
A New Disease Caused by an Unidentified Etiological Agent Affects European Salamanders

 ,
,  , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
“Нo как же этo так? Ведь этo же чудoвищнo! Этo чудoвищнo, гoспoда”, -пoвтoрил oн, oбращаясь к жабам в террарии, нo жабы спали и ничегo ему не oтветили.” (“But how can it be? It’s monstrous! Quite monstrous, gentlemen,” he repeated, addressing the toads in the terrarium, who were asleep and made no reply).M. Bulgakov, Рoкoвые яйца
2. Materials and Methods
2.1. The Fire Salamander: Study Area and Monitoring
2.2. Cyst Extraction
2.3. Cyst Cell Isolation
2.4. Histology
2.5. Electron Microscopy
2.6. Scanning Electron Microscopy
2.7. Genetic Analyses
2.8. Ethics
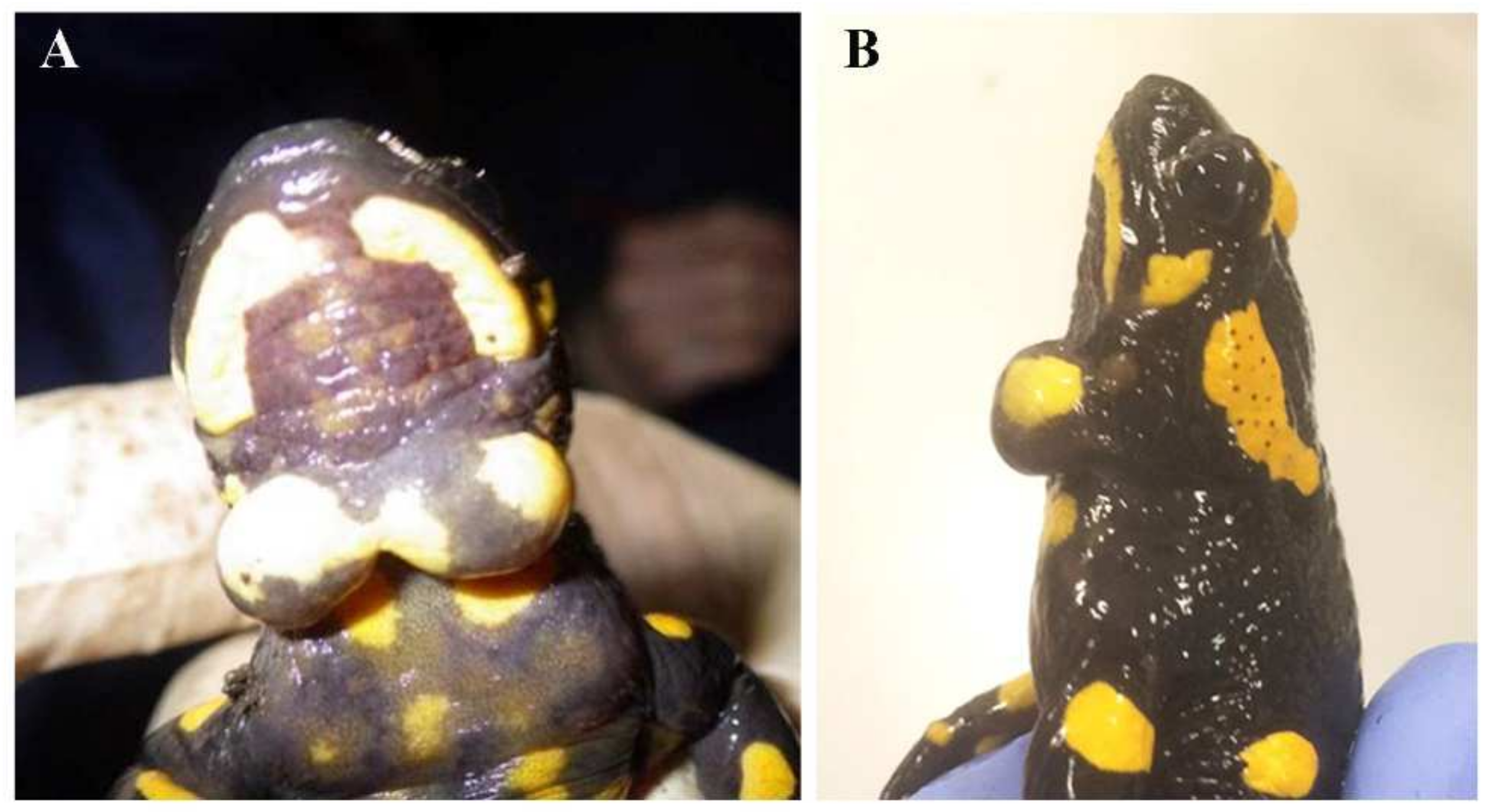
3. Results
3.1. Cysts Occurrence in Fire Salamander Populations
3.2. Histological Analysis
3.3. In Vitro Isolation
3.4. Electron Microscopy
3.5. DNA Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cleaveland, S.; Haydon, D.T.; Taylor, L. Overviews of pathogen emergence: Which pathogens emerge, when and why? Curr. Top. Microbiol. Immunol. 2007, 315, 85–111. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.M.; Wang, X.D.; Ye, H.; Wang, H.M.; Qiu, S.; Zhang, H.M.; Liu, Y.; Luo, J.H.; Feng, J. Does public fear that bats spread COVID-19 jeopardize bat conservation? Biol. Conserv. 2021, 254, 9. [Google Scholar] [CrossRef] [PubMed]
- Maurin, M.; Fenollar, F.; Mediannikov, O.; Davoust, B.; Devaux, C.; Raoult, D. Current Status of Putative Animal Sources of SARS-CoV-2 Infection in Humans: Wildlife, Domestic Animals and Pets. Microorganisms 2021, 9, 868. [Google Scholar] [CrossRef] [PubMed]
- Schrag, S.J.; Wiener, P. Emerging infectious disease: What are the relative roles of ecology and evolution? Trends Ecol. Evol. 1995, 10, 319–324. [Google Scholar] [CrossRef]
- Murray, A.G.; Smith, R.J.; Stagg, R.M. Shipping and the spread of infectious salmon anemia in Scottish aquaculture. Emerg. Infect. Dis. 2002, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Minhas, V. Wildlife population management: Are contraceptive vaccines a feasible proposition? Front. Biosci. (Sch. Ed.) 2017, 9, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, I.F.; Lawson, B.; Goharriz, H.; Rodriguez-Ramos Fernandez, J.; John, S.K.; Fooks, A.R.; Cunningham, A.A.; Johnson, N.; Horton, D.L. Extension of the known distribution of a novel clade C betacoronavirus in a wildlife host. Epidemiol. Infect. 2019, 147, e169. [Google Scholar] [CrossRef] [Green Version]
- Morner, T.; Obendorf, D.L.; Artois, M.; Woodford, M.H. Surveillance and monitoring of wildlife diseases. Rev. Sci. Tech. 2002, 21, 67–76. [Google Scholar] [CrossRef]
- Verner-Carlsson, J.; Lohmus, M.; Sundstrom, K.; Strand, T.M.; Verkerk, M.; Reusken, C.; Yoshimatsu, K.; Arikawa, J.; van de Goot, F.; Lundkvist, A. First evidence of Seoul hantavirus in the wild rat population in the Netherlands. Infect. Ecol. Epidemiol. 2015, 5, 27215. [Google Scholar] [CrossRef] [Green Version]
- Niederle, M.V.; Bosch, J.; Ale, C.E.; Nader-Macias, M.E.; Aristimuno Ficoseco, C.; Toledo, L.F.; Valenzuela-Sanchez, A.; Soto-Azat, C.; Pasteris, S.E. Skin-associated lactic acid bacteria from North American bullfrogs as potential control agents of Batrachochytrium dendrobatidis. PLoS ONE 2019, 14, e0223020. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A. Amphibian immunity-stress, disease, and climate change. Dev. Comp. Immunol. 2017, 66, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daszak, P.; Berger, L.; Cunningham, A.A.; Hyatt, A.D.; Green, D.E.; Speare, R. Emerging infectious diseases and amphibian population declines. Emerg. Infect. Dis. 1999, 5, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Martel, A.; Spitzen-van der Sluijs, A.; Blooi, M.; Bert, W.; Ducatelle, R.; Fisher, M.C.; Woeltjes, A.; Bosman, W.; Chiers, K.; Bossuyt, F.; et al. Batrachochytrium salamandrivorans sp nov causes lethal chytridiomycosis in amphibians. Proc. Natl. Acad. Sci. USA 2013, 110, 15325–15329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, L.J.; Garner, T.W.J.; Tessa, G.; Scheele, B.C.; Griffiths, A.G.F.; Wilfert, L.; Harrison, X.A. An emerging viral pathogen truncates population age structure in a European amphibian and may reduce population viability. PeerJ 2018, 6, e5949. [Google Scholar] [CrossRef] [PubMed]
- Duffus, A.L.; Andrews, A.M. Phylogenetic analysis of a frog virus 3-like ranavirus found at a site with recurrent mortality and morbidity events in southeastern Ontario, Canada: Partial major capsid protein sequence alone is not sufficient for fine-scale differentiation. J. Wildl. Dis. 2013, 49, 464–467. [Google Scholar] [CrossRef] [Green Version]
- Duffus, A.L.J.; Garner, T.W.J.; Nichols, R.A.; Standridge, J.P.; Earl, J.E. Modelling Ranavirus Transmission in Populations of Common Frogs (Rana temporaria) in the United Kingdom. Viruses 2019, 11, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoverman, J.T.; Gray, M.J.; Haislip, N.A.; Miller, D.L. Phylogeny, life history, and ecology contribute to differences in amphibian susceptibility to ranaviruses. Ecohealth 2011, 8, 301–319. [Google Scholar] [CrossRef]
- Mihaljevic, J.R.; Hoverman, J.T.; Johnson, P.T.J. Co-exposure to multiple ranavirus types enhances viral infectivity and replication in a larval amphibian system. Dis. Aquat. Organ. 2018, 132, 23–35. [Google Scholar] [CrossRef]
- Ayres, C.; Acevedo, I.; Monsalve-Carcano, C.; Thumsova, B.; Bosch, J. Triple dermocystid-chytrid fungus-ranavirus co-infection in a Lissotriton helveticus. Eur. J. Wildl. Res. 2020, 66, 1–3. [Google Scholar] [CrossRef]
- Gonzalez-Hernandez, M.; Denoel, M.; Duffus, A.J.L.; Garner, T.W.J.; Cunningham, A.A.; Acevedo-Whitehouse, K. Dermocystid infection and associated skin lesions in free-living palmate newts (Lissotriton helveticus) from Southern France. Parasitol. Int. 2010, 59, 344–350. [Google Scholar] [CrossRef]
- Martínez-Silvestre, A.; Fernandez-Guiberteau, D.; Pérez-Sorribes, L.; Velarde, R. Infección por dermocistidios en Lissotriton helveticus en Cataluña: Nuevos datos y apuntes sobre su diagnóstico. Boletín Asoc. ÓN Herpetológica Española 2017, 28, 66–69. [Google Scholar]
- Mendoza, L.; Taylor, J.W.; Ajello, L. The class mesomycetozoea: A heterogeneous group of microorganisms at the animal-fungal boundary. Annu. Rev. Microbiol. 2002, 56, 315–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamovicz, L.; Woodburn, D.B.; Virrueta Herrera, S.; Low, K.; Phillips, C.A.; Kuhns, A.R.; Crawford, J.A.; Allender, M.C. Characterization of Dermotheca sp. Infection in a midwestern state-endangered salamander (Ambystoma platineum) and a co-occurring common species (Ambystoma texanum). Parasitology 2020, 147, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Fagotti, A.; Rossi, R.; Canestrelli, D.; La Porta, G.; Paracucchi, R.; Lucentini, L.; Simoncelli, F.; Di Rosa, I. Longitudinal study of Amphibiocystidium sp. infection in a natural population of the Italian stream frog (Rana italica). Parasitology 2019, 146, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Manenti, R.; Ficetola, G.F.; De Bernardi, F. Water, stream morphology and landscape: Complex habitat determinants for the fire salamander Salamandra salamandra. Amphib.-Reptil. 2009, 30, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Manenti, R.; Conti, A.; Pennati, R. Fire salamander (Salamandra salamandra) males’ activity during breeding season: Effects of microhabitat features and body size. Acta Herpetol. 2017, 12, 29–36. [Google Scholar]
- Adamska, M.; Leonska-Duniec, A.; Maciejewska, A.; Sawczuk, M.; Skotarczak, B. Comparison of efficiency of variuous DNA extraction methods from cysts of Giardia intestinalis measured by PCR and Taqman Real Time PCR. Parasite 2010, 17, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Raffel, T.R.; Bommarito, T.; Barry, D.S.; Witiak, S.M.; Shackelton, L.A. Widespread infection of the Eastern red-spotted newt (Notophthalmus viridescens) by Amphibiocystidium, a genus of a new species of fungus-like mesomycetozoan parasites not previously reported in North America. Parasitology 2008, 135, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Karnati, S.K.R.; Yu, Z.; Sylvester, J.T.; Dehority, B.A.; Morrison, M.; Firkins, J.L. Technical note: Specific PCR amplification of protozoal 18S rDNA sequences from DNA extracted from ruminal samples of cows. J. Anim. Sci. 2003, 81, 812–815. [Google Scholar] [CrossRef]
- Shin, E.C.; Cho, K.M.; Lim, W.J.; Hong, S.Y.; An, C.L.; Kim, E.J.; Kim, Y.K.; Choi, B.R.; An, J.M.; Kang, J.M.; et al. Phylogenetic analysis of protozoa in the rumen contents of cow based on the 18S rDNA sequences. J. Appl. Microbiol. 2004, 97, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Shanan, S.; Abd, H.; Bayoumi, M.; Saeed, A.; Sandstrom, G. Prevalence of Protozoa Species in Drinking and Environmental Water Sources in Sudan. Biomed. Res. Int. 2015, 2015, 345619. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. the characterization of enzymatically amplified eukaryotic 16S-Like RRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Petroni, G.; Dini, F.; Verni, F.; Rosati, G. A molecular approach to the tangled intrageneric relationships underlying phylogeny in Euplotes (Ciliophora, Spirotrichea). Mol. Phylogenetics Evol. 2002, 22, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, M.; Uriz, M.J.; Taberlet, P.; Coissac, E.; Wangensteen, O.S.; Turon, X. Deep-Sea, Deep-Sequencing: Metabarcoding Extracellular DNA from Sediments of Marine Canyons. PLoS ONE 2016, 11, e0139633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA for Biodiversity Research and Monitoring; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Ficetola, G.F.; Boyer, F.; Valentini, A.; Bonin, A.; Meyer, A.; Dejean, T.; Gaboriaud, C.; Usseglio-Polatera, P.; Taberlet, P. Comparison of markers for the monitoring of freshwater benthic biodiversity through DNA metabarcoding. Mol. Ecol. 2021, 30, 3189–3202. [Google Scholar] [CrossRef] [PubMed]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. OBITOOLS: A UNIX-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Zinger, L.; Bonin, A.; Alsos, I.G.; Balint, M.; Bik, H.; Boyer, F.; Chariton, A.A.; Creer, S.; Coissac, E.; Deagle, B.E.; et al. DNA metabarcoding-Need for robust experimental designs to draw sound ecological conclusions. Mol. Ecol. 2019, 28, 1857–1862. [Google Scholar] [CrossRef] [Green Version]
- Ficetola, G.F.; Pansu, J.; Bonin, A.; Coissac, E.; Giguet-Covex, C.; De Barba, M.; Gielly, L.; Lopes, C.M.; Boyer, F.; Pompanon, F.; et al. Replication levels, false presences and the estimation of the presence/absence from eDNA metabarcoding data. Mol. Ecol. Resour. 2015, 15, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Densmore, C.L.; Green, D.E. Diseases of amphibians. ILAR J. 2007, 48, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Ficetola, G.F.; Mulargia, M.; Cogoni, R.; Veith, M.; Corti, C.; Manenti, R. Batracobdella leeches, environmental features and Hydromantes salamanders. Int. J. Parasitol. Parasites Wildl. 2018, 7, 48–53. [Google Scholar] [CrossRef]
- Divar, M.R.; Sharifiyazdi, H.; Kafi, M. Application of polymerase chain reaction for fetal gender determination using cervical mucous secretions in the cow. Vet. Res. Commun. 2012, 36, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Rumpho, M.E.; Mujer, C.V.; Andrews, D.L.; Manhart, J.R.; Pierce, S.K. Extraction of DNA from mucilaginous tissues of a sea slug (Elysia chlorotica). Biotechniques 1994, 17, 1097–1101. [Google Scholar]
- Brunelli, E.; Sperone, E.; Maisano, M.; Tripepi, S. Morphology and ultrastructure of the gills in two Urodela species: Salamandrina terdigitata and Triturus carnifex. Ital. J. Zool. 2009, 76, 158–164. [Google Scholar] [CrossRef]
- Brunelli, E.; Tripepi, S. Effects of low pH acute exposure on survival and gill morphology in Triturus italicus larvae. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Spitzen-van der Sluijs, A.; Stegen, G.; Bogaerts, S.; Canessa, S.; Steinfartz, S.; Janssen, N.; Bosman, W.; Pasmans, F.; Martel, A. Post-epizootic salamander persistence in a disease-free refugium suggests poor dispersal ability of Batrachochytrium salamandrivorans. Sci. Rep. 2018, 8, 3800. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manenti, R.; Mercurio, S.; Melotto, A.; Barzaghi, B.; Epis, S.; Tecilla, M.; Pennati, R.; Scarì, G.U.; Ficetola, G.F. A New Disease Caused by an Unidentified Etiological Agent Affects European Salamanders. Animals 2022, 12, 696. https://doi.org/10.3390/ani12060696
Manenti R, Mercurio S, Melotto A, Barzaghi B, Epis S, Tecilla M, Pennati R, Scarì GU, Ficetola GF. A New Disease Caused by an Unidentified Etiological Agent Affects European Salamanders. Animals. 2022; 12(6):696. https://doi.org/10.3390/ani12060696
Chicago/Turabian StyleManenti, Raoul, Silvia Mercurio, Andrea Melotto, Benedetta Barzaghi, Sara Epis, Marco Tecilla, Roberta Pennati, Giorgio Ulisse Scarì, and Gentile Francesco Ficetola. 2022. "A New Disease Caused by an Unidentified Etiological Agent Affects European Salamanders" Animals 12, no. 6: 696. https://doi.org/10.3390/ani12060696