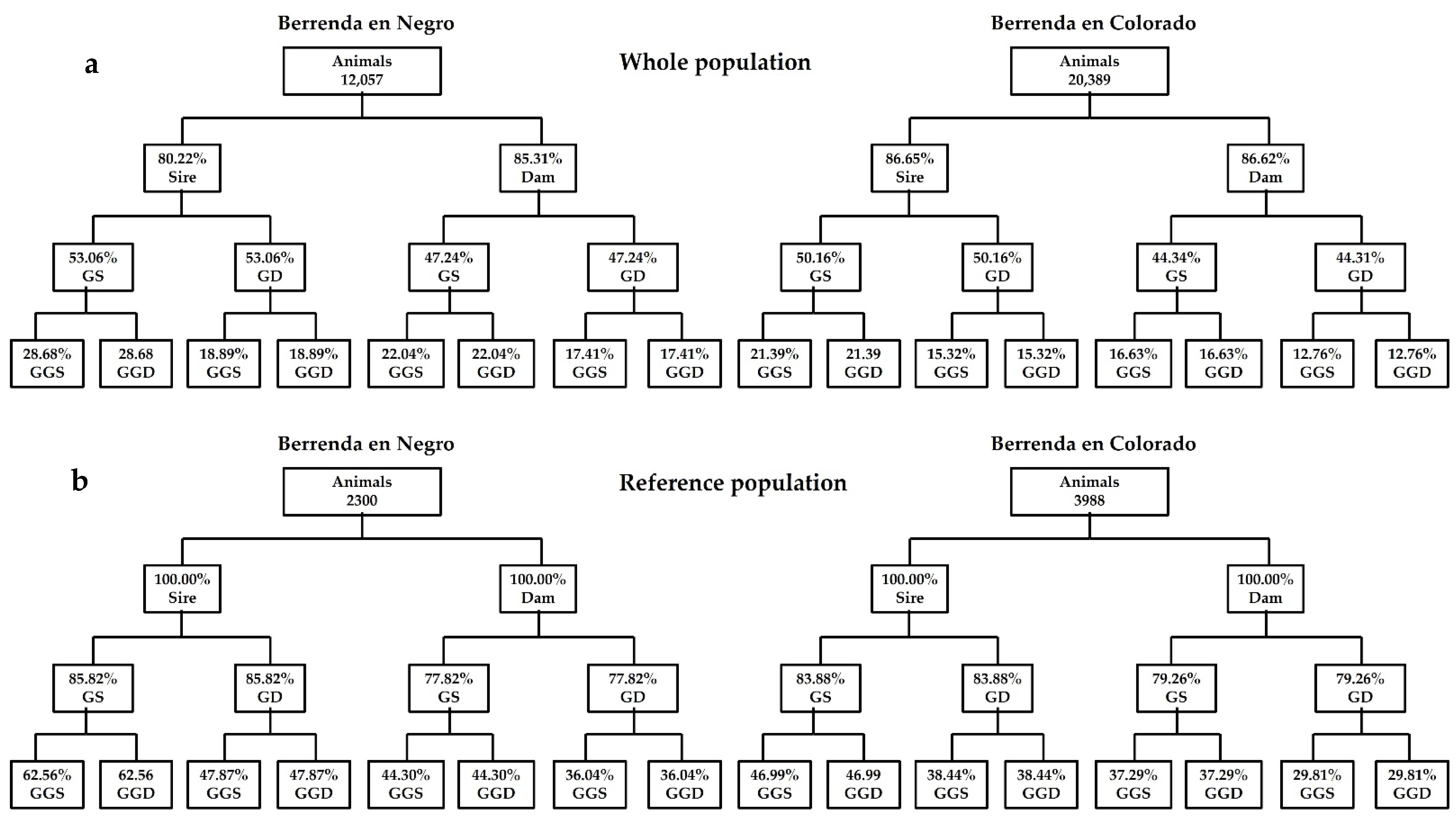
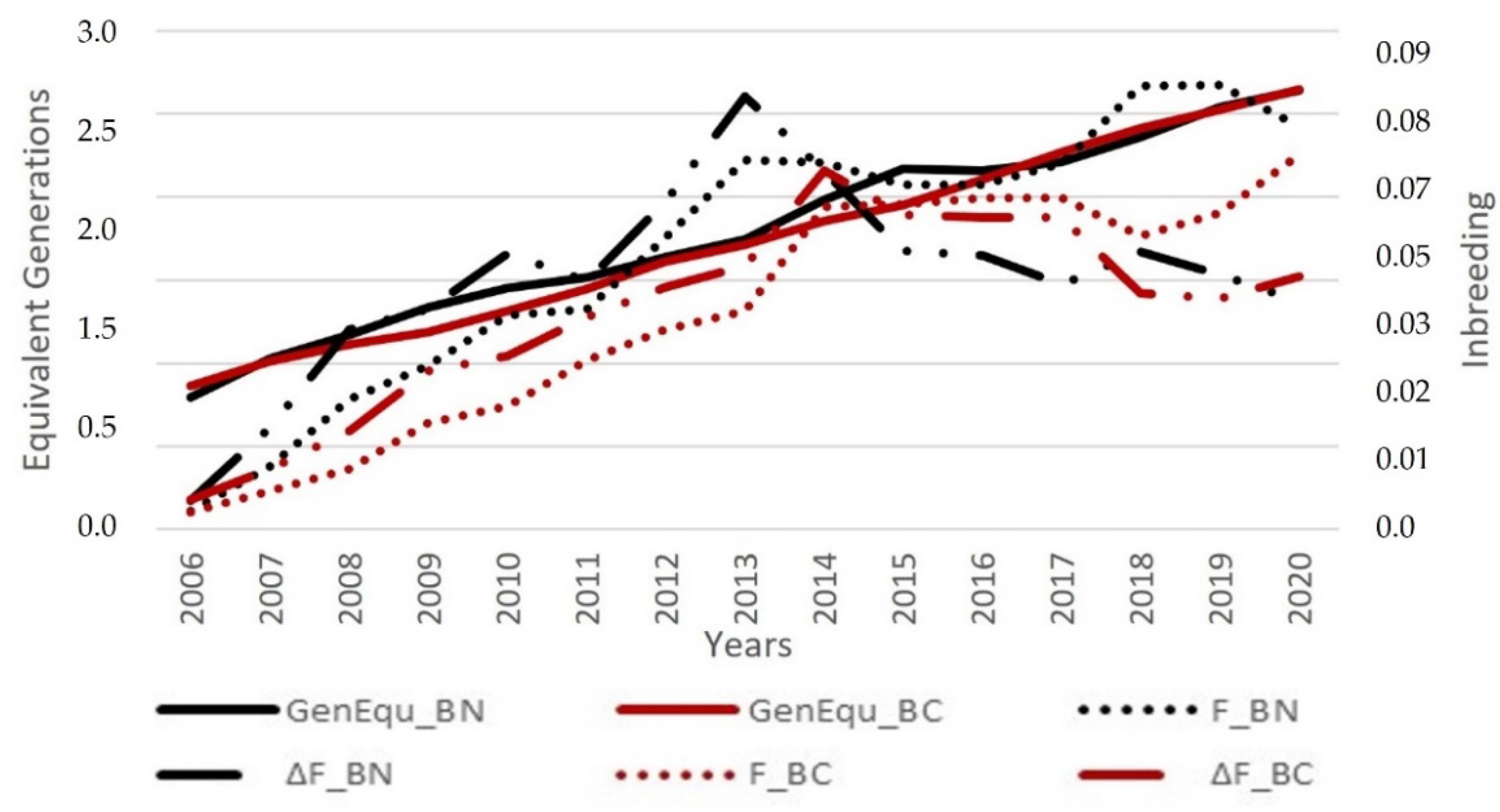
3.1. Population Structure and Pedigree Completeness Level
3.2. Studbooks’ Compression and Integrity of the Information
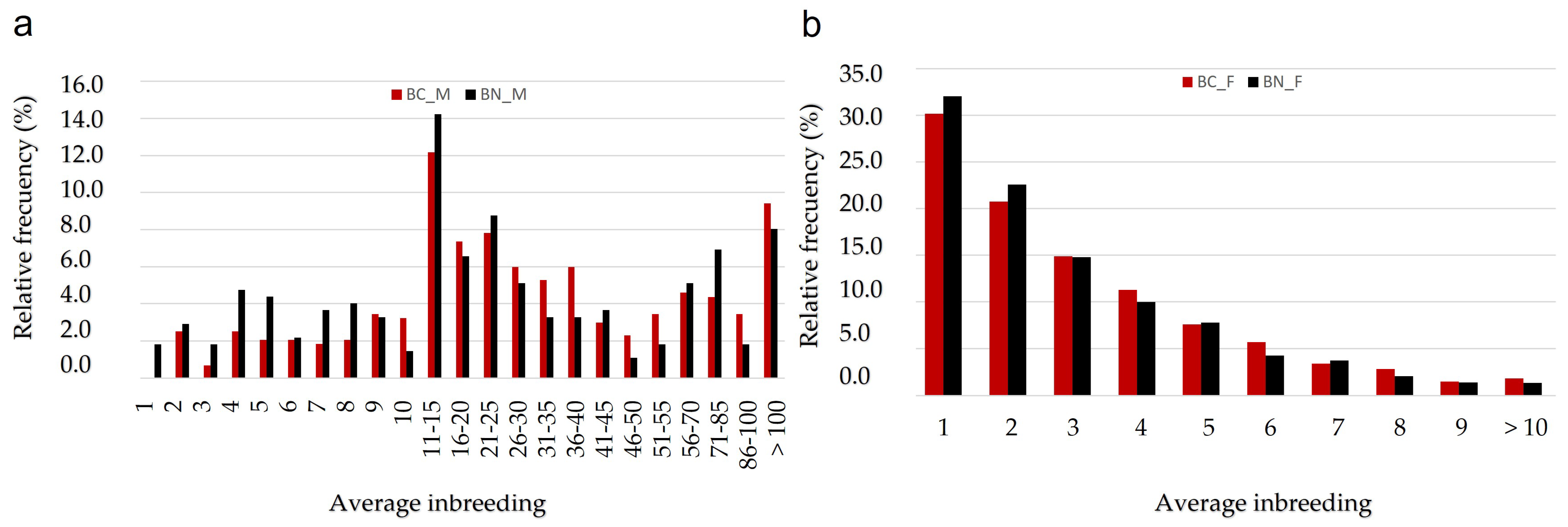
3.3. Inbreeding Analysis and Non-Random Mating Rate
3.4. Probability of Genes Origin and Ancestral Contributions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- BOE. Real Decreto 45/2019, de 8 de Febrero, por el Que se Establecen las Normas Zootécnicas Aplicables a los Animales Reproductores de Raza Pura, Porcinos Reproductores Híbridos y su Material Reproductivo, se Actualiza el Programa Nacional de Conservación, Mejora y Fomento de las Razas Ganaderas y se Modifican los Reales Decretos 558/2001, de 25 de Mayo; 1316/1992, de 30 de Octubre; 1438/1992, de 27 de Noviembre; y 1625/2011, de 14 de Noviembre. 2019. Available online: https://www.boe.es/eli/es/rd/2019/02/08/45/dof/spa/pdf (accessed on 20 November 2021).
- BOE. Orden de 30 de Julio de 1979 por la Que se Establece el Catálogo Oficial de Razas de Ganado de España. 1979. Available online: https://www.boe.es/boe/dias/1979/08/09/pdfs/A18725-18726.pdf (accessed on 30 December 2021).
- Spanish Breed Information System of the Ministry of Agriculture, Fisheries and Food (ARCA, by Its Initials in Spanish). 2021. Available online: https://www.mapa.gob.es/es/ganaderia/temas/zootecnia/razas-ganaderas/razas/catalogo-razas/bovino/berrenda-negro/iframe-ejemplo-arca.aspx (accessed on 20 November 2021).
- Alderson, L. The categorisation of types and breeds of cattle in Europe. Arch. Zootec. 1992, 41, 4. [Google Scholar]
- Rodero, E.; González, A.; Luque, A. Las razas bovinas andaluzas de protección especial: Berrenda en Colorado, Berrenda en Negro, Cárdena Andaluza, Negra Andaluza de las Campiñas, Pajuna y Marismeña. Patrim. Ganad. Andaluz. 2007, 2, 53–120. [Google Scholar]
- Pedrosa, S. El ADN Mitocondrial en el Análisis de la Domesticación Animal: Origen de las Razas Ovinas y Bovinas Ibéricas. Ph.D. Thesis, University of Leon, Leon, Spain, 2006. [Google Scholar]
- Azor Ortiz, P.J.; Valera Córdoba, M.; Gómez Ortiz, M.D.; Molina Alcalá, A. Assessment of Gene Flow Level Among Nucleus or Feral Losina Horse Using Melecular Coancestry Information. In Proceedings of the Annual Meeting of the European Association for Animal Production (57th EAAP), Antalya, Turkey, 17–20 September 2006; p. 110. [Google Scholar]
- BOE. Administrative Resolution from the Ministry of Agriculture, Orden AAA/1945/2013. 2013. Available online: https://www.boe.es/eli/es/o/2013/10/11/aaa1945/dof/spa/pdf (accessed on 20 November 2021).
- Web Page of “Agrupación de Asociaciones de Criadores de Ganado Vacuno de la Raza Berrenda en Negro y en Colorado”. Available online: www.anabe.net (accessed on 20 November 2021).
- Rodero, E.; Gonzalez, A.; Luque, M.; Herrera, M.; Gutierrez-Estrada, J.C. Classification of Spanish autochthonous bovine breeds. Morphometric study using classical and heuristic techniques. Livest. Sci. 2012, 143, 226–232. [Google Scholar] [CrossRef]
- Aparicio Sánchez, G. Zootecnia Especial. Etnologia Compendiada; Imprenta. Moderna: Cordoba, Spain, 1947. [Google Scholar]
- Mason, I.L. A World Dictionary of Livestock Breeds, Types and Varieties of Livestock; Technical Communication No. 8 of the Commonwealth Bureau of Animal Breeding and Genetics; Commonwealth Agricultural Bureaux: Wallingford, UK, 1951. [Google Scholar]
- Carolino, N.; Vitorino, A.; Carolino, I.; Pais, J.; Henriques, N.; Silveira, M.; Vicente, A. Genetic Diversity in the Portuguese Mertolenga Cattle Breed Assessed by Pedigree Analysis. Animals 2020, 10, 1990. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Cano, R.; González-Martínez, A.; Muñoz-Mejias, M.E.; Valera, P.; Rodero, E. Removal to undesirable MC1R gene alleles from ‘Berrenda en Negro’, an endangered Spanish cattle breed, to enhance breed conservation programs. Livest. Sci. 2022. accepted. [Google Scholar] [CrossRef]
- Rodero-Serrano, E.; Demyda-Peyrás, S.; González-Martinez, A.; Rodero-Franganillo, A.; Moreno-Millán, M. The chromosome translocation rob (1;29) in Andalusian endangered cattle breeds. Livest. Sci. 2013, 158, 32–39. [Google Scholar] [CrossRef]
- Ferraz-de-Oliveira, M.I.; Azeda, C.; Pinto-Correia, T. Management of montados and dehesas for high nature value: An interdisciplinary pathway. Agroforest. Syst. 2016, 90, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Sal, A.; Rodríguez, M.A.; De Miguel, J.M. Matter transfer and land use by cattle in a dehesa ecosystem of Central Spain. In Quercus ilex L. Ecosystems: Function, Dynamics and Management; Springer: Dordrecht, The Netherlands, 1992; pp. 345–354. [Google Scholar]
- Perea, R.; López-Sánchez, A.; Roig, S. The use of shrub cover to preserve Mediterranean oak dehesas: A comparison between sheep, cattle and wild ungulate management. Appl. Veg. Sci. 2016, 19, 244–253. [Google Scholar] [CrossRef]
- Escribano, A.J.; Gaspar, P.; Mesias, F.J.; Escribano, M.; Pulido, A.F. Competitiveness of ex-tensive beef cattle farms located in the dehesa ecosystem (SW Europe). In Proceedings of the Annual Meeting of the European Association for Animal Production (65th EAAP), Copenhagen, Denmark, 25–29 August 2014; pp. 25–29. [Google Scholar]
- Horrillo, A.; Gaspar, P.; Mesías, F.J.; Elghannam, A.; Escribano, M. Understanding the barriers and exploring the possibilities of the organic livestock sector in dehesa agroforestry systems: A multi-actor approach for effective diagnosis. Renew. Agric. Food Syst. 2020, 35, 663–677. [Google Scholar] [CrossRef]
- Cañon, J.; Garcia, D.; Delgado, J.V.; Dunner, S.; da Gama, L.T.; Landi, V.; Martin-Burriel, I.; Martinez, A.; Penedo, C.; Rodellar, C.; et al. Relative breed contributions to neutral genetic diversity of a comprehensive representation of Iberian native cattle. Animals 2011, 5, 1323–1334. [Google Scholar] [CrossRef] [Green Version]
- Felius, M.; Koolmees, P.A.; Theunissen, B.; Lenstra, J.A.; European Cattle Genetic Diversity Consortium. On the breeds of cattle—historic and current classifications. Diversity 2010, 3, 660–692. [Google Scholar] [CrossRef] [Green Version]
- Laloë, D.; Moazami-Goudarzi, K.; Lenstra, J.A.; Marsan, P.A.; Azor, P.; Baumung, R.; Bradley, D.G.; Bruford, M.W.; Cañón, J.; Dolf, G.; et al. European Cattle Genetic Diversity Consortium and Econogene Consortium. Spatial trends of genetic variation of domestic ruminants in Europe. Diversity 2010, 2, 932–945. [Google Scholar] [CrossRef] [Green Version]
- Menwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- FAO. Global Plan of Action for Animal Genetic Resources and the Interlaken Declaraction. Rome. 2007. Available online: https://www.fao.org/3/a1404e/a1404e.pdf (accessed on 20 November 2021).
- Makanjuola, B.O.; Miglior, F.; Abdalla, E.A.; Maltecca, C.; Schenkel, F.S.; Baes, C.F. Effect of genomic selection on rate of inbreeding and coancestry and effective population size of Holstein and Jersey cattle populations. J. Dairy Sci. 2020, 103, 5183–5199. [Google Scholar] [CrossRef]
- Leroy, G.; Mary-Huard, T.; Verrier, E.; Danvy, S.; Charvolin, E.; Danchin-Burge, C. Methods to estimate effective population size using pedigree data: Examples in dog, sheep, cattle and horse. Genet. Sel. Evol. 2013, 45, 1. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.C.; Gonçalves de Rezende, M.P.; Dadousis, C.; Biffani, S.; Negrini, R.; Souza Carneiro, P.L.; Bozzi, R. Population structure and genetic diversity of Italian beef breeds as a tool for planning conservation and selection strategies. Animals 2019, 9, 880. [Google Scholar] [CrossRef] [Green Version]
- Valera, M.; Molina, A.; Gutiérrez, J.P.; Gómez, J.; Goyache, F. Pedigree analysis in the Andalusian horse: Population structure, genetic variability and influence of the Carthusian strain. Livest. Prod. Sci. 2005, 95, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Zanella, R.; Lago, L.V.; Da Silva, A.N.; Pértille, F.; De Carvalho, N.S.; do Carmo Panetto, J.C.; Zanella, G.C.; Facioli, F.L.; Vinicius, M.; Da Silva, M.V.G. Genetic characterization of Indubrasil cattle breed population. Vet. Sci. 2018, 5, 98. [Google Scholar] [CrossRef] [Green Version]
- Cañas-Álvarez, J.J.; Gónzalez-Rodríguez, A.; Martín-Collado, D.; Avilés, C.; Altarriba, J.; Baro, J.A.; De la Fuente, L.F.; Díaz, C.; Molina, A.; Varona, L.; et al. Monitoring changes in the demographic and genealogical structure of the main Spanish local beef breeds. J. Anim. Sci. 2014, 92, 4364–4374. [Google Scholar] [CrossRef]
- Zhang, J.; Song, H.; Zhang, Q.; Ding, X. Assessment of relationships between pigs based on pedigree and genomic information. Animals 2020, 14, 697–705. [Google Scholar] [CrossRef]
- Cervantes, I.; Goyache, F.; Molina, A.; Valera, M.; Gutiérrez, J.P. Estimation of effective population size from the rate of coancestry in pedigreed populations. J. Anim. Breed. Genet. 2011, 128, 56–63. [Google Scholar] [CrossRef]
- Eusebi, P.G.; Martinez, A.; Cortes, O. Genomic tools for effective conservation of livestock breed diversity. Diversity 2020, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Gicquel, E.; Boettcher, P.; Besbes, B.; Furre, S.; Fernández, J.; Danchin-Burge, C.; Berger, B.; Baumung, R.; Feijóo, J.R.J.; Leroy, G. Impact of conservation measures on demography and genetic variability of livestock breeds. Animals 2020, 14, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Fioretti, M.; Negrini, R.; Biffani, S.; Quaglia, A.; Valentini, A.; Nardone, A. Demographic structure and population dynamics of Maremmana cattle local breed after 35 years of traditional selection. Livest. Sci. 2020, 232, 103903. [Google Scholar] [CrossRef]
- Jansson, M.; Laikre, L. Pedigree data indicate rapid inbreeding and loss of genetic diversity within populations of native, traditional dog breeds of conservation concern. PLoS ONE 2018, 13, e0202849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doublet, A.C.; Croiseau, P.; Fritz, S.; Michenet, A.; Hozé, C.; Danchin-Burge, C.; Laloë, D.; Restoux, G. The impact of genomic selection on genetic diversity and genetic gain in three French dairy cattle breeds. Genet. Sel. Evol. 2019, 51, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Surveying and Monitoring of Animal Genetic Resources. FAO Animal Production and Health Guidelines. No. 7. Rome. 2011. Available online: https://www.fao.org/3/ba0055e/ba0055e00.htm (accessed on 20 November 2021).
- González, R.; González, A.; Muñoz, E.; Valera, P.; Rodero, E. Evolución de la variabilidad genética de las dos razas bovinas berrendas autóctonas españolas (Periodo 2010–2018). In Proceedings of the XII Congreso de la Federación Iberoamericana de Razas Criollas y Autóctonas, Veracruz, Mexico, 9–12 October 2019. [Google Scholar]
- Cañón, J.; Gutiérrez, J.P.; Dunner, S.; Goyache, F.; Vallejo, M. Herdbook analyses of the Asturiana beef cattle breeds. Genet. Sel. Evol. 1994, 26, 65–75. [Google Scholar] [CrossRef]
- Gutiérrez, J.P.; Altarriba, J.; Díaz, C.; Quintanilla, R.; Cañón, J.; Piedrafita, J. Pedigree analysis of eight Spanish beef cattle breeds. Genet. Sel. Evol. 2003, 35, 43–63. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Justo, J.R.; Rivero, C.J.; Adán, S.; Rois, D.; Lama, J. Análisis de la información genealógica en las razas bovinas morenas gallegas. Arch. Zootec. 2007, 56, 607–615. [Google Scholar]
- Cortés, O.; Sevane, N.; Baro, J.A.; Cañón, J. Pedigree analysis of a highly fragmented population, the Lidia cattle breed. Livest. Sci. 2014, 167, 1–8. [Google Scholar] [CrossRef]
- Jordana, J.; Ferrando, A.; Marmi, J.; Avellanet, R.; Aranguren-Méndez, J.A.; Goyache, F. Molecular, genealogical and morphometric characterisation of the Pallaresa, a Pyrenean relic cattle breed: Insights for conservation. Livest. Sci. 2010, 132, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Benavente, M.F.; Santana, R.; Delgado, J.V. Gestión de la consanguinidad en la raza bovina palmera. AICA 2017, 10, 78–82. [Google Scholar]
- James, J.W. A note on selection differentials and generation length when generations overlap. Anim. Prod. 1977, 24, 109–112. [Google Scholar] [CrossRef]
- MacCluer, J.W.; Boyce, A.J.; Dyke, B.; Weitkamp, L.R.; Pfenning, D.W.; Parsons, C.J. Inbreeding and pedigree structure in Standardbred horses. J. Hered. 1983, 74, 394–399. [Google Scholar] [CrossRef]
- Maignel, L.; Boichard, D.; Verrier, E. Genetic variability of French dairy breeds estimated from pedigree infor-mation. Interbull. Bull. 1996, 14, 49–54. [Google Scholar]
- Goyache, F.; Gutiérrez, J.P.; Fernández, I.; Gómez, E.; Álvarez, I.; Díez, J.; Royo, L.J. Using pedigree information to monitor genetic variability of endangered populations: The Xalda sheep breed of Asturias as an example. J. Anim. Breed. Genet. 2003, 120, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.P.; Goyache, F. A note on ENDOG: A computer program for monitoring genetic variability of populations using pedigree information. J. Anim. Breed. Genet. 2005, 122, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.P.; Cervantes, I.; Molina, A.; Valera, M.; Goyache, F. Individual increase in inbreeding allows estimating effective sizes from pedigrees. Genet. Sel. Evol. 2008, 40, 359–378. [Google Scholar] [CrossRef]
- Gutiérrez, J.P.; Cervantes, I.; Goyache, F. Improving the estimation of realized effective population sizes in farm animals. J. Anim. Breed. Genet. 2009, 126, 327–332. [Google Scholar] [CrossRef]
- Cervantes, I.; Goyache, F.; Molina, A.; Valera, M.; Gutiérrez, J.P. Application of individual increase in inbreeding to estimate realized effective sizes from real pedigrees. J. Anim. Breed. Genet. 2008, 125, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Leroy, G.; Gicquel, E.; Boettcher, P.; Besbes, B.; Furre, S.; Fernandez, J.; Dnachin-Burge, C.; Alnahhas, N.; Baumung, R. Coancestry rate’s estimate of effective population size for genetic variability monitoring. Conserv. Genet. Resour. 2020, 12, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A. A numerical description of breed structure. J. Agric. Sci. 1953, 43, 334–336. [Google Scholar] [CrossRef]
- James, J.W. Computation of genetic contributions from pedigrees. Theor. Appl. Genet. 1972, 42, 272–273. [Google Scholar] [CrossRef] [PubMed]
- Boichard, D.; Maignel, L.; Verrier, E. The value of using probabilities of gene origin to measure genetic variability in a population. Gene. Sel. Evol. 1997, 29, 5–23. [Google Scholar] [CrossRef]
- Ballou, J.D.; Lacy, R.C. Identifying genetically important individuals for management of genetic variation in pedigreed populations. In Population Management for Survival and Recovery: Analytical Methods and Strategies in Small Population Management; Ballou, J.D., Gilpin, M., Foose, T.J., Eds.; Columbia University Press: New York, NY, USA, 1995; pp. 76–111. [Google Scholar]
- Caballero, A.; Toro, M.A. Interrelations between effective population size and other pedigree tools for the management of conserved populations. Genet. Res. 2000, 75, 331–343. [Google Scholar] [CrossRef]
- Lacy, R.C. Clarification of Genetic Terms and Their Use in the Management of Captive Populations; Wiley Subscription Services, Inc., A Wiley Company: New York, NY, USA, 1995; Volume 14, pp. 565–577. [Google Scholar]
- Caballero, A.; Toro, M.A. Analysis of genetic diversity for the management of conserved subdivided populations. Conserv. Genet. 2002, 3, 289–299. [Google Scholar] [CrossRef]
- Bartolomé, E.; Goyache, F.; Molina, A.; Cervantes, I.; Valera, M.; Gutiérrez, J.P. Pedigree estimation of the (sub) population contribution to the total gene diversity: The horse coat colour case. Animals 2010, 4, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, J.P.; Goyache, F.; Cervantes, I. User’s Guide of ENDOG v4.8: A Computer Program for Monitoring Genetic Variability of Populations Using Pedigree Information. 2010. Available online: https://www.google.es/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwikubqz9d3zAhWE0eAKHb9kAvQQFnoECAkQAQ&url=http%3A%2F%2Fwww.ucm.es%2Finfo%2Fprodanim%2Fhtml%2FJP_Web_archivos%2FEN_Us_G_.pdf&usg=AOvVaw24Q-pLC8wGYNFnKGulBqhp (accessed on 20 November 2021).
- Commission Regulation (EC) No 445/2002 of 26 February 2002 Laying Down Detailed Rules for the Application of Council Regulation (EC) No 1257/1999 on Support for Rural Development from the European Agricultural Guidance and Guarantee Fund (EAGGF). Available online: https://eur-lex.europa.eu/eli/reg/2002/445/oj (accessed on 10 January 2022).
- Regulation (EU) 2019/817 of the European Parliament and of the Council of 20 May 2019 on Establishing a Framework for Interoperability between EU Information Systems in the Field of Borders and Visa and Amending Regulations (EC) No 767/2008, (EU) 2016/399, (EU) 2017/2226, (EU) 2018/1240, (EU) 2018/1726 and (EU) 2018/1861 of the European Parliament and of the Council and Council Decisions 2004/512/EC and 2008/633/JHA. Available online: https://eur-lex.europa.eu/eli/reg/2019/817/oj (accessed on 10 January 2022).
- Santana, M.L., Jr.; Pereira, R.J.; Bignardi, A.B.; Ayres, D.R.; Menezes, G.D.O.; Silva, L.O.C.; Leroy, G.; Machado, C.H.C.; Josahkian, L.A.; Albuquerque, L.G. Structure and genetic diversity of Brazilian Zebu cattle breeds assessed by pedigree analysis. Livest. Sci. 2016, 187, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Santana, M.L., Jr.; Pereira, R.J.; Bignardi, A.B.; El Faro, L.; Tonhati, H.; Albuquerque, L.G. History, structure, and genetic diversity of Brazilian Gir cattle. Livest. Sci. 2014, 163, 26–33. [Google Scholar] [CrossRef]
- Rezende, M.P.G.; Conde, E.A.S.L.; Borges, A.C.; Carneiro, P.L.S.; Martins Filho, R.; Malhado, C.H.M. Population structure of the Nellore herd reared in the Brazilian Northeast semi-arid. Cienc. Anim. Bras. 2017, 18, e-38048. [Google Scholar] [CrossRef] [Green Version]
- Malhado, C.H.M.; Malhado, A.C.M.; Martins Filho, R.; Carneiro, P.L.S.; Pala, A.; Carrillo, J.A. Age at first calving of Nellore cattle in the semi-arid region of northeastern Brazil using linear, threshold, censored and penalty models. Livest. Sci. 2013, 154, 28–33. [Google Scholar] [CrossRef]
- Van Grevenhof, E.M.; Van Arendonk, J.A.; Bijma, P. Response to genomic selection: The Bulmer effect and the potential of genomic selection when the number of phenotypic records is limiting. Genet. Sel. Evol. 2012, 44, 26. [Google Scholar] [CrossRef] [Green Version]
- González de Rezende, M.P.; Malhado, C.H.M.; Biffani, S.; Souza Carneiro, P.L.; Bozzi, R. Genetic diversity derived from pedigree information and estimation of genetic parameters for reproductive traits of Limousine and Charolais cattle raised in Italy. Ital. J. Anim. Sci. 2020, 19, 762–771. [Google Scholar] [CrossRef]
- Kamiti, D.; Ilatsia, E.; Bett, R.; Kahi, A. Population structure and demographic trends of the registered Sahiwal cattle in Kenya. Trop. Anim. Health Prod. 2016, 48, 1029–1036. [Google Scholar] [CrossRef]
- Illa, S.K.; Gollamoori, G.; Nath, S. Evaluation of selection program by assessing the genetic diversity and inbreeding effects on Nellore sheep growth through pedigree analysis. Asian-Australas. J. Anim. Sci. 2020, 33, 1369. [Google Scholar] [CrossRef] [Green Version]
- Cassell, B.G.; Adamec, V.; Pearson, R.E. Effect of incomplete pedigrees on estimates of inbreeding and inbreeding depression for days to first service and summit milk yield in Holsteins and Jerseys. J. Dairy Sci. 2003, 86, 2967–2976. [Google Scholar] [CrossRef]
- Bouquet, A.; Venot, E.; Laloë, D.; Forabosco, F.; Fogh, A.; Pabiou, T.; Moore, K.; Eriksson, J.-Å.; Renand, G.; Phocas, F. Genetic structure of the European Charolais and Limousin cattle metapopulations using pedigree analyses. J. Anim. Sci. 2011, 89, 1719–1730. [Google Scholar] [CrossRef]
- Regulation (EU) 2016/1012 of the European Parliament and of the Council of 8 June 2016 on Zootechnical and Genealogical Conditions for the Breeding, Trade in and Entry into the Union of Purebred Breeding Animals, Hybrid Breeding Pigs and the Germinal Products thereof and Amending Regulation (EU) No 652/2014, Council Directives 89/608/EEC and 90/425/EEC and Repealing Certain Acts in the Area of Animal Breeding (‘Animal Breeding Regulation’). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32016R1012&from=EN (accessed on 20 November 2021).
- Schäler, J.; Krüger, B.; Thaller, G.; Hinrichs, D. Comparison of ancestral, partial, and genomic inbreeding in a local pig breed to achieve genetic diversity. Conserv. Genet. Resour. 2020, 12, 77–86. [Google Scholar] [CrossRef]
- Burrow, H.M. The effects of inbreeding in beef cattle. Anim. Breed. Abstr. 1993, 61, 737–751. [Google Scholar]
- Wainwright, W.; Vosough Ahmadi, B.; Mcvittie, A.; Simm, G.; Moran, D. Prioritising support for cost effective rare breed conservation using multi-criteria decision analysis. Front. Ecol. Evol. 2019, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Vozzi, P.A.; Marcondes, C.R.; Magnabosco, C.U.; Bezerra, L.A.F.; Lôbo, R.B. Structure and genetic variability in Nellore (Bos indicus) cattle by pedigree analysis. Genet. Mol. Biol. 2006, 29, 482–485. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Ballou, S.E.J.D.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar] [CrossRef]
- Caballero, A.; Santiago, E.; Toro, M.A. Systems of mating to reduce inbreeding in selected populations. Anim. Sci. 1996, 62, 431–442. [Google Scholar] [CrossRef]
- Fernández, J.; Meuwissen, T.H.E.; Toro, M.A.; Mäki-Tanila, A. Management of genetic diversity in small farm animal populations. Animal 2011, 5, 1684–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenbroek, K. Utilisation and Conservation of Farm Animal Genetic Resources; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; p. 232. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Population | Reference Population | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Generation Interval | Average Age of Breeding Animals | Generation Interval | Average Age of Breeding Animals | |||||||||
| N | Mean ± SE | SD | N | Mean ± SE | SD | N | Mean ± SE | SD | N | Mean ± SE | SD | |
| Sire-Son | ||||||||||||
| BN | 187 | 4.92 ± 0.17 | 2.30 | 3902 | 5.54 ± 0.04 | 2.49 | 36 | 5.47 ± 0.45 | 2.68 | 514 | 5.92 ± 0.12 | 2.81 |
| BC | 264 | 5.58 ± 0.15 | 2.50 | 7810 | 6.04 ± 0.03 | 2.68 | 40 | 6.39 ± 0.47 | 2.99 | 1020 | 6.21 ± 0.09 | 2.73 |
| Sire-Daughter | ||||||||||||
| BN | 1912 | 5.40 ± 0.05 | 2.40 | 5770 | 5.70 ± 0.03 | 2.64 | 599 | 5.79 ± 0.49 | 2.96 | 1786 | 6.02 ± 0.13 | 2.99 |
| BC | 3083 | 5.72 ± 0.04 | 2.45 | 9857 | 6.02 ± 0.03 | 2.71 | 914 | 6.43 ± 0.46 | 2.91 | 2968 | 6.36 ± 0.09 | 2.92 |
| Dam-Son | ||||||||||||
| BN | 187 | 6.70 ± 0.26 | 3.60 | 4203 | 7.55 ± 0.06 | 4.03 | 36 | 6.67 ± 0.73 | 4.38 | 514 | 7.81 ± 0.18 | 4.18 |
| BC | 264 | 7.35 ± 0.23 | 3.72 | 7810 | 7.55 ± 0.04 | 3.87 | 40 | 7.68 ± 0.60 | 3.78 | 1020 | 7.89 ± 0.12 | 4.02 |
| Sire-Daughter | ||||||||||||
| BN | 1912 | 7.09 ± 0.09 | 3.96 | 6083 | 7.53 ± 0.05 | 4.09 | 599 | 7.27 ± 0.71 | 4.24 | 1786 | 7.72 ± 0.19 | 4.31 |
| BC | 3082 | 7.33 ± 0.07 | 3.91 | 9852 | 7.60 ± 0.04 | 3.98 | 914 | 7.40 ± 0.60 | 3.77 | 2968 | 7.77 ± 0.13 | 4.01 |
| Total | ||||||||||||
| BN | 4198 | 6.20 ± 0.05 | 3.36 | 19,958 | 6.62 ± 0.03 | 3.56 | 1270 | 6.50 ± 0.10 | 3.73 | 4600 | 6.87 ± 0.06 | 3.78 |
| BC | 6693 | 6.52 ± 0.04 | 3.36 | 35,329 | 6.80 ± 0.02 | 3.46 | 1908 | 6.92 ± 0.08 | 3.40 | 7976 | 7.06 ± 0.04 | 3.57 |
| Berrenda en Negro | Berrenda en Colorado | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Generation | n | F (%) | Inb (%) | FInb (%) | AR (%) | Ne | n | F (%) | Inb (%) | FInb (%) | AR (%) | Ne |
| 0 | 2386 | 0 | 0.18 | 4235 | 0 | 0.02 | ||||||
| 1 | 5250 | 2.25 | 8.59 | 26.18 | 0.80 | 22.2 | 10,517 | 1.98 | 8.00 | 24.79 | 0.33 | 25.2 |
| 2 | 3624 | 9.20 | 60.43 | 15.22 | 1.38 | 7.0 | 6426 | 6.72 | 47.82 | 14.06 | 0.53 | 10.3 |
| 3 | 774 | 11.88 | 84.24 | 14.11 | 2.34 | 16.9 | 718 | 8.57 | 63.79 | 13.43 | 0.64 | 25.2 |
| 4 | 23 | 12.10 | 100 | 12.10 | 2.45 | 202.9 | 1 | 19.53 | 100.00 | 19.53 | 0.58 | 4.1 |
| Berrenda en Negro | Berrenda en Colorado | |||||
|---|---|---|---|---|---|---|
| Maximum a | Complete b | Equivalent c | Maximum a | Complete b | Equivalent c | |
| Average number of generations | 2.15 | 1.24 | 1.62 | 1.97 | 1.25 | 1.54 |
| Inbreeding rate (%) per generation (ΔF) | 2.47 | 4.59 | 4.02 | 1.75 | 3.53 | 3.01 |
| Effective Population Size (Ne) | 20.22 | 10.90 | 12.44 | 28.62 | 14.18 | 16.62 |
| N | ΔF | F10 | F50 | Nei | Nec | Equivalent Subpopulations (Nec/Nei) | |
|---|---|---|---|---|---|---|---|
| BN | 1716 | 3.01 | 6.15 | 33.7 | 9.93 ± 3.76 | 92.28 ± 4.21 | 9.29 ± 3.54 |
| BC | 2968 | 4.02 | 4.34 | 18.78 | 11.58 ± 4.81 | 169.92 ± 5.74 | 14.67 ± 6.11 |
| BN | BC | |||
|---|---|---|---|---|
| Parameters 1 | Total | Reference Population | Total | Reference Population |
| Total number of animals | 12,057 | 2300 | 20,389 | 3988 |
| Total number of animals with both known parents | 9671 | 17,662 | ||
| Total number of founders | 2386 | 732 | 2727 | 1240 |
| Number of ancestors | 1657 | 726 | 2741 | 1242 |
| Average inbreeding coefficient | 4.53 | 7.00 | 3.44 | 5.70 |
| Average relationship | 0.95 | 0.40 | ||
| Effective number of founders | 88 | 58 | 238 | 140 |
| Effective number of ancestors | 87 | 57 | 234 | 133 |
| Number of Founder Herds | 75 | 135 | ||
| Effective number of founder herds | 23.9 | 60.9 | ||
| Equivalent number of founder genomes | 104.77 | 251.12 | ||
| fa/fe | 1.01 | 1.01 | 1.01 | 1.05 |
| fg/fe | 1.19 | 1.05 | ||
| fg/fa | 1.20 | 1.09 | ||
| Genetic Diversity | 0.9952 | 0.9983 | ||
| GD Loss caused by bottle necks or genetic drift since founders level | −0.0013 | −0.0003 | ||
| Number of ancestors explaining 50% of genetic variability | 50 | 36 | 101 | 57 |
| Genetic conservation Index | 2.80 | 2.82 | ||
| Berrenda en Negro | Berrenda en Colorado | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sex | Herd of Origin | Year of Birth | Genetic Contribution (%) | No. of Descendants | Sex | Herd of Origin | Year of Birth | Genetic Contribution (%) | No. of Descendants | |
| Whole Population | ||||||||||
| 1 | M | 52 | 1999 | 0.06 | 264 | M | 84 | 2003 | 0.02 | 315 |
| 2 | M | 84 | 2003 | 0.11 | 276 | H | 162 | 2005 | 0.04 | 6 |
| 3 | M | 35 | 2001 | 0.14 | 54 | M | 2 | 2003 | 0.05 | 219 |
| 4 | M | 19 | 2003 | 0.17 | 287 | M | 72 | 2003 | 0.07 | 198 |
| 5 | M | 151 | 2003 | 0.19 | 90 | M | 8 | 2001 | 0.08 | 175 |
| Reference Population | ||||||||||
| 1 | M | 84 | 2003 | 0.07 | 276 | M | 84 | 2003 | 0.04 | 315 |
| 2 | M | 52 | 1999 | 0.13 | 264 | H | 162 | 2005 | 0.07 | 6 |
| 3 | M | 35 | 2001 | 0.17 | 54 | M | 72 | 2003 | 0.08 | 198 |
| 4 | M | 151 | 2003 | 0.21 | 90 | M | 112 | 2006 | 0.1 | 265 |
| 5 | H | 52 | 2003 | 0.24 | 5 | M | 48 | 2006 | 0.12 | 98 |
| Generation | Berrenda en Negro | Berrenda en Colorado | ||
|---|---|---|---|---|
| Current | Effective | Current | Effective | |
| 1 | 99 | 36.97 | 166 | 66.40 |
| 2 | 55 | 16.91 | 88 | 36.47 |
| 3 | 27 | 8.18 | 42 | 19.06 |
| 4 | 14 | 6.24 | 11 | 6.35 |
| 5 | 3 | 2.81 | 2 | 1.19 |
| Herd Types | Berenda en Negro | Berrenda en Colorado | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| UPB | UOB | SB | NH | PPB% | UPB | UOB | SB | NH | PPB% | |
| Nucleus herd | No | Yes | Yes | 0 | 0 | No | Yes | Yes | 0 | 0 |
| Multiplier herd type A | Yes | Yes | Yes | 62 | 35.37 | Yes | Yes | Yes | 116 | 35.63 |
| Multiplier herd type B | Yes | No | Yes | 5 | 100 | Yes | No | Yes | 4 | 100 |
| Commercial herd type A | Yes | Yes | No | 30 | 32.75 | Tes | Yes | No | 44 | 29.13 |
| Commercial herd type B | Yes | No | No | 22 | 100 | Yes | No | No | 46 | 100 |
| Isolated herd | No | Yes | No | 0 | 0 | No | Yes | No | 0 | 0.00 |
| Total | 119 | 210 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Cano, R.; González-Martínez, A.; Muñoz-Mejías, M.E.; Valera, P.; Rodero, E. Analyses of Genetic Diversity in the Endangered “Berrenda” Spanish Cattle Breeds Using Pedigree Data. Animals 2022, 12, 249. https://doi.org/10.3390/ani12030249
González-Cano R, González-Martínez A, Muñoz-Mejías ME, Valera P, Rodero E. Analyses of Genetic Diversity in the Endangered “Berrenda” Spanish Cattle Breeds Using Pedigree Data. Animals. 2022; 12(3):249. https://doi.org/10.3390/ani12030249
Chicago/Turabian StyleGonzález-Cano, Rafael, Ana González-Martínez, María Eva Muñoz-Mejías, Pablo Valera, and Evangelina Rodero. 2022. "Analyses of Genetic Diversity in the Endangered “Berrenda” Spanish Cattle Breeds Using Pedigree Data" Animals 12, no. 3: 249. https://doi.org/10.3390/ani12030249