
Evaluating the Behavioural Responses of Healthy Newborn Calves to a Thoracic Squeeze

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of the Animals
2.2. Procedures
2.3. Treatments
2.4. Criteria for the Discontinuation of the Squeeze
2.5. Data Collection
2.6. Data Analysis
3. Results
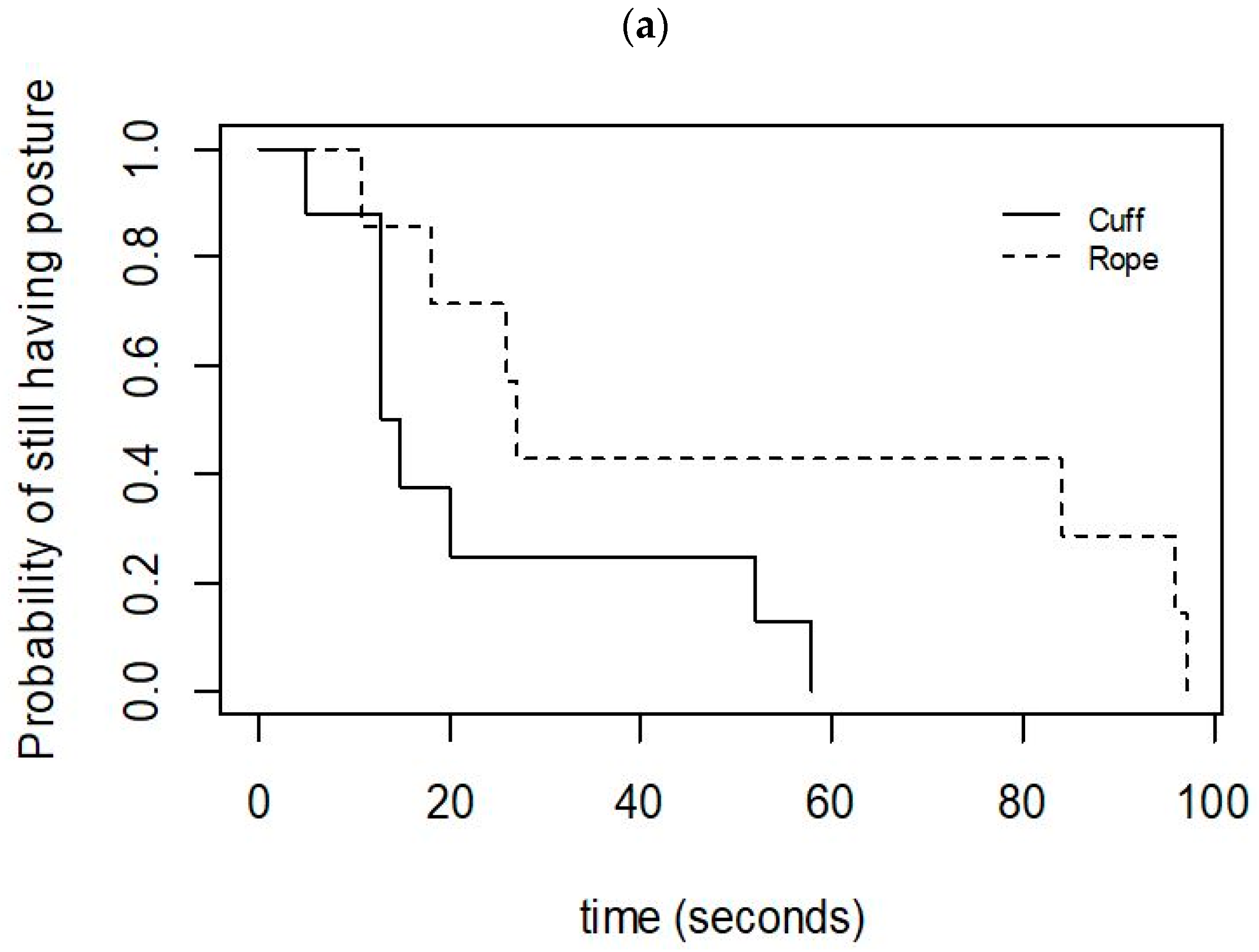
3.1. Success and Maintenance of Induction
3.2. Relationship between the Induction Category, Time to Induction, and Behaviour during the Squeeze
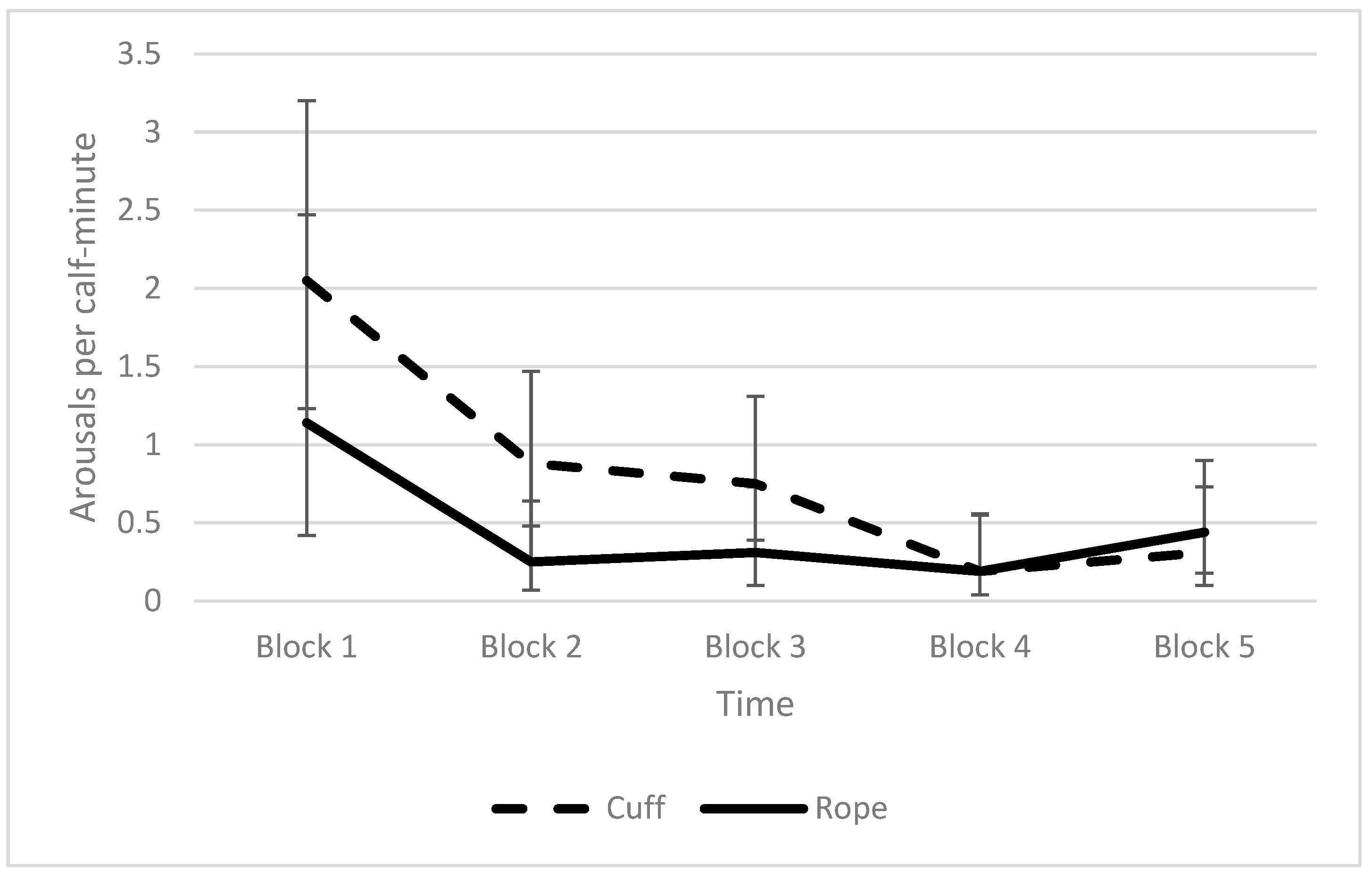
3.3. Effect of the Method of Application on the Induction, Discontinuation, and Behaviour during the Squeeze
4. Discussion
4.1. Behavioural Responses
4.2. Potential Mechanisms Underlying the Thoracic Squeeze
4.3. Comparison of the Cuff and Rope Methods
4.4. Possible Applications for the Thoracic Squeeze
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellor, D.; Lentle, R.G. Survival implications of the development of behavioural responsiveness and awareness in different groups of mammalian young. N. Z. Vet. J. 2015, 63, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Tuchscherer, M.; Puppe, B.; Tuchscherer, A.; Tiemann, U. Early identification of neonates at risk: Traits of newborn piglets with respect to survival. Theriogenology 2000, 54, 371–388. [Google Scholar] [CrossRef]
- Aleman, M.; Weich, K.; Madigan, J. Survey of veterinarians using a novel physical compression squeeze procedure in the man-agement of neonatal maladjustment syndrome in foals. Animals 2017, 7, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, E.M.; Jarvis, S.; D’Eath, R.B.; Ross, D.W.; Robson, S.K.; Farish, M.; Nevison, I.M.; Lawrence, A.B.; Edwards, S.A. Investigating the behavioural and physiological indicators of neonatal survival in pigs. Theriogenology 2008, 69, 773–783. [Google Scholar] [CrossRef]
- Mellor, D.; Stafford, K. Animal welfare implications of neonatal mortality and morbidity in farm animals. Vet. J. 2004, 168, 118–133. [Google Scholar] [CrossRef]
- Laster, D.B.; Gregory, K.E. Factors Influencing Peri- and Early Postnatal Calf Mortality. J. Anim. Sci. 1973, 37, 1092–1097. [Google Scholar] [CrossRef] [Green Version]
- Raboisson, D.; Delor, F.; Cahuzac, E.; Gendre, C.; Sans, P.; Allaire, G. Perinatal, neonatal, and rearing period mortality of dairy calves and replacement heifers in France. J. Dairy Sci. 2013, 96, 2913–2924. [Google Scholar] [CrossRef] [Green Version]
- Barrier, A.; Ruelle, E.; Haskell, M.; Dwyer, C. Effect of a difficult calving on the vigour of the calf, the onset of maternal behaviour, and some behavioural indicators of pain in the dam. Prev. Vet. Med. 2011, 103, 248–256. [Google Scholar] [CrossRef]
- Murray, C.F.; Leslie, K.E. Newborn calf vitality: Risk factors, characteristics, assessment, resulting outcomes and strategies for improvement. Vet. J. 2013, 198, 322–328. [Google Scholar] [CrossRef]
- Kolkman, I.; de Vliegher, S.; Hoflack, G.; Van Aert, M.; Laureyns, J.; de Kruif, A.; Opsomer, G. Caesarean section. Slov. Vet. Res. 2006, 43 (Suppl. 10), 329. [Google Scholar]
- Stilwell, G.; Mellor, D.J.; Holdsworth, S.E. Potential benefit of a thoracic squeeze technique in two newborn calves delivered by caesarean section. N. Z. Vet. J. 2019, 68, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J. Transitions in neuroinhibition and neuroactivation in neurologically mature young at birth, including the potential role of thoracic compression during labour. Aleman, M.; Weich, KM.; Madigan, J.E. Survey of veterinarians using a novel physical compression squeeze procedure in the management of neonatal maladjustment syndrome in foals. Animals 2017, 7, 69–81. [Google Scholar]
- Flora, T.; Smallman, M.; Kutzler, M.A. Resuscitation Compression for Newborn Sheep. Vet. Clin. N. Am. Food Anim. Pract. 2021, 37, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, S.E.; Kells, N.J.; Chidgey, K.L.; Vallée, E.; Ward, N.; Mellor, D.J.; Beausoleil, N.J. Characterisation of the Behavioural Effects of a Thoracic Squeeze in Healthy Newborn Piglets. Animals 2021, 11, 2465. [Google Scholar] [CrossRef] [PubMed]
- Mee, J.F. Managing the Calf at Calving Time. In Proceedings of the 41st Annual Conference American Association of Bovine Practitioners, Charlotte, NC, USA, 25–27 September 2008. [Google Scholar]
- Silva, B.T.; Henklein, A.; Marques, R.d.S.; de Oliveira, P.L.; Leite, S.B.P.; Novo, S.M.F.; Baccili, C.C.; dos Reis, J.F.; Gomes, V. Vital parameters of Holstein calves from birth to weaning. Rev. Bras. Med. Vet. 2016, 38, 229–304. [Google Scholar]
- Campler, M.; Munksgaard, L.; Jensen, M. The effect of housing on calving behavior and calf vitality in Holstein and Jersey dairy cows. J. Dairy Sci. 2015, 98, 1797–1804. [Google Scholar] [CrossRef] [Green Version]
- Doornbos, D.E.; Bellows, R.A.; Burfening, P.J.; Knapp, B.W. Effects of Dam Age, Prepartum Nutrition and Duration of Labor on Productivity and Postpartum Reproduction in Beef Females. J. Anim. Sci. 1984, 59, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schuenemann, G.; Nieto, I.; Bas, S.; Galvão, K.; Workman, J. Assessment of calving progress and reference times for obstetric intervention during dystocia in Holstein dairy cows. J. Dairy Sci. 2011, 94, 5494–5501. [Google Scholar] [CrossRef] [Green Version]
- Piccione, G.; Casella, S.; Pennisi, P.; Giannetto, C.; Costa, A.; Caola, G. Monitoring of physiological and blood parameters during perinatal and neonatal period in calves. Arq. Bras. Med. Veterinária Zootec. 2010, 62, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cook, C.; Devine, C.; Gilbert, K.; Tavener, A.; Day, A. Electroencephalograms and electrocardiograms in young bulls following upper cervical vertebrae-to-brisket stunning. N. Z. Vet. J. 1991, 39, 121–125. [Google Scholar] [CrossRef]
- Yakan, S.; Aksoy, O.; Ermutlu, C.S. Comparison of Use of Isoflurane or Sevoflurane for Anaesthesia Induced by Mask in Calves. Acta Sci. Vet. 2020, 48, 1–9. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. RStudio; RStudio Team Inc.: Boston, MA, USA, 2018. [Google Scholar]
- Toth, B.; Aleman, M.; Brosnan, R.J.; Dickinson, P.J.; Conley, A.; Stanley, S.D.; Nogradi, N.; Williams, C.D.; Madigan, J.E. Evaluation of squeeze-induced somnolence in neonatal foals. Am. J. Vet. Res. 2012, 73, 1881–1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, A.; Clutton, R. Are neuromuscular blocking agents being misused in laboratory pigs? Br. J. Anaesth. 2016, 116, 476–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeven, M.T.W.; Gerritzen, M.A.; Hellebrekers, L.J.; Kemp, B. Indicators used in livestock to assess unconsciousness after stunning: A review. Animal 2015, 9, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, W.N. Syncope. N. Engl. J. Med. 2000, 343, 1856–1862. [Google Scholar] [CrossRef]
- Hainsworth, R. Syncope: What is the trigger? Br. Med. J. Heart 2003, 89, 123–124. [Google Scholar] [CrossRef] [Green Version]
- Wieling, W.; Thijs, R.D.; Van Dijk, N.; Wilde, A.A.M.; Benditt, D.G.; Van Dijk, J.G. Symptoms and signs of syncope: A review of the link between physiology and clinical clues. Brain 2009, 132, 2630–2642. [Google Scholar] [CrossRef]
- Monassi, C.R.; Leite-Panissi, C.R.A.; Menescal-De-Oliveira, L. Ventrolateral periaqueductal gray matter and the control of tonic immobility. Brain Res. Bull. 1999, 50, 201–208. [Google Scholar] [CrossRef]
- Ong, R.M. Tonic Immobility by Dogs: A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of Master of Veterinary Science in Veterinary Ethology at Massey University. Ph.D. Thesis, Massey University, Palmerston North, NZ, USA, 1993. [Google Scholar]
- Brandão, M.L.; Zanoveli, J.; Ruiz-Martinez, R.C.; Oliveira, L.C.; Landeira-Fernandez, J. Different patterns of freezing behavior organized in the periaqueductal gray of rats: Association with different types of anxiety. Behav. Brain Res. 2008, 188, 1–13. [Google Scholar] [CrossRef]
- Terpou, B.A.; Harricharan, S.; McKinnon, M.C.; Frewen, P.; Jetly, R.; Lanius, R.A. The effects of trauma on brain and body: A unifying role for the midbrain periaqueductal gray. J. Neurosci. Res. 2019, 97, 1110–1140. [Google Scholar] [CrossRef]
- Gentle, M.J.; Jones, R.; Woolley, S.C. Physiological changes during tonic immobility in Gallus gallus var domesticus. Physiol. Behav. 1989, 46, 843–847. [Google Scholar] [CrossRef]
- Valance, D.; Després, G.; Richard, S.; Constantin, P.; Mignon-Grasteau, S.; Leman, S.; Boissy, A.; Faure, J.-M.; Leterrier, C. Changes in Heart Rate Variability during a tonic immobility test in quail. Physiol. Behav. 2008, 93, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Webster, D.G.; Lanthorn, T.H.; Dewsbury, D.A.; Meyer, M.E. Tonic immobility and the dorsal immobility response in twelve species of muroid rodents. Behav. Neural Biol. 1981, 31, 32–41. [Google Scholar] [CrossRef]
- Vieira, E.B.; Menescal-De-Oliveira, L.; Leite-Panissi, C.R.A. Functional mapping of the periaqueductal gray matter involved in organizing tonic immobility behavior in guinea pigs. Behav. Brain Res. 2011, 216, 94–99. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description |
|---|---|
| Latency to loss of posture | Time from the start of cuff inflation or rope tightening to enter lateral recumbency from a standing position. |
| Latency to cessation of movement | Time from the start of cuff inflation or rope tightening to the cessation of all limb and head movements for 3 s. |
| Pedal reflex | Status of reflex determined by pinching the interdigital cleft between the claws of a front foot to elicit withdrawal of the limb. Present was assigned for a full limb withdrawal response; Reduced for a slight limb withdrawal or a limb twitch response; and Absent for no muscle twitch or limb movement. |
| Palpebral reflex | Status of reflex determined by lightly brushing or touching the eyelashes or skin in the lateral corner of the eye, to elicit a blink or eye twitch response. Present was assigned for a full twitch or blink response; Reduced for a small twitch response; and Absent for no muscle twitch. |
| Low-intensity arousals | Low intensity, short duration movements. Characterised by eye opening and vigorous limb and head movements for 10 s or less, before a state of reduced responsiveness was resumed. |
| High-intensity arousals | High intensity movements, lasting longer than 5 s. Characterised by opening of the eyes and vigorous movements with righting onto all four feet. |
| Category | A (Not Induced) | B (Induced but Discontinued) | C (Induced and Maintained) | D (Induced but Unstable) | Total |
|---|---|---|---|---|---|
| Rope | 2 | 2 | 4 | 0 | 8 |
| Cuff | 1 | 2 | 3 | 2 | 8 |
| Total | 3 | 4 | 7 | 2 | 16 |
| Percentage (95% CI) | 19% (4–46%) | 25% (7–52%) | 44% (20–70%) | 12% (2–38%) | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holdsworth, S.E.; Kells, N.J.; Vallée, E.; Ward, N.; Mellor, D.J.; Beausoleil, N.J. Evaluating the Behavioural Responses of Healthy Newborn Calves to a Thoracic Squeeze. Animals 2022, 12, 840. https://doi.org/10.3390/ani12070840
Holdsworth SE, Kells NJ, Vallée E, Ward N, Mellor DJ, Beausoleil NJ. Evaluating the Behavioural Responses of Healthy Newborn Calves to a Thoracic Squeeze. Animals. 2022; 12(7):840. https://doi.org/10.3390/ani12070840
Chicago/Turabian StyleHoldsworth, Sophia E., Nikki J. Kells, Emilie Vallée, Neil Ward, David J. Mellor, and Ngaio J. Beausoleil. 2022. "Evaluating the Behavioural Responses of Healthy Newborn Calves to a Thoracic Squeeze" Animals 12, no. 7: 840. https://doi.org/10.3390/ani12070840
APA StyleHoldsworth, S. E., Kells, N. J., Vallée, E., Ward, N., Mellor, D. J., & Beausoleil, N. J. (2022). Evaluating the Behavioural Responses of Healthy Newborn Calves to a Thoracic Squeeze. Animals, 12(7), 840. https://doi.org/10.3390/ani12070840