1. Introduction
The modern, intensive pig industry requires high-performing sows that are also very demanding. Over the years sows have become more prolific and produce large, heavy litters with increased daily gains and high requirements for energy consumption from milk [
1,
2,
3]. These fast-growing litters impose an energy deficit on the sow, especially when entering a catabolic state towards the end of the gestation and early lactation [
4]. Commonly, sows are fed a restricted diet shortly before, and for the first days after farrowing to prevent decreased lactation feed intake or postpartum dysgalactia syndrome [
5,
6]. Therefore, increasing energy and nutrient retention from feed is crucial to maintain performance while keeping feed costs reasonable [
2,
6]; the addition of animal fats, such as tallow or lard, can help nutritionists achieve these aims [
3]. Despite their high energy density, however, animal fats can be highly variable in their composition and quality [
7] and are associated with comparatively incomplete absorption, since they mainly contain saturated, long-chain fatty acids (FAs). These saturated long-chain fatty acids (LCFAs) are known to be less readily incorporated in the micelles of the intestinal lumen [
8,
9], a prerequisite for exposure to the activity of pancreatic lipases [
10,
11].
Addition of digestive enzymes and emulsifiers within the feed of both sows and fattening pigs has also been used to aid in digestion, absorption, and retention of fat and other nutrients [
9,
12]. Lysolecithin (LL) is one of the most studied emulsifying agents added to improve digestibility [
13]. Usually produced by processing soybean oil [
14,
15], commercially available LLs contain diverse mixtures of phospholipids (PLs), lysophospholipids (LPLs), triglycerides (TGs) and other compounds [
16]. It has been reported that LL emulsifying properties vary depending on its source and dietary fat composition, essentially on the ratio of saturated vs. unsaturated FAs [
8,
13,
17,
18], though this is debated when the wider effects of husbandry are considered [
19]. At the same time, LL can interact with the structure and function of biological membranes, further enhancing nutrient absorption by the intestinal epithelial cells [
14,
16].
The sensitive performance and metabolic balance of sows on transition from gestation to lactation and the possible positive effect of LL supplementation has been a topic of research. Zhao et al. [
20] studied the effect of LL addition (beginning on gestation day 110) to a low-animal fat or a high-animal fat diet given to gestating/lactating sows on their performance, litter weight gain, milk composition and total tract digestibility of energy and nutrients, while Wang et al. [
21] also explored the impact of linearly increasing levels of the same as above LL-containing product (administered from gestation day 107) on performance and digestibility indices. They furthermore studied the respective impact on milk/colostrum immunoglobulin concentrations and on selected blood parameters. However, the feed did not contain animal fat and sows were allowed ad libitum consumption during the entire lactation period.
Overall, it could be hypothesized that dietary supplementation of lysolecithin during the demanding period of the last part of gestation and throughout lactation in sows could affect their metabolism and performance by improving digestibility and absorption of fat as well as other nutrients. In this context, the objective of the study was to of LL supplementation on an animal fat-containing diet from three weeks before expected farrowing and during the lactation period, on sow and litter performance and on selected blood metabolism associated parameters in sows.
3. Results
The effects of treatment on the performance parameters of sows and piglets are summarized in
Table 2 and
Table 3. Bodyweights differed neither on gestation day 108 nor at weaning. Bodyweight loss during those five weeks rose to almost a quarter of their initial weight. Sows of the CG group had a significantly
(p = 0.030) thicker backfat layer than those of the LLG group at d108 and d14; however, the difference in backfat thickness between the groups gradually disappeared towards weaning. Particularly between d14 and weaning, the backfat loss was smaller in LLG than in CG (
p = 0.006). The degree of backfat tissue mobilization did not affect their post-weaning reproductive performance, as sows of both groups came to estrus soon and almost concurrently: 5.83 vs. 6.03 d after weaning, for groups CG and LLG, respectively (
p = 0.834).
Litter size did not differ between the groups. Losses due to mortality during lactation were within acceptable criteria and comparable between groups. Litter weights were similar at birth, but litters of sows whose feed was LL-supplemented arrived significantly heavier at weaning (p = 0.027).
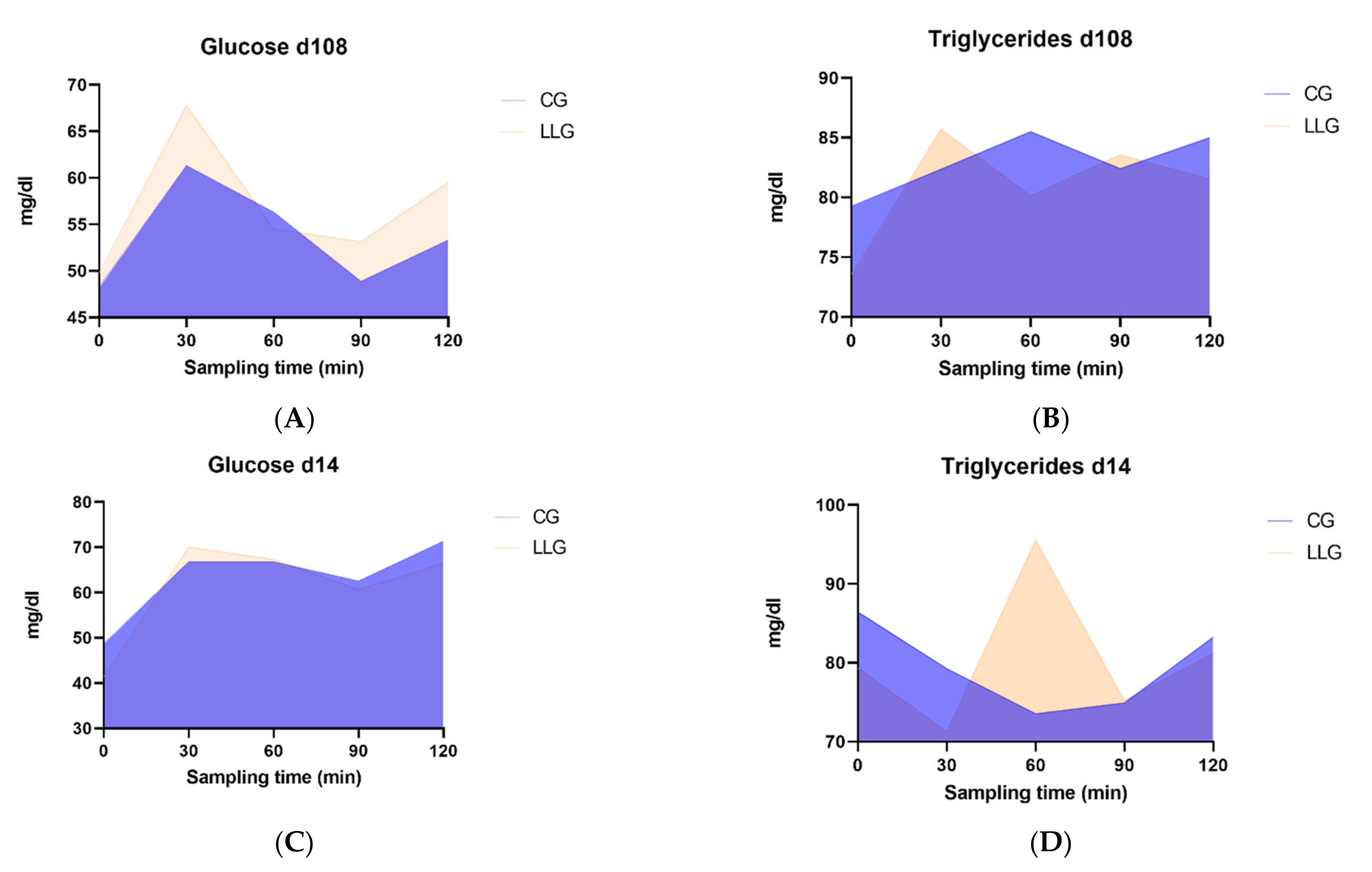
The concentration-time profiles of glucose and triglycerides in blood following the morning meal on gestation day 108 and on lactation day 14 are given in
Table 4 and
Table 5, respectively, while the AUC are provided in
Figure 1. Although data from gestation day 108 revealed no differences in glucose between the two groups, pre-prandial levels on lactation day 14 showed a trend (
p = 0.074) to be lower in the LLG animals. No differences were discerned between the treatments at any time point, on any day for the triglyceride concentrations. Repeated measures analysis of glucose and triglycerides levels collected at the end of gestation showed no significant treatment effect (
p = 0.262 for glucose and
p = 0.225 for triglycerides), no significant interaction between treatment and time of sampling (
p = 0.626 for glucose and
p = 0.492 for triglycerides) and a significant time effect (
p < 0.001 for glucose and
p = 0.044 for triglycerides). Moreover, repeated measures analysis of glucose and triglycerides levels collected at mid-lactation showed no significant treatment effect (
p = 0.420 for glucose and
p = 0.851 for triglycerides), no significant interaction between treatment and time of sampling (
p = 0.583 for glucose and
p = 0.247 for triglycerides) and a significant time effect for glucose (
p < 0.001) but not for triglycerides (
p = 0.247).
On gestation day 108, differences in urea NEFA and creatinine blood concentrations between the two groups were not statistically significant (
Table 6). Leptin concentration was significantly higher (
p = 0.021) in the animals of the CG group compared to LLG. This was not the case on lactation day 14, when differences in leptin, along with NEFA and creatinine, were non-significant between CG and LLG groups. On the contrary, urea levels were significantly (
p = 0.002) elevated in the animals of group LLG.
4. Discussion
Sows during the last third of gestation have dramatically increased needs for dietary energy [
3,
20]. Two thirds of total fetal growth occur within this period, uterine blood flow rises, and the development of insulin resistance leads to an increase in blood glucose levels and fetal glucose uptake. Lactate and NEFA blood levels are also elevated towards the end of gestation, indicating a negative energy balance and mobilization of fat (and protein) reserves, even in cases where ad libitum feeding is allowed [
4]. Subsequent milk production is also highly energetically demanding, and means sows need both a suitable dietary energy supply and improved digestion, absorption, and retention processes. The present study explored how LL supplementation throughout the last third of gestation and the entire lactation period could support performance.
According to our results, LL did not significantly affect the body weight of sows at either time point. Backfat thickness of sows belonging to the LLG group was lower than that of sows of the CG group at gestation day 108 and lactation day 14, implying a similar peripartal loss rate; however, this was not the case at weaning, since animals of both groups had similar backfat reserves, probably signifying a reduction in body fat loss during the last half of lactation due to the addition of LL in the feed. Indeed, it was revealed that sows in the LLG group mobilized less backfat than CG sows during the last half of the lactation period. The second half of the lactation period is the period with the highest metabolic demands for the sow, because milk production is maximized as well as piglet growth. A lower backfat mobilization during this crucial period, implies that the sows in the LLG group may have been able to tolerate the energy needs, by not only mobilizing body reserves, but most probably by an improved digestion and metabolic efficiency of nutrients due to lysolecithin supplementation. In the study of Laws et al. [
22] sows supplemented with palm oil, lost less backfat than those who were not supplemented. The reduction in backfat loss was attributed to increased dietary energy intake during lactation. Elsewhere, supplementing various levels of soybean lecithin oil in lactating sows’ diets, did not affect backfat mobilization during lactation [
23]. In the latter study, backfat was measured at farrowing and at weaning, and therefore no indication of backfat mobilization during the mid-lactation could be noted. In accordance with our findings, lysophospholipid supplementation in sows from day 107 of gestation onwards, resulted in a reduction of body weight and backfat loss during lactation [
21]. This effect was probably mediated by an improved availability of nutrients due to the action of lysophospholipid. It is plausible that lysolecithin also improved digestibility and availability of nutrients in the present study. Such an effect could be supported by the increased urea levels in the LLG group during lactation, which may suggest an improved utilization of absorbed amino acids, irrespective of lean body mass mobilization, since that would have led to a change in blood creatinine. Whereas it is commonly considered that the primary function of amino acids from dietary protein is to build proteins in the body, many amino acids have an important role in the sow’s metabolism around farrowing when insulin sensitivity is low: they provide glucogenic substrate to the citric acid cycle [
24]. The practical relevance of an improved nutrient utilization in the case of lysolecithin should be reflected in litter growth during lactation. Indeed, litter weight at weaning was higher in the LLG group. This finding agrees with those of Wang et al. [
21], who reported improved litter weight gain in the lysophospholipid supplemented group. Similarly, supplementation of soybean lecithin in sow diets during gestation and lactation resulted in improved piglet weight at weaning [
23].
In the study of Zhao et al. [
20], the addition of LL in beef tallow-containing sow diets led to a significant reduction in body weight and backfat loss between farrowing and weaning due to increased digestibility of dry matter, nitrogen, gross energy and crude fat. Due to the different measurement schedule, though, it is difficult to assess the exact point in lactation when the positive effect of LL becomes more prominent. It must be stressed that a direct comparison with our results must be carried out cautiously, since no animal fats were included in the feed and, as aforementioned, a significant interaction between LL activity and fat type is to be expected.
The positive results observed on piglet weight at weaning could also be attributed to possible effects on composition of sow milk and colostrum. Zhao et al. [
20] reported that the addition of LL in beef tallow-containing sow diets can lead to an increase in the fat and lactose content of the milk, depending on the stage of lactation. Wang et al. [
21] also reported a positive effect of LL supplementation on IgG/A/M levels in sow colostrum and milk and on IgA/M levels in piglet blood. An increase in immunoglobulin (mainly IgA) content of the colostrum, milk, sow blood and suckling piglet blood was confirmed by Shi et al. [
23] who, furthermore, found a respective increase in the levels of phosphatidylcholine and total PLs and a significantly decreased diameter of fat globules in the milk of sows fed on a lecithin-enforced diet. Therefore, it is plausible that the increased litter weight at weaning in our study could be attributed to an elevated energy/nutrient content of the consumed milk, combined with its increased digestibility and immunoglobulin levels, offering passive immunity and resilience to bacterial and viral pathogens and to food antigens [
23].
Glucose is the main energy source for both the sow and her litter [
4,
25]. Its homeostasis balances on the dietary carbohydrate intake, the metabolic needs and a set of hormones (e.g., insulin and glucagon) and organs (e.g., kidneys, liver) that regulate its utilization, storage and elimination. According to our results, the preprandial glucose concentrations on lactation day 14 were lower in the animals of the LLG group, although the difference with the CG group did not reach statistical significance. This could generally indicate either a reduction in the assimilation of feed nutrients or a better peripheral utilization. Lower blood glucose levels in sows have been associated with an improved insulin sensitivity [
24]. Wang et al. [
21] reported increased blood glucose levels due to lysophospholipid supplementation. Nevertheless, in the latter study blood parameters were evaluated at approximately 3 h postprandially and cannot be fully compared to our results. According to Laws et al. [
22] dietary fat supplementation in sows can result in elevated glucose concentrations because of a fat-induced glucose intolerance. In our study we have used commercially relevant fat levels, nearly half the supplemented quantity of those used by Laws et al. [
22]. Presumably if fat levels were higher than the current ones, the differences in the pre- and postprandial levels of glucose and triglycerides may have become significant. Furthermore, the delayed interval for the peak value of triglyceride concentration in the LLG group both at day 110 of gestation and at day 14 of lactation, may indicate a prolonged digestion and absorption procedure of dietary lipids due to a more efficient emulsification procedure and breakdown to smaller micelles by the action of lysolecithin. This field warrants further investigation.
A promising finding was detected for postprandial urea levels during lactation reflecting the use of amino acids as energy source, which were significantly higher in the LLG sows. We postulate that the addition of LL in the feed results in a trend for more sustained absorption and systematic delivery at least of proteins at a rate which allows for a better peripheral utilization. This effect could be attributed to the overall improvement of crude protein digestibility along with other nutrients, as demonstrated by Wang et al. [
21] in the case of lysophospholipid supplementation.
Leptin is a hormone produced by adipocytes. It acts as a regulator of energy homeostasis and serves as an indicator of body condition since its levels reflect body fat stores [
2,
24]. On gestation day 108, significantly higher leptin concentration in the blood of sows of group CG were probably correlated with increased backfat thickness. On lactation day 14, although backfat at the P2 position was still significantly thicker in the CG group, the divergence was probably beginning to drop enough (due to the decrease in the fat loss rate in the animals of the LLG group), so that the respective difference in leptin concentrations was not deemed significant. Nevertheless, the physiological significance of the higher leptin levels in the CG group may be related to a post-metabolic effect on the growth of the progeny. It was previously shown that significant negative correlations exist between ante-partum leptin levels and litter weight and litter growth during lactation [
26]. Interestingly, in the CG group the higher leptin levels ante-partum co-existed with the lower litter weight at weaning.


 ,
, 

{kind=link}