
A Culture-Based Study of Micromycetes Isolated from the Urban Nests of Grey Heron (Ardea cinerea) in SW Poland

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection
2.3. Isolation of Micromycetes from Samples Using Culture-Dependent Approach
2.3.1. Procedure of Placing Biological Material on Petri Dishes
2.3.2. A Damp Chamber Procedure
2.4. Fungal Identification
2.5. Data Analyses
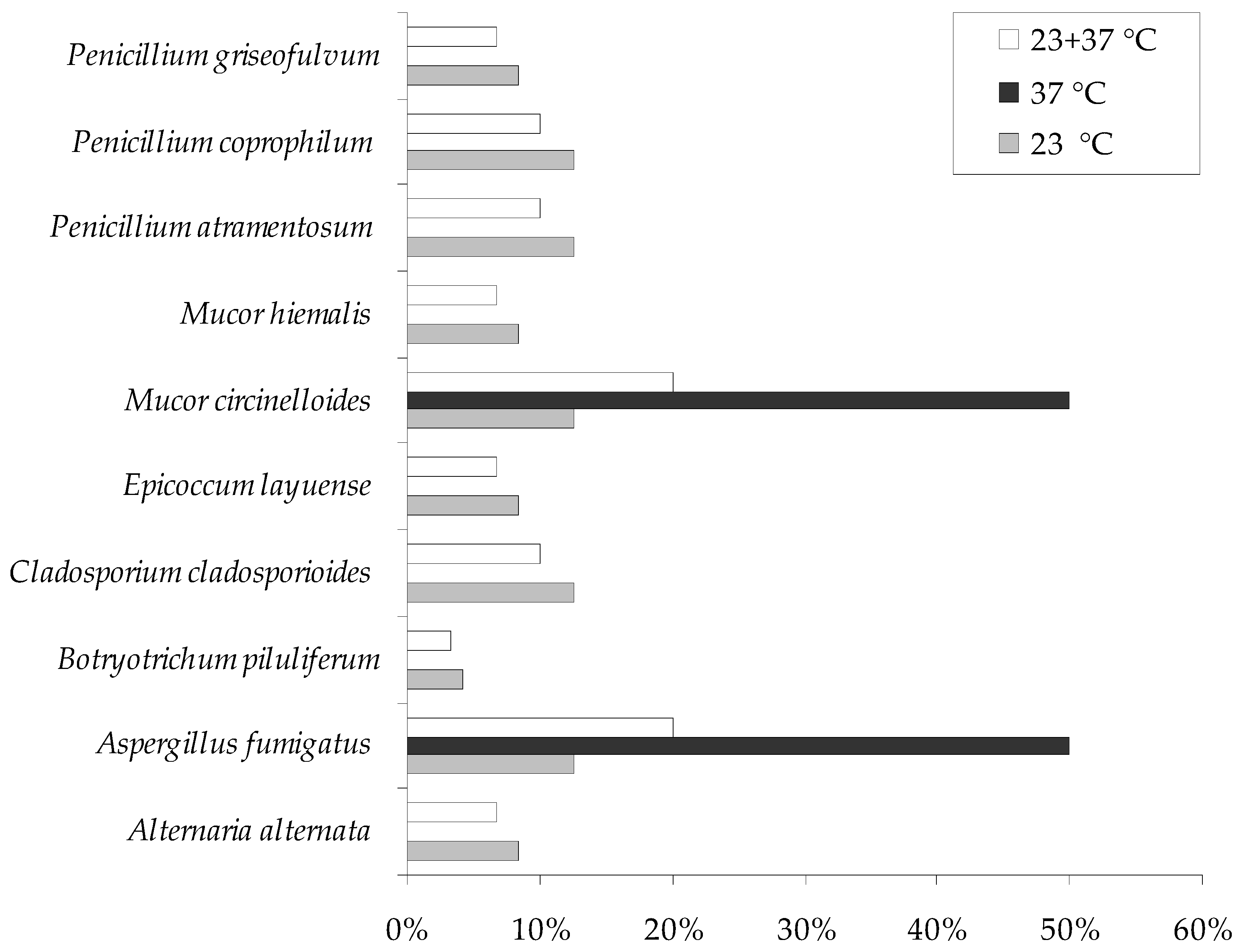
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jusino, M.A.; Lindner, D.L.; Banik, M.T.; Walters, J.R. Heart rot hotel: Fungal communities in red-cockaded woodpecker excavations. Fungal Ecol. 2015, 14, 33–43. [Google Scholar] [CrossRef]
- Jusino, M.A.; Lindner, D.L.; Banik, M.T.; Rose, K.R.; Walters, J.R. Experimental evidence of a symbiosis between red-cockaded woodpeckers and fungi. Proc. R. Soc. B 2016, 283, 20160106. [Google Scholar] [CrossRef]
- Caiafa, M.V.; Jusino, M.A.; Wilkie, A.C.; Díaz, I.A.; Sieving, K.E.; Smith, M.E. Discovering the role of Patagonian birds in the dispersal of truffles and other mycorrhizal fungi. Curr. Biol. 2021, 31, 5558–5570.e3. [Google Scholar] [CrossRef]
- Beernaert, L.A.; Pasmans, F.; Van Waeyenberghe, L.; Haesebrouck, F.; Martel, A. Aspergillus infections in birds: A review. Avian Pathol. 2010, 39, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Quesada, Ó.; Rodríguez, F.; Herráez, P.; Seara, D.; de los Monteros, A.E. Mucor ramosissimus associated with feather loss in canaries (Serinus canarius). Avian Dis. Dig. 2007, 2, e25. [Google Scholar] [CrossRef] [Green Version]
- Simi, W.B.; Leite, D.P., Jr.; Paula, C.R.; Hoffmann-Santos, H.D.; Takahara, D.T.; Hahn, R.C. Yeasts and filamentous fungi in psittacidae and birds of prey droppings in midwest region of Brazil: A potential hazard to human health. Braz. J. Biol. 2019, 79, 414–422. [Google Scholar] [CrossRef]
- Cafarchia, C.; Romito, D.; Coccioli, C.; Camarda, A.; Otranto, D. Phospholipase activity of yeasts from wild birds and possible implications for human disease. Med. Mycol. 2008, 46, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Elliott, T.F.; Jusino, M.A.; Trappe, J.M.; Lepp, H.; Ballard, G.A.; Bruhl, J.J.; Vernes, K. A global review of the ecological significance of symbiotic associations between birds and fungi. Fungal Divers. 2019, 98, 161–194. [Google Scholar] [CrossRef]
- Watkins, R.A.; King, J.S.; Johnston, S.A. Nutritional Requirements and Their Importance for Virulence of Pathogenic Cryptococcus Species. Microorganisms 2017, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Oladele, R.O.; Ayanlowo, O.O.; Richardson, M.D.; Denning, D.W. Histoplasmosis in Africa: An emerging or a neglected disease? PLoS Negl. Trop. Dis. 2018, 12, e0006046. [Google Scholar] [CrossRef]
- Refai, M.K.; El-Hariri1and, M.; Alarousy, R. Cryptococcosis in Animals and Birds: A Review. Eur. J. Acad. Res. 2017, 4, 202–223. [Google Scholar]
- Deem, S.L. Fungal diseases of birds of prey. Vet Clin. N. Am. Exot. Anim. Pract. 2003, 6, 363–376. [Google Scholar] [CrossRef]
- Vázquez-González, D.; Perusquía-Ortiz, A.M.; Hundeiker, M.; Bonifaz, A. Opportunistic yeast infections: Candidiasis, cryptococcosis, trichosporonosis and geotrichosis. J. Dtsch. Dermatol. Ges. 2013, 11, 381–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubálek, Z. Influence of pH on the occurrence of fungi in bird’s nests. Z. Allg. Mikrobiol. 1976, 16, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Korniłłowicz-Kowalska, T.; Kitowski, I.; Iglik, H. Geophilic dermatophytes and other keratinophilic fungi in the nests of wetland birds. Acta Mycol. 2011, 46, 83–107. [Google Scholar] [CrossRef] [Green Version]
- Korniłłowicz-Kowalska, T.; Kitowski, I.; Bohacz, J.; Kwiatkowska, E. Fungal Frequency and Diversity in the Nests of Wetland Birds from Poland: Relationships between Birds, Nest Properties and Inhabiting Fungi. Avian Biol. Res. 2018, 11, 245–262. [Google Scholar] [CrossRef]
- Korniłłowicz-Kowalska, T.; Kitowski, I. Aspergillus fumigatus and other thermophilic fungi in nests of wetland birds. Mycopathologia 2013, 175, 43–56. [Google Scholar] [CrossRef]
- Żółkoś, K.; Meissner, W.; Kalisiński, M.; Górska, E.; Melin, M.; Ibron, I.; Wysocki, D. Liczebność i rozmieszczenie kolonii czapli siwej Ardea cinerea w północnej Polsce. Ornis Pol. 2010, 51, 30–42. (In Polish) [Google Scholar]
- Wylegała, P.; Batycki, A.; Mizera, T. Liczebność i rozmieszczenie stanowisk czapli siwej Ardea cinerea w Wielkopolsce w latach 2006–2010. Ornis Pol. 2011, 52, 75–85. (In Polish) [Google Scholar]
- Kasprzykowski, Z.; Goławski, A. Elementy ekologii rozrodu bąka Botaurus stellaris i czapli siwej Ardea cinerea na stawach hodowlanych w Siedlcach. Ornis Pol. 2014, 55, 257–263. (In Polish) [Google Scholar]
- Przymencki, M.; Litwiniak, K.; Celiński, D. Śródmiejska kolonia czapli siwej Ardea cinerea we Wrocławiu. Ornis Pol. 2019, 60, 221–226. (In Polish) [Google Scholar]
- Beuch, S.; Biwo, T.; Grochowski, P.; Czechowski, P.; Lenkiewicz, W.; Przymencki, M.; Wasiak, P.; Betleja, J.; Bzoma, S. Aktualny stan populacji lęgowej czapli siwej Ardea cinerea i kormorana Phalacrocorax carbo w Śląskim Regionie Ornitologicznym. Ptaki Śląska 2021, 27, 5–24. (In Polish) [Google Scholar]
- Litwiniak, K.; Przymencki, M. An attempt of wintering of Leptoglossus occidentalis Heidemann, 1910 (Hemiptera: Heteroptera: Coreidae) in Grey Heron Ardea cinerea nests. Heteroptera Pol.–Acta Faun. 2021, 15, 125–127. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulkifli, N.A.; Zakaria, L. Morphological and Molecular Diversity of Aspergillus from Corn Grain Used as Livestock Feed. HAYATI J. Biosci. 2017, 24, 26–34. [Google Scholar] [CrossRef]
- Wang, X.W.; Houbraken, J.; Groenewald, J.Z.; Meijer, M.; Andersen, B.; Nielsen, K.F.; Crous, P.W.; Samson, R.A. Diversity and taxonomy of Chaetomium and chaetomium-like fungi from indoor environments. Stud. Mycol. 2016, 84, 145–224. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.A.; Wiederhold, N.P.; Cano-Lira, J.F.; Guarro, J. New species of Cladosporium associated with human and animal infections. Persoonia 2016, 36, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Hou, L.W.; Duan, W.J.; Crous, P.W.; Cai, L. Didymellaceae revisited. Stud. Mycol. 2017, 87, 105–159. [Google Scholar] [CrossRef]
- Wagner, L.; Stielow, J.B.; de Hoog, G.S.; Bensch, K.; Schwartze, V.U.; Voigt, K.; Alastruey-Izquierdo, A.; Kurzai, O.; Walther, G. A new species concept for the clinically relevant Mucor circinelloides complex. Persoonia 2020, 44, 67–97. [Google Scholar] [CrossRef]
- Samson, R.A.; Hong, S.; Peterson, S.W.; Frisvad, J.C.; Varga, J. Polyphasic taxonomy of Aspergillus section Fumigati and its teleomorph Neosartorya. Stud. Mycol. 2007, 59, 147–203. [Google Scholar] [CrossRef]
- Visagie, C.M.; Renaud, J.B.; Burgess, K.M.; Malloch, D.W.; Clark, D.; Ketch, L.; Urb, M.; Louis-Seize, G.; Assabgui, R.; Sumarah, M.W.; et al. Fifteen new species of Penicillium. Persoonia 2016, 36, 247–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Ogórek, R.; Dyląg, M.; Kozak, B. Dark stains on rock surfaces in Driny Cave (Little Carpathian Mountains, Slovakia). Extremophiles 2016, 20, 641–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubálek, Z. Keratinophilic fungi associated with free-living mammals and birds. In Biology of Dermatophytes and Other Keratinophilic Fungi; Kushwaha, R.K.S., Guarro, J., Eds.; Revista Iberoamericana de Micología: Bilbao, Spain, 2000; pp. 93–103. [Google Scholar]
- Baggott, G.K.; Graeme-Cook, K. Microbiology of natural incubation. In Avian Incubation: Behaviour, Environment and Evolution; Deeming, D.C., Ed.; Oxford University Press: Oxford, UK, 2002; pp. 179–191. [Google Scholar]
- Deshmukh, K.R.; Pethe, A.S. Effect of Sunlight on Growth and Melanin Pigment Production in Bacteria and Fungi. IJIRSET 2017, 6, 1668–1670. [Google Scholar] [CrossRef]
- Liu, S.; Youngchim, S.; Zamith-Miranda, D.; Nosanchuk, J.D. Fungal Melanin and the Mammalian Immune System. J. Fungi 2021, 7, 264. [Google Scholar] [CrossRef] [PubMed]
- Pasanen, A.L. A review: Fungal exposure assessment in in-door environments. Indoor Air 2001, 11, 87–98. [Google Scholar] [CrossRef]
- Rastogi, G.; Sani, R.K. Molecular techniques to assess microbial community structure, function, and dynamics in the environment. In Microbes and Microbial Technology; Ahmad, I., Farah, A., John, P., Eds.; Springer Science & Business Media, Agricultural and Environmental Applications: Berlin/Heidelberg, Germany, 2011; pp. 29–57. [Google Scholar]
- MacNeil, L.; Kauri, T.; Robertson, W. Molecular techniques and their potential application in monitoring the microbiological quality of indoor air. Can. J. Microbiol. 1995, 41, 657–675. [Google Scholar] [CrossRef]
- Wu, P.C.; Su, H.J.; Ho, H.M. A comparison of sampling media for environmental viable fungi collected in a hospital environment. Environ. Res. 2000, 82, 253–257. [Google Scholar] [CrossRef]
- Macher, J.M. Review of methods to collect settled dust and isolate culturable microorganisms. Indoor Air 2001, 11, 99–110. [Google Scholar] [CrossRef]
- Ogórek, R.; Kozak, B.; Višňovská, Z.; Tančinová, D. Phenotypic and genotypic diversity of airborne fungal spores in Demänovská Ice Cave (Low Tatras, Slovakia). Aerobiologia 2018, 34, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Marshall, V.; Poulson-Cook, S.; Moldenhauer, J. Comparative mold and yeast recovery analysis (the effect of differing incubation temperature ranges and growth media). PDA J. Pharm. Sci. Technol. 1998, 52, 165–169. [Google Scholar]
- Meletiadis, J.; Meis, J.F.G.M.; Mouton, J.W.; Verweij, P.E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 2001, 39, 478–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uthayasooriyan, M.; Pathmanathan, S.; Ravimannan, N.; Sathyaruban, S. Formulation of alternative culture media for bacterial and fungal grow. Der. Pharm. Lett. 2016, 8, 431–436. [Google Scholar]
- Ogórek, R.; Kalinowska, K.; Pląskowska, E.; Baran, E.; Moszczyńska, E. Zanieczyszczenia powietrza grzybami na różnych podłożach hodowlanych w wybranych pomieszczeniach kliniki dermatologicznej. Część 1. Mikol. Lek. 2011, 18, 30–38. (In Polish) [Google Scholar]
- Litman, M.L. A culture medium for the primary isolation of fungi. Science 1947, 106, 109–111. [Google Scholar] [CrossRef]
- Ruben, J. The evolution of endothermy in mammals and birds—From physiology to fossils. Ann. Rev. Physiol. 1995, 57, 69–95. [Google Scholar] [CrossRef]
- Kornillowicz-Kowalska, T.; Kitowski, I. Nests of Marsh harrier (Circus aeruginosus L.) as refuges of potentially phytopathogenic and zoopathogenic fungi. Saudi J. Biol. Sci. 2018, 25, 136–143. [Google Scholar] [CrossRef]
- Korniłłowicz-Kowalska, T.; Kitowski, I. Diversity of fungi in nests and pellets of Montagu’s harrier (Circus pygargus) from eastern Poland—importance of chemical and ecological factors. Ecol. Chem. Eng. S 2009, 16, 453–471. [Google Scholar]
- Shim, S.H.; Swenson, D.C.; Gloer, J.B.; Dowd, P.F.; Wicklow, D.T.; Penifulvin, A. A Sesquiterpenoid-Derived Metabolite Containing a Novel Dioxa [5,5,5,6] fenestrane Ring System from a Fungicolous Isolate of Penicillium griseofulvum. Org. Lett. 2006, 8, 1225–1228. [Google Scholar] [CrossRef]
- Nigam, P.S. Production of Secondary Metabolites–Fungi. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C., Patel, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 570–578. [Google Scholar]
- Aninat, C.; Hayashi, Y.; André, F.; Delaforge, M. Molecular Requirements for Inhibition of Cytochrome P450 Activities by Roquefortine. Chem. Res. Toxicol. 2001, 14, 1259–1265. [Google Scholar] [CrossRef]
- Aninat, C.; André, F.; Delaforge, M. Oxidative metabolism by P450 and function coupling to efflux systems: Modulation of mycotoxin toxicity. Food Addit. Contam. 2005, 22, 361–368. [Google Scholar] [CrossRef]
- Kokkonen, M.; Jestoi, M.; Rizzo, A. The effect of substrate on mycotoxin production of selected Penicillium strains. Int. J. Food Microbiol. 2005, 99, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Rabha, J.; Dhruva, K.J. Metabolic Diversity of Penicillium. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Rodriguez-Couto, S., Eds.; Elsevier Books: Amsterdam, The Netherlands, 2018; pp. 217–234. [Google Scholar]
- Aho, R.; Westerling, B.; Ajello, L.; Padhye, A.A.; Samson, R.A. Avian penicilliosis caused by Penicillium griseofulvum in a captive toucanet. J. Med. Vet. Mycol. 1990, 28, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Kozakiewicz, Z.; Smith, D. Physiology of Aspergillus. In Biotechnology Hand-Books 7. Aspergillus; Atkins, R.F., Sherwood, R.F., Eds.; Plenum Press: New York, NY, USA, 1994; p. 23. [Google Scholar]
- Nwagu, T.; Okolo, B.N. Growth Profile and Amylase Hydrolytic Activity of a Thermophilic Fungi Aspergillus fumigatus Isolated from Soil. Asian J. Biotechnol. 2011, 3, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Olias, P.; Gruber, A.D.; Hafez, M.H.; Lierz, M.; Slesiona, S.; Brock, M.; Jacobsen, I.D. Molecular epidemiology and virulence assessment of Aspergillus fumigatus isolates from white stork chicks and their environment. Vet. Microbiol. 2011, 148, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Hattab, Z.; Ben Lasfar, N.; Abid, M.; Bellazreg, F.; Fathallah, A.; Hachfi, W.; Letaief, A. Alternaria alternata infection causing rhinosinusitis and orbital involvement in an immunocompetent patient. New Microbes New Infect. 2019, 32, 100561. [Google Scholar] [CrossRef]
- Wiest, P.; Wiese, K.; Jacobs, M.R.; Morrissey, A.B.; Abelson, T.I.; Witt, W.; Lederman, M.M. Alternaria Infection in a Patient with Acquired Immunodeficiency Syndrome: Case Report and Review of Invasive Alternaria Infections. Rev. Infect. Dis. 1987, 9, 799–803. [Google Scholar] [CrossRef]
- Hoog, G.S. Atlas of Clinical Fungi, 2nd ed.; Amer Society for Microbiology: Utrecht, The Netherlands, 2000; pp. 1–1126. [Google Scholar]
- Kantarcioǧlu, A.S.; Yücel, A.; Hoog, G.S. Case report. Isolation of Cladosporium cladosporioides from cerebrospinal fluid. Mycoses 2002, 45, 500–503. [Google Scholar] [CrossRef]
- Ogórek, R.; Lejman, A.; Pusz, W.; Miłuch, A.; Miodyńska, P. Characteristics and taxonomy of Cladosporium fungi. Mik. Lek. 2012, 19, 80–85. [Google Scholar]
- Jakubas, D. Factors affecting the breeding success of the grey heron (Ardea cinerea) in northern Poland. J. Ornithol. 2005, 146, 27–33. [Google Scholar] [CrossRef]
- Kanegusuku, A.G.; Stankovic, I.N.; Cote-Hammarlof, P.A.; Yong, P.H.; White-Ziegler, C.A. A Shift to Human Body Temperature (37 °C) Rapidly Reprograms Multiple Adaptive Responses in Escherichia coli That Would Facilitate Niche Survival and Colonization. J. Bacteriol. 2021, 203, e00363-21. [Google Scholar] [CrossRef]
- Macura, A.B.; Skóra, M.; Krzyściak, P. Atlas Grzybów Chorobotwórczych Człowieka; Medpharm: Wrocław, Poland, 2011; pp. 66–83. (In Polish) [Google Scholar]
- Chander, J. Mucormycosis. In Textbook of Medical Mycology, 4th ed.; Jaypee Brothers Medical Publishers Ltd.: New Delhi, India, 2018; pp. 534–596. [Google Scholar]
- Magraya, A.R.; Ganaiab, B.A.; Ahmad, F. Isolation, identification and pathogenicity patterns of Mucor hiemalis in cultured Cyprinis carpio communis using challenged system. Aquaculture 2020, 518, 734837. [Google Scholar] [CrossRef]
- Rajachan, O.; Kanokmedhakul, K.; Soytong, K.; Kanokmedhakul, S. Mycotoxins from the Fungus Botryotrichum piluliferum. J. Agric. Food Chem. 2017, 65, 1337–1341. [Google Scholar] [CrossRef] [PubMed]
- Del Frari, G.; Cabral, A.; Nascimento, T.; Boavida Ferreira, R.; Oliveira, H.; Sarrocco, S. Epicoccum layuense a potential biological control agent of esca-associated fungi in grapevine. PLoS ONE 2019, 14, e0213273. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Fungi | Isolate | GenBank Accession No. | The Sequence Length (bp) | Identity with Sequence from GenBank | ||||
|---|---|---|---|---|---|---|---|---|
| Query Cover, % | Identity, % | Accession | Isolate | |||||
| Procedure of placing biological material on Petri dishes | Alternaria alternata | UWR_210 | MW763150 | 475 | 100 | 100.00 | MN615420 | YZU 191238 |
| Aspergillus fumigatus | UWR_211 | MW763151 | 505 | 100 | 100.00 | MT597427 | MEBP0074 | |
| Botryotrichum piluliferum | UWR_212 | MW763152 | 386 | 100 | 99.48 | MH857131 | CBS 131.53 | |
| Cladosporium cladosporioides | UWR_213 | MW763153 | 458 | 100 | 100.00 | MT598826 | T1 | |
| Epicoccum layuense | UWR_214 | MW763154 | 460 | 100 | 100.00 | MT573479 | 17 | |
| Mucor circinelloides | UWR_215 | MW763155 | 567 | 100 | 100.00 | MT603954 | CMRC 573 | |
| Mucor hiemalis | UWR_216 | MW763156 | 523 | 100 | 100.00 | MT366055 | NG4 | |
| Penicillium atramentosum | UWR_217 | MW763157 | 446 | 100 | 100.00 | MK281569 | SCAU199 | |
| Penicillium coprophilum | UWR_218 | MW763158 | 512 | 100 | 100.00 | MT410465 | Mzz9 | |
| Penicillium griseofulvum | UWR_262 | MZ391135 | 490 | 100 | 100.00 | MH006592 | MPR1 | |
| A damp chamber procedure | Aspergillus fumigatus | UWR_297 | OL657062 | 400 | 100 | 100.00 | MT597433 | MEBP0082 |
| Penicillium coprophilum | UWR_263 | MZ391136 | 487 | 100 | 100.00 | MK450685 | CMV005G5 | |
| Penicillium griseofulvum | UWR_264 | MZ391137 | 440 | 100 | 100.00 | KY069877 | PPRI_22701 | |
| Fungi | 23 ± 0.5 °C | 37 ± 0.5 °C | ||||
|---|---|---|---|---|---|---|
| Nest with Visible Fungal Growth | Nest without Visible Fungal Growth | L. occidentalis | Nest with Visible Fungal Growth | Nest without Visible Fungal Growth | L. occidentalis | |
| Alternaria alternata | x | x | ||||
| Aspergillus fumigatus | x | x | x | x | x | x |
| Botryotrichum piluliferum | x | |||||
| Cladosporium ladosporioides | x | x | x | |||
| Epicoccum layuense | x | x | ||||
| Mucor circinelloides | x | x | x | x | x | x |
| Mucor hiemalis | x | x | ||||
| Penicillium atramentosum | x | x | x | |||
| Penicillium coprophilum | x | x | x | |||
| Penicillium griseofulvum | x | x | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogórek, R.; Borzęcka, J.; Kłosińska, K.; Piecuch, A.; Przymencki, M.; Litwiniak, K.; Suchodolski, J. A Culture-Based Study of Micromycetes Isolated from the Urban Nests of Grey Heron (Ardea cinerea) in SW Poland. Animals 2022, 12, 676. https://doi.org/10.3390/ani12060676
Ogórek R, Borzęcka J, Kłosińska K, Piecuch A, Przymencki M, Litwiniak K, Suchodolski J. A Culture-Based Study of Micromycetes Isolated from the Urban Nests of Grey Heron (Ardea cinerea) in SW Poland. Animals. 2022; 12(6):676. https://doi.org/10.3390/ani12060676
Chicago/Turabian StyleOgórek, Rafał, Justyna Borzęcka, Katarzyna Kłosińska, Agata Piecuch, Marcin Przymencki, Klaudia Litwiniak, and Jakub Suchodolski. 2022. "A Culture-Based Study of Micromycetes Isolated from the Urban Nests of Grey Heron (Ardea cinerea) in SW Poland" Animals 12, no. 6: 676. https://doi.org/10.3390/ani12060676
APA StyleOgórek, R., Borzęcka, J., Kłosińska, K., Piecuch, A., Przymencki, M., Litwiniak, K., & Suchodolski, J. (2022). A Culture-Based Study of Micromycetes Isolated from the Urban Nests of Grey Heron (Ardea cinerea) in SW Poland. Animals, 12(6), 676. https://doi.org/10.3390/ani12060676