
Identification of Conservation Priority Areas and a Protection Network for the Siberian Musk Deer (Moschus moschiferus L.) in Northeast China

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
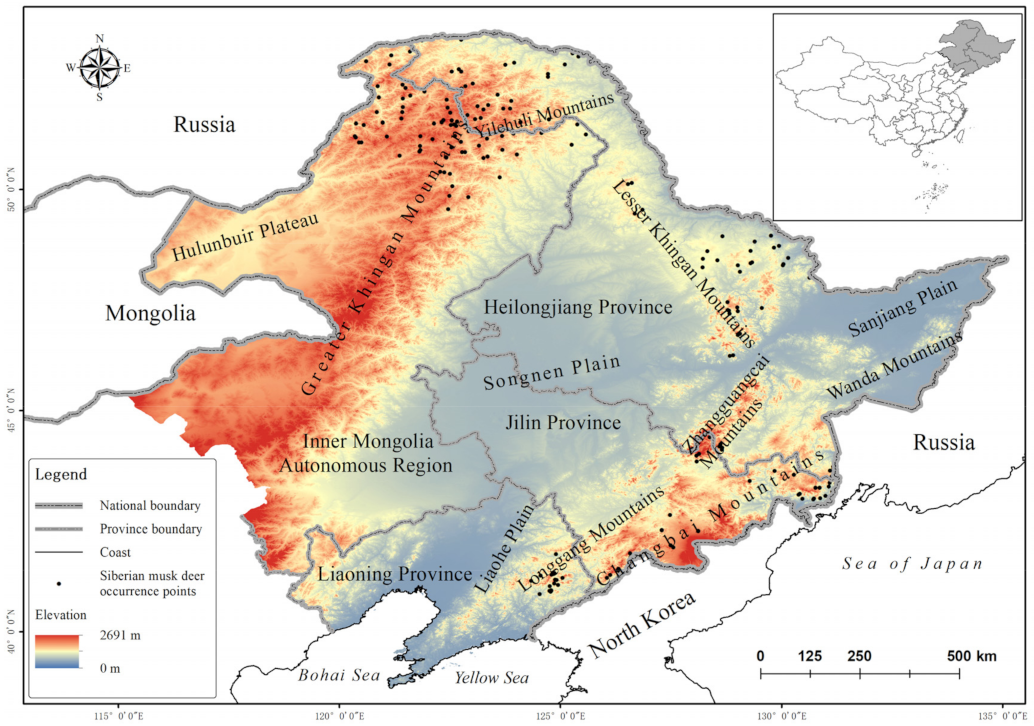
2.1. Study Area
2.2. Data Collection
2.3. Species Distribution Modeling
2.4. Habitat Quality Ranking and Identification of Conservation Priority Areas
2.5. Landscape Analysis and Potential Connectivity Corridor Construction
3. Results
3.1. Model Performance, Important Variables and Human Influence on M. moschiferus
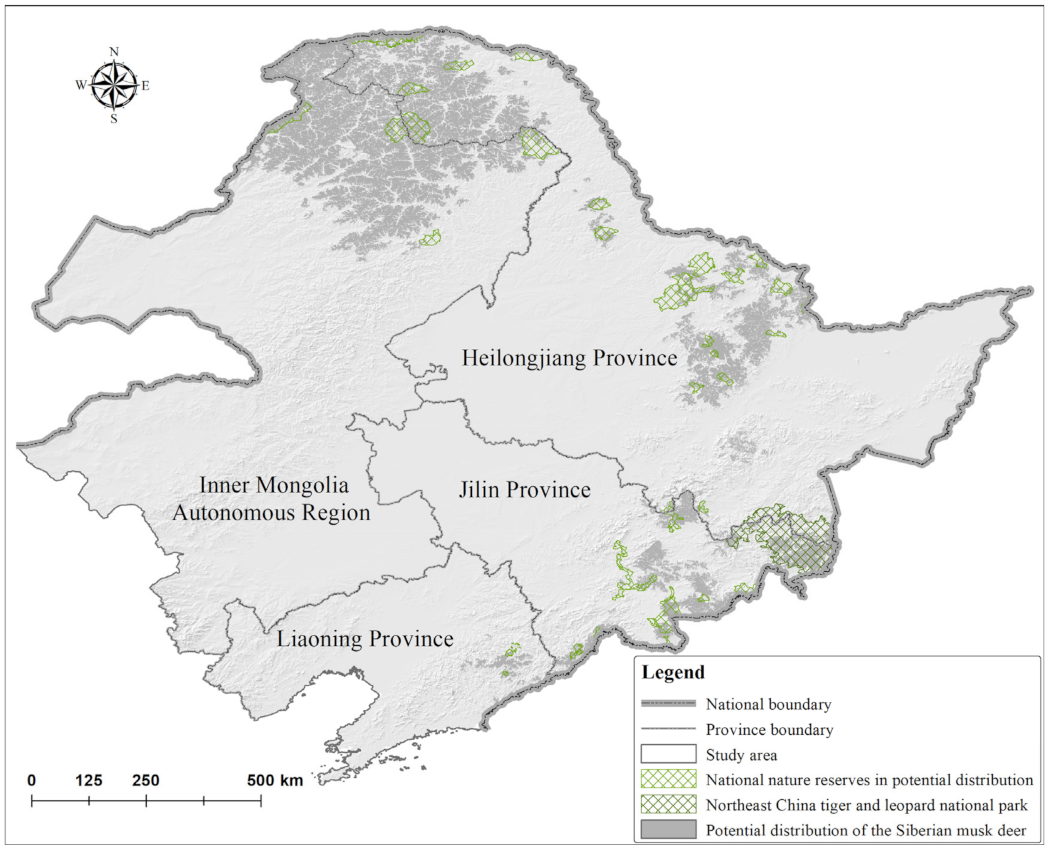
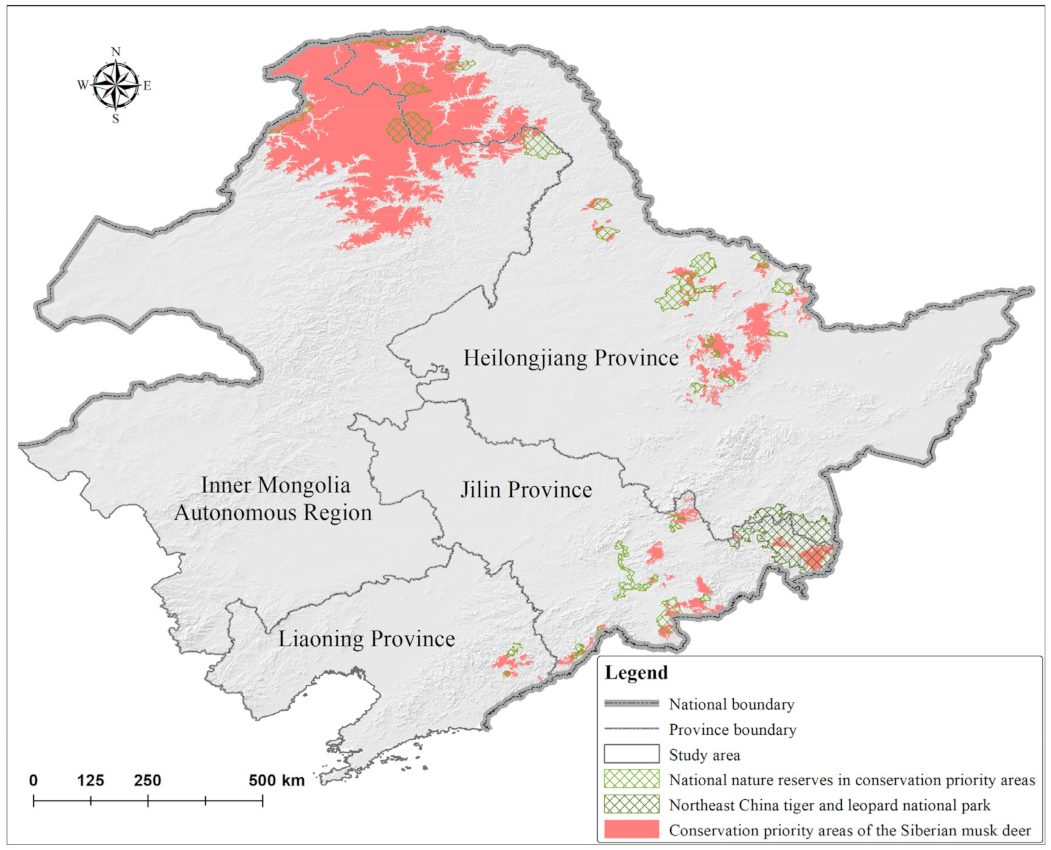
3.2. Potential Distribution, Conservation Priority Areas, and Conservation Status of M. moschiferus
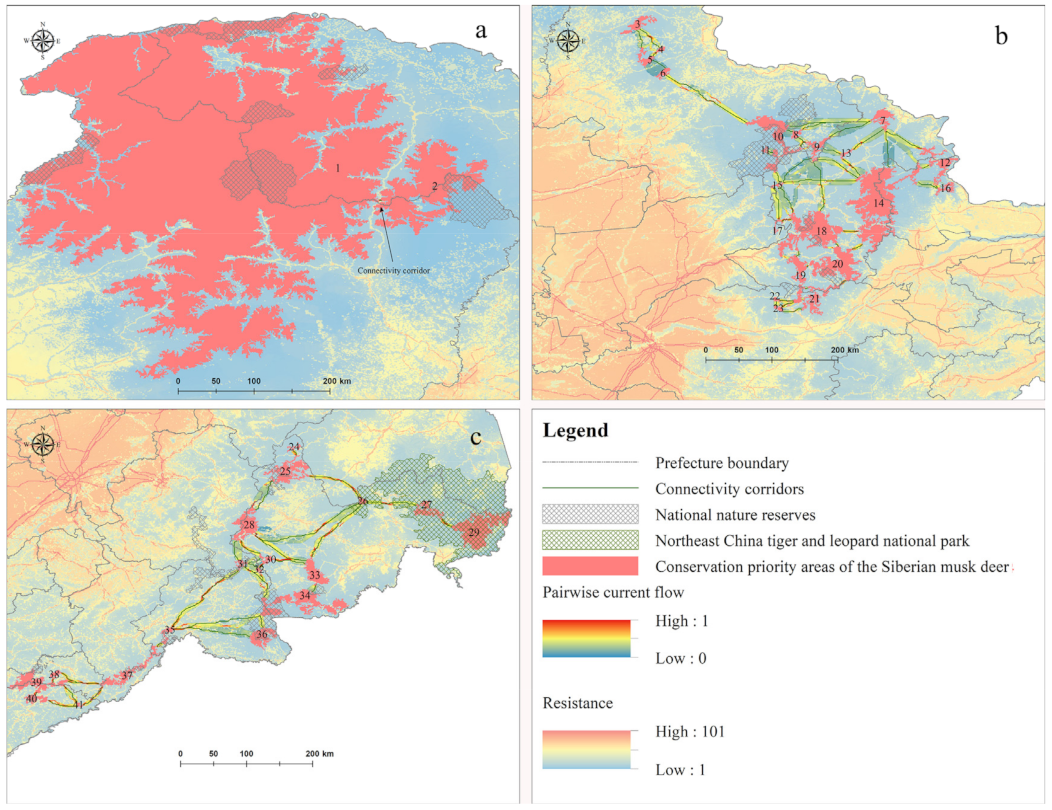
3.3. Patch Fragmentation of Core Patches and Potential Connectivity Corridors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrowski, S.; Rahmani, H.; Ali, J.; Ali, R.; Zahler, P. Musk deer Moschus cupreus persist in the eastern forests of Afghanistan. Oryx 2016, 50, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.S.; Meng, X.X.; Xia, L.; Feng, Z.J. Conservation status and causes of decline of musk deer (Moschus spp.) in China. Biol. Conserv. 2003, 109, 333–342. [Google Scholar] [CrossRef]
- Lee, W.S.; Rhim, S.J. Changes in distribution area of Korean musk deer (Moschus mo-schiferus parvipes) from 1950s to 1999 in South Korea. J. For. Res. 2002, 13, 135–136. [Google Scholar] [CrossRef]
- The IUCN Red List of Threatened Species 2015. Available online: https://www.iucnredlist.org/species/13897/61977573 (accessed on 1 October 2021).
- Sheng, H.L.; Liu, Z.X. The Musk Deer in China; Shanghai Scientific and Technical Publishers: Shanghai, China, 2007. [Google Scholar]
- Wu, J.Y.; Wang, W. The Musk Deer of China; China Foresty Publishing House: Beijing, China, 2006. [Google Scholar]
- Gao, E.H.; Yu, D.Y.; Li, Q. Status and conservation countermeasures of Musk Deer in China. For. Resour. Manag. 2005, 45–47. [Google Scholar] [CrossRef]
- Sheng, H.L. Current situation of Musk Deer Resources and rescue measures in China. Chin. J. Wildl. 1996, 10–12. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, W.S.; Zhao, W.Y.; Li, J.M. Status and administrative measures of Moschus moschiferus resources in Liaoning province. Liaoning For. Sci. Technol. 2005, 57–59. [Google Scholar] [CrossRef]
- Zhang, D.D.; Zhu, H.Q.; Jiang, C.Y.; Li, C.; Zhang, X.D.; Liu, C.F.; Qu, D.j.; Guo, H. Research Situation of Impact Factors on Habitat Selection of Siberian Moschus moschiferus. J. Econ. Anim. 2014, 18, 44–46. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Pimm, S.; Jenkins, C.; Abell, R.; Brooks, T.; Gittleman, J.; Joppa, L.; Raven, P.; Roberts, C.; Sexton, J. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Zahler, P.; Badamjav, L.; Reading, R.; Wingard, J.; Amgalanbaatar, S.; Gombobaatar, S.; Barton, N.; Yo, O. Illegal and Unsustainable Wildlife Hunting and Trade in Mongolia. Mong. J. Biol. Sci. 2004, 2, 23–31. [Google Scholar] [CrossRef]
- Slaght, J.; Milakovsky, B.; Maksimova, D.; Seryodkin, I.; Zaitsev, V.; Panichev, A.M.; Miquelle, D. Anthropogenic influences on the distribution of a Vulnerable coniferous forest specialist: Habitat selection by the Siberian musk deer Moschus moschiferus. Oryx 2017, 53, 174–180. [Google Scholar] [CrossRef]
- Tsendjav, D.; Batbold, D.O. Musk Deer Conservation: Mongolia’s Role in the International Musk Trade; WWF Mongolia Programme Office: Ulaanbaatar, Mongolia, 2003. [Google Scholar]
- Wemmer, C. Deer: Status Survey and Conservation Action Plan; IUCN: Gland, Switzerland; Cambridge, UK, 1998. [Google Scholar]
- Coad, L.; Leverington, F.; Knights, K.; Geldmann, J.; Eassom, A.; Kapos, V.; Kingston, N.; de Lima, M.; Zamora, C.; Cuardros, I.; et al. Measuring impact of protected area management interventions: Current and future use of the Global Database of Protected Area Management Effectiveness. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140281. [Google Scholar] [CrossRef]
- Yang, R.; Cao, Y.; Hou, S.Y.; Peng, Q.Y.; Wang, X.S.; Wang, F.Y.; Tseng, T.H.; Yu, L.; Carver, S.; Convery, I.; et al. Cost-effective priorities for the expansion of global terrestrial protected areas: Setting post-2020 global and national targets. Sci. Adv. 2020, 6, eabc3436. [Google Scholar] [CrossRef]
- Feng, C.T.; Cao, M.; Wang, W.; Wang, H.; Liu, F.Z.; Zhang, L.B.; Du, J.H.; Zhou, Y.; Huang, W.J.; Li, J.S. Which management measures lead to better performance of China’s protected areas in reducing forest loss? Sci. Total Environ. 2021, 764, 142895. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.X.; Liu, X.M.; Zhou, D.Q.; Ma, K.P.; Wu, Q.; Li, G.Y. Some opinions on the integration and optimization of natural protected areas in China. Biodivers. Sci. 2021, 29, 290–294. [Google Scholar] [CrossRef]
- Wang, W.; Feng, C.T.; Liu, F.Z.; Li, J.S. Biodiversity conservation in China: A review of recent studies and practices. Environ. Sci. Ecotechnol. 2020, 2, 100025. [Google Scholar] [CrossRef]
- Li, W.H. Degradation and restoration of forest ecosystems in China. For. Ecol. Manag. 2004, 201, 33–41. [Google Scholar] [CrossRef]
- Viña, A.; McConnell, W.J.; Yang, H.; Xu, Z.; Liu, J. Effects of conservation policy on China’s forest recovery. Sci. Adv. 2016, 2, e1500965. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.W.; Costanza, R.; Dai, Q.; Stoeckl, N.; Gu, X.D.; Farber, S.; Nie, Y.G.; Kubiszewski, I.; Hu, Y.B.; Swaisgood, R.; et al. The Value of Ecosystem Services from Giant Panda Reserves. Curr. Biol. 2018, 28, 2174–2180. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.m.; Feng, L.M.; Mou, P.; Wu, J.G.; Smith, J.L.D.; Xiao, W.H.; Yang, H.T.; Dou, H.L.; Zhao, X.D.; Cheng, Y.C.; et al. Amur tigers and leopards returning to China: Direct evidence and a landscape conservation plan. Landsc. Ecol. 2016, 31, 491–503. [Google Scholar] [CrossRef]
- Tang, X.P.; Jiang, Y.F.; Liu, Z.L.; Chen, J.Z.; Liang, B.K.; Lin, C. Top-level Design of the Natural Protected Area System in China. For. Resour. Manag. 2019, 1–7. [Google Scholar] [CrossRef]
- Su, X.K.; Han, W.Y.; Liu, G.H. Potential priority areas and protection network for Yunnan snub-nosed monkey (Rhinopithecus bieti) in Southwest China. J. Geogr. Sci. 2019, 29, 1211–1227. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.H. China Climate; Science Press: Beijing, China, 2013. [Google Scholar]
- Yuan, C.Y. Superiority of Forest Resources in Northeast China. J. Northeast. For. Univ. 2007, 35, 65–67. [Google Scholar] [CrossRef]
- Zhao, Z.J. Fauna of Rare and Endangered Species of Vertebrates of Northeast China; China Forestry Publishing House: Beijing, China, 1999. [Google Scholar]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from correlated climate variables: A major source of uncertainty for predicting species distributions under climate change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Williams, K.J.; Belbin, L.; Austin, M.P.; Stein, J.L.; Ferrier, S. Which environmental variables should I use in my biodiversity model? Int. J. Geogr. Inf. Sci. 2012, 26, 2009–2047. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Naimi, B.; Araújo, M. sdm: A reproducible and extensible R platform for species distribution modelling. Ecography 2016, 39, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Coetzee, B.; Robertson, M.P.; Erasmus, B.; Van Rensburg, B.; Thuiller, W. Ensemble models predict Important Bird Areas in southern Africa will become less effective for conserving endemic birds under climate change. Glob. Ecol. Biogeogr. 2009, 18, 701–710. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, L.; Wu, S.; Xia, W.; Yang, L.; Li, M.; Chen, M.; Luan, X. Use of historical data to improve conservation of the black grouse (Lyrurus tetrix) in Northeast China. Ecosphere 2020, 11, e03090. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.; Anderson, R.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.; Huettmann, F.; Leathwick, J.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Thuiller, W. BIOMOD—Optimizing predictions of species distributions and projecting potential future shifts under global change. Glob. Chang. Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Yang, L.; Shi, K.C.; Ma, C.; Ren, G.P.; Fan, P.F. Mechanisms underlying altitudinal and horizontal range contraction: The western black crested gibbon. J. Biogeogr. 2020, 48, 321–331. [Google Scholar] [CrossRef]
- Luo, M.; Wang, H.; Lyu, Z. Evaluating the performance of species distribution models Biomod2 and MaxEnt using the giant panda distribution data. Chin. J. Appl. Ecol. 2017, 28, 4001–4006. [Google Scholar] [CrossRef]
- Gallien, L.; Münkemüller, T.; Albert, C.H.; Boulangeat, I.; Thuiller, W. Predicting potential distributions of invasive species: Where to go from here? Divers. Distrib. 2010, 16, 331–342. [Google Scholar] [CrossRef]
- Zhang, X.L.; Li, Y.C.; Wang, Y.Y.; Cai, H.Y.; Zeng, H.; Wang, Z.H. Influence of future climate change in suitable habitats of tea in different countries. Biodivers. Sci. 2019, 27, 15–26. [Google Scholar] [CrossRef]
- Soberón, J.; Nakamura, M. Niches and distributional areas: Concepts, methods, and assumptions. Proc. Natl. Acad. Sci. USA 2009, 106, 19644–19650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobgay, S.; Mahavik, N. Potential habitat distribution of Himalayan red panda and their connectivity in Sakteng Wildlife Sanctuary, Bhutan. Ecol. Evol. 2020, 10, 12929–12939. [Google Scholar] [CrossRef]
- Moilanen, A.; Franco, A.M.A.; Early, R.I.; Fox, R.; Wintle, B.; Thomas, C.D. Prioritizing Multiple-Use Landscapes for Conservation: Methods for Large Multi-Species Planning Problems. Proc. R. Soc. B Biol. Sci. 2005, 272, 1885–1891. [Google Scholar] [CrossRef] [Green Version]
- Moilanen, A. Landscape Zonation, benefit functions and target-based planning: Unifying reserve selection strategies. Biol. Conserv. 2007, 134, 571–579. [Google Scholar] [CrossRef]
- Wu, J.P.; Zhang, Y.; Fu, D.F.; Lan, W.X. Comparison of Home Range of Siberian Musk Deer and Coral in Winter. J. Northeast. For. Univ. 2008, 36, 58–60. [Google Scholar]
- McGarigal, K.; Marks, B. FRAGSTATS—Spatial Pattern Analysis Program for Quantifying Landscape Structure; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1995. [Google Scholar] [CrossRef]
- Linkage Mapper Connectivity Analysis Software. Available online: http://www.circuitscape.org/linkagemapper (accessed on 1 October 2021).
- Pinchpoint Mapper Connectivity Analysis Software. Available online: http://www.circuitscape.org/linkagemapper (accessed on 1 October 2021).
- McRae, B.; Dickson, B.; Keitt, T.; Shah, V. Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 2008, 89, 2712–2724. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Oakleaf, J.R.; Theobald, D.M.; Baruch-Mordo, S.; Kiesecker, J. Managing the middle: A shift in conservation priorities based on the global human modification gradient. Glob. Chang. Biol. 2019, 25, 811–826. [Google Scholar] [CrossRef]
- Wu, J.P.; Zhang, H.L.; Zhang, Y. The Habitat Selection of Siberian Musk Deer in Winter in Daxing’an Mountains. Chin. J. Zool. 2007, 42, 45–50. [Google Scholar] [CrossRef]
- Zhang, D.D.; Zhu, H.Q.; Ge, Z.Y.; Chang, S.H.; Li, C.; Zhang, X.D. Selection of musk deer winter habitat in Huangnihe Nature Reserve. J. Northwest AF Univ. Nat. Sci. Ed. 2015, 43, 15–20. [Google Scholar] [CrossRef]
- Wu, J.P.; Zhou, L.L.; Mu, L.Q. Summer habitat selection by Siberian musk deer (Moschus moschiferus) in Tonghe forest area in the Lesser Khingan Mountains. Acta Theriol. Sin. 2006, 26, 44–48. [Google Scholar] [CrossRef]
- Jiang, F.; Zhang, J.J.; Gao, H.M.; Cai, Z.Y.; Zhou, X.W.; Li, S.Q.; Zhang, T.Z. Musk deer (Moschus spp.) face redistribution to higher elevations and latitudes under climate change in China. Sci. Total Environ. 2020, 704, 135335. [Google Scholar] [CrossRef]
- Yi, B.Z.; Ye, Y.G. The Development of Forest Reserves in Northeast China Since The End of Qing Dynasty and Its Environmental Cost. Agric. Hist. China 2004, 23, 115–123. [Google Scholar] [CrossRef]
- Butchart, S.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.; Almond, R.; Baillie, J.; Bertzky, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.W.; WANG, Z.C.; Luo, Y.M.; Wang, C.; Cai, F.K.; Gong, Z.C.; Tai, Z.J.; Sui, Y.C.; Piao, Z.J. Impact of Human Economic Activities on Animal Diversity in Changbai Mountain Nature Reserve. J. Beihua Univ. Nat. Sci. 2012, 13, 444–450. [Google Scholar]
- Li, W.; Bao, H.; Zhang, M. Habitat analysis and design of potential ecological corridors for Amur tiger in Northeastern China. Acta Theriol. Sin. 2017, 37, 317–326. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | VIF | Variable Importance | Source |
|---|---|---|---|
| Mean annual temperature | 6.261 | 0.052 | (http://www.worldclim.org/, accessed on 23 May 2020) |
| Mean annual precipitation | 2.644 | 0.237 | (http://www.worldclim.org/, accessed on 23 May 2020) |
| Elevation | 3.328 | 0.015 | (http://www.cgiar-csi.org/, accessed on 16 November 2020) |
| Slope | 1.044 | 0.021 | (http://www.cgiar-csi.org/, accessed on 16 November 2020) |
| Birch distribution | 2.854 | 0.012 | (http://www.resdc.cn/, accessed on 8 December 2017) |
| Larch distribution | 2.118 | 0.022 | (http://www.resdc.cn/, accessed on 8 December 2017) |
| Shrub distribution | 3.177 | 0.005 | (http://www.resdc.cn/, accessed on 8 December 2017) |
| Forest density | 4.095 | 0.046 | (http://www.resdc.cn/, accessed on 8 December 2017) |
| Human modification of terrestrial land | 3.145 | 0.237 | (https://sedac.ciesin.columbia.edu/, accessed on 1 October 2019) |
| Distance from roads | 1.508 | 0.002 | (https://www.webmap.cn/, accessed on 7 September 2021) |
| Settlement density | 3.442 | 0.053 | (https://www.webmap.cn/, accessed on 7 September 2021) |
| Human density | 1.295 | 0.006 | (http://www.resdc.cn/, accessed on 8 October 2020) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Fan, Y.; Chen, M.; Xia, W.; Wang, J.; Zhan, Z.; Wang, W.; Khan, T.U.; Wu, S.; Luan, X. Identification of Conservation Priority Areas and a Protection Network for the Siberian Musk Deer (Moschus moschiferus L.) in Northeast China. Animals 2022, 12, 260. https://doi.org/10.3390/ani12030260
Zhang C, Fan Y, Chen M, Xia W, Wang J, Zhan Z, Wang W, Khan TU, Wu S, Luan X. Identification of Conservation Priority Areas and a Protection Network for the Siberian Musk Deer (Moschus moschiferus L.) in Northeast China. Animals. 2022; 12(3):260. https://doi.org/10.3390/ani12030260
Chicago/Turabian StyleZhang, Chao, Yuwei Fan, Minhao Chen, Wancai Xia, Jiadong Wang, Zhenjie Zhan, Wenlong Wang, Tauheed Ullah Khan, Shuhong Wu, and Xiaofeng Luan. 2022. "Identification of Conservation Priority Areas and a Protection Network for the Siberian Musk Deer (Moschus moschiferus L.) in Northeast China" Animals 12, no. 3: 260. https://doi.org/10.3390/ani12030260