
The Impact of Probiotic Bacillus subtilis on Injurious Behavior in Laying Hens


Abstract
Simple Summary
Abstract
1. Introduction
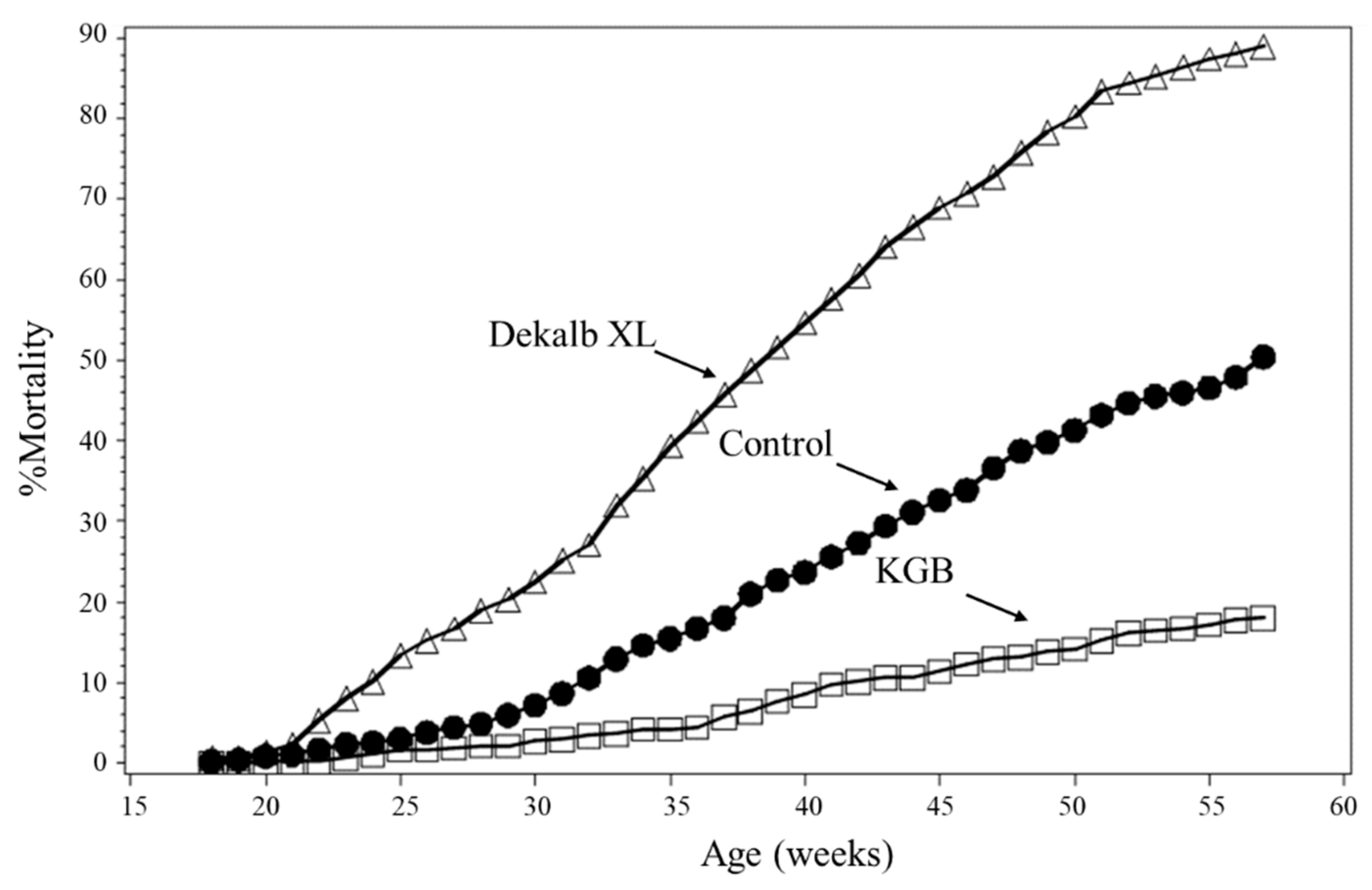
2. Production Environments and Related Stress in Commercial Laying Hens
3. Injurious Behavior in Laying Hens
3.1. Feather Pecking
3.2. Aggressive Pecking and Cannibalism
3.3. Management and Beak Trimming in Laying Hens
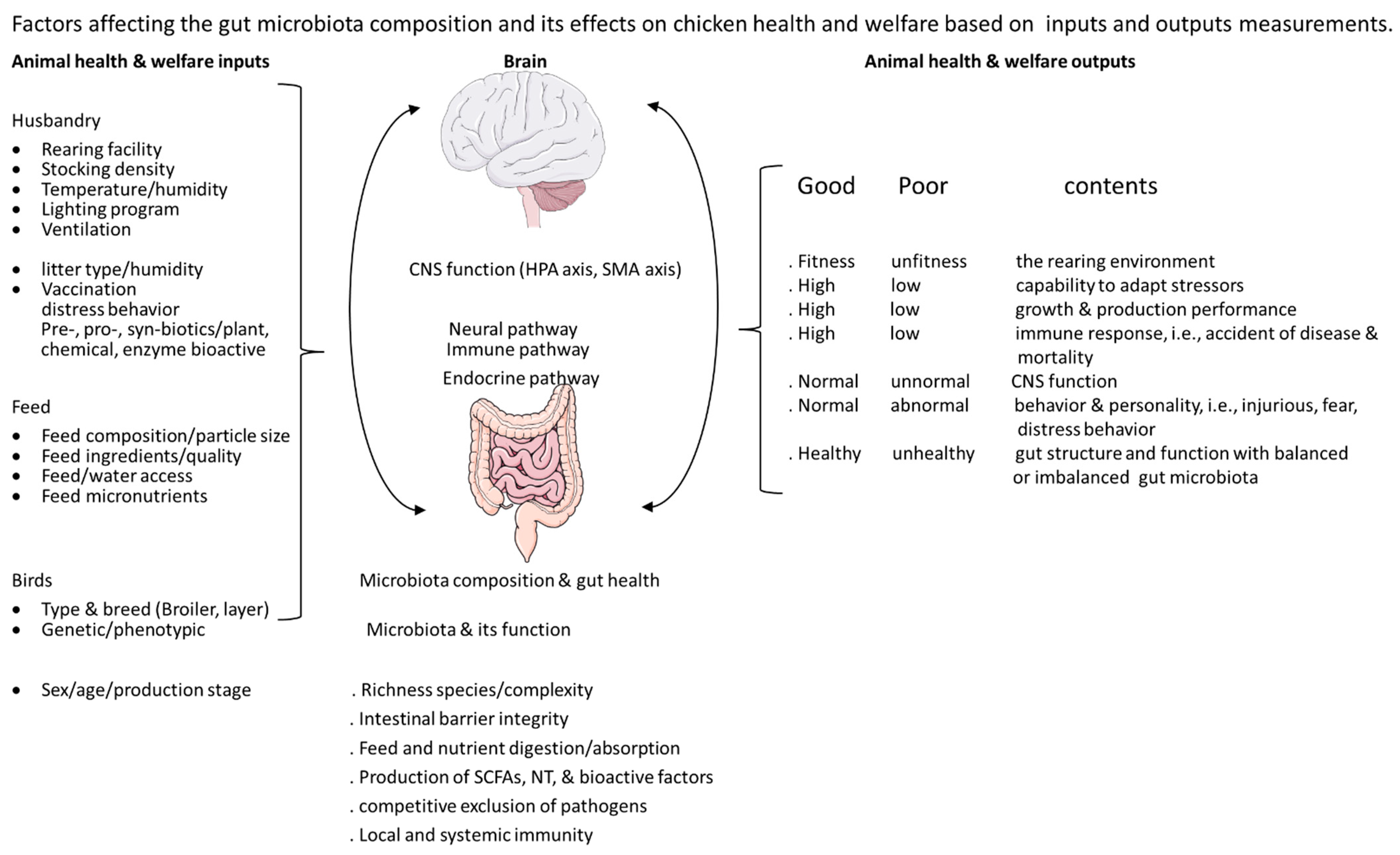
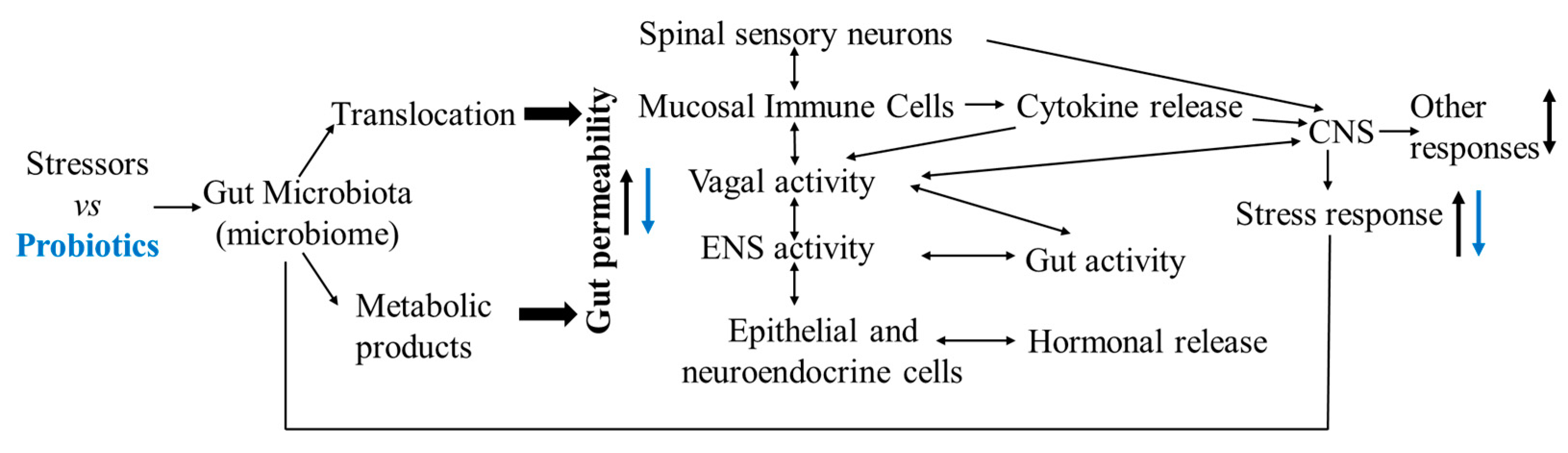
4. Gut Microbiota, Stress, Injurious Behavior, and the Microbiota–Gut–Brain Axis
5. Probiotics, Bacillus subtilis-Based Probiotics, Social Challenge-Induced Aggression
Bacillus subtilis
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faraj, J.; Takanti, V.; Tavakoli, H.R. The gut-brain axis: Literature overview and psychiatric applications. Fed. Pract. 2021, 38, 356–362. [Google Scholar] [PubMed]
- Ge, T.; Yao, X.; Zhao, H.; Yang, W.; Zou, X.; Peng, F.; Li, B.; Cui, R. Gut microbiota and neuropsychiatric disorders: Implications for neuroendocrine-immune regulation. Pharmacol. Res. 2021, 173, 105909. [Google Scholar] [CrossRef] [PubMed]
- Generoso, J.S.; Giridharan, V.V.; Lee, J.; Macedo, D.; Barichello, T. The role of the microbiota-gut-brain axis in neuropsychiatric disorders. Braz. J. Psychiatry 2021, 43, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wu, X. Brain neurotransmitter modulation by gut microbiota in anxiety and depression. Front. Cell Dev. Biol. 2021, 9, 649103. [Google Scholar] [CrossRef]
- Jacobson, A.; Yang, D.; Vella, M.; Chiu, I.M. The intestinal neuro-immune axis: Crosstalk between neurons, immune cells, and microbes. Mucosal Immunol. 2021, 14, 555–565. [Google Scholar] [CrossRef]
- Konjevod, M.; Nikolac Perkovic, M.; Sáiz, J.; Svob Strac, D.; Barbas, C.; Rojo, D. Metabolomics analysis of microbiota-gut-brain axis in neurodegenerative and psychiatric diseases. J. Pharm. Biomed. Anal. 2021, 194, 113681. [Google Scholar] [CrossRef]
- Maiuolo, J.; Gliozzi, M.; Musolino, V.; Carresi, C.; Scarano, F.; Nucera, S.; Scicchitano, M.; Oppedisano, F.; Bosco, F.; Ruga, S.; et al. The contribution of gut microbiota-brain axis in the development of brain disorders. Front. Neurosci. 2021, 15, 616883. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L., IV; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Sun, P.; Su, L.; Zhu, H.; Li, X.; Guo, Y.; Du, X.; Zhang, L.; Qin, C. Gut microbiota regulation and their implication in the development of neurodegenerative disease. Microorganisms 2021, 9, 2281. [Google Scholar] [CrossRef]
- El-Hakim, Y.; Bake, S.; Mani, K.K.; Sohrabji, F. Impact of intestinal disorders on central and peripheral nervous system diseases. Neurobiol. Dis. 2022, 165, 105627. [Google Scholar] [CrossRef]
- Matisz, C.E.; Gruber, A.J. Neuroinflammatory remodeling of the anterior cingulate cortex as a key driver of mood disorders in gastrointestinal disease and disorders. Neurosci. Biobehav. Rev. 2022, 133, 104497. [Google Scholar] [CrossRef] [PubMed]
- Yarandi, S.S.; Peterson, D.A.; Treisman, G.J.; Moran, T.H.; Pasricha, P.J. Modulatory effects of gut microbiota on the central nervous system: How gut could play a role in neuropsychiatric health and diseases. J. Neurogastroenterol. Motil. 2016, 22, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Sylvia, K.E.; Jewell, C.P.; Rendon, N.M.; St John, E.A.; Demas, G.E. Sex-specific modulation of the gut microbiome and behaviour in Siberian hamsters. Brain Behav. Immun. 2017, 60, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Goswami, A.; Wendt, F.R.; Pathak, G.A.; Tylee, D.S.; De Angelis, F.; De Lillo, A.; Polimanti, R. Role of microbes in the pathogenesis of neuropsychiatric disorders. Front. Neuroendocrinol. 2021, 62, 100917. [Google Scholar] [CrossRef]
- Tremblay, A.; Lingrand, L.; Maillard, M.; Feuz, B.; Tompkins, T.A. The effects of psychobiotics on the microbiota-gut-brain axis in early-life stress and neuropsychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 105, 110142. [Google Scholar] [CrossRef]
- Trzeciak, P.; Herbet, M. Role of the intestinal microbiome, intestinal barrier and psychobiotics in depression. Nutrients 2021, 13, 927. [Google Scholar] [CrossRef]
- Aziz, M.N.M.; Kumar, J.; Muhammad Nawawi, K.N.; Raja Ali, R.A.; Mokhtar, N.M. Irritable bowel syndrome, depression, and neurodegeneration: A bidirectional communication from gut to brain. Nutrients 2021, 13, 3061. [Google Scholar] [CrossRef]
- Person, H.; Keefer, L. Psychological comorbidity in gastrointestinal diseases: Update on the brain-gut-microbiome axis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 107, 110209. [Google Scholar] [CrossRef]
- Alagiakrishnan, K.; Halverson, T. Microbial therapeutics in neurocognitive and psychiatric disorders. J. Clin. Med. Res. 2021, 13, 439–459. [Google Scholar] [CrossRef]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaz, P. The role of microbiota-gut-brain axis in neuropsychiatric and neurological disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Heyman, M.; Ménard, S. Probiotic microorganisms: How they affect intestinal pathophysiology. Cell. Mol. Life Sci. 2002, 59, 1151–1165. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO (Food and Agriculture Organization of the United Nations and World Health Organization). Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; World Health Organization: Geneva, Switzerland, 2002; pp. 1–11. [Google Scholar]
- Anker-Ladefoged, C.; Langkamp, T.; Mueller-Alcazar, A. The potential impact of selected bacterial strains on the stress response. Healthcare 2021, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Liu, J.; Ling, Z. Short-chain fatty acids-producing probiotics: A novel source of psychobiotics. Crit. Rev. Food Sci. Nutr. 2021, 6, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Dey, G.; Mookherjee, S. Probiotics-targeting new milestones from gut health to mental health. FEMS Microbiol. Lett. 2021, 368, fnab096. [Google Scholar] [CrossRef]
- Ding, Y.; Bu, F.; Chen, T.; Shi, G.; Yuan, X.; Feng, Z.; Duan, Z.; Wang, R.; Zhang, S.; Wang, Q.; et al. A next-generation probiotic: Akkermansia muciniphila ameliorates chronic stress-induced depressive-like behaviour in mice by regulating gut microbiota and metabolites. Appl. Microbiol. Biotechnol. 2021, 105, 8411–8426. [Google Scholar] [CrossRef]
- Methiwala, H.N.; Vaidya, B.; Addanki, V.K.; Bishnoi, M.; Sharma, S.S.; Kondepudi, K.K. Gut microbiota in mental health and depression: Role of pre/pro/synbiotics in their modulation. Food Funct. 2021, 12, 4284–4314. [Google Scholar] [CrossRef]
- Wang, H.; He, S.; Xin, J.; Zhang, T.; Sun, N.; Li, L.; Ni, X.; Zeng, D.; Ma, H.; Bai, Y. Psychoactive effects of lactobacillus johnsonii against restraint stress-induced memory dysfunction in mice through modulating intestinal inflammation and permeability-a study based on the gut-brain axis hypothesis. Front. Pharmacol. 2021, 12, 662148. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Y.; Cai, R.; Li, Y.; Gu, B. A narrative review of relationship between gut microbiota and neuropsychiatric disorders: Mechanisms and clinical application of probiotics and prebiotics. Ann. Palliat. Med. 2021, 10, 2304–2313. [Google Scholar] [CrossRef]
- Meng, H.Y.H.; Mak, C.C.H.; Mak, W.Y.; Zuo, T.; Ko, H.; Chan, F.K.L. Probiotic supplementation demonstrates therapeutic potential in treating gut dysbiosis and improving neurocognitive function in age-related dementia. Eur. J. Nutr. 2022, 1–34. [Google Scholar] [CrossRef]
- Janczak, A.M.; Riber, A.B. Review of rearing-related factors affecting the welfare of laying hens. Poult. Sci. 2015, 94, 1454–1469. [Google Scholar] [CrossRef]
- Decina, C.; Berke, O.; van Staaveren, N.; Baes, C.F.; Widowski, T.M.; Harlander-Matauschek, A. A cross-sectional study on feather cover damage in Canadian laying hens in non-cage housing systems. BMC Vet. Res. 2019, 15, 435. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, L.; Jensen, P. Effects of stress during commercial hatching on growth, egg production and feather pecking in laying hens. PLoS ONE 2022, 17, e0262307. [Google Scholar] [CrossRef] [PubMed]
- Ellen, E.D.; Bijma, P. Can breeders solve mortality due to feather pecking in laying hens? Poult. Sci. 2019, 98, 3431–3442. [Google Scholar] [CrossRef] [PubMed]
- Fijn, L.B.; Staay, F.J.V.; Goerlich-Jansson, V.C.; Arndt, S.S. Importance of basic research on the causes of feather pecking in relation to welfare. Animals 2020, 10, 213. [Google Scholar] [CrossRef]
- Iffland, H.; Wellmann, R.; Preuß, S.; Tetens, J.; Bessei, W.; Piepho, H.P.; Bennewitz, J. A novel model to explain extreme feather pecking behaviour in laying hens. Behav. Genet. 2020, 50, 41–50. [Google Scholar] [CrossRef]
- De Haas, E.N.; Newberry, R.C.; Edgar, J.; Riber, A.B.; Estevez, I.; Ferrante, V.; Hernandez, C.E.; Kjaer, J.B.; Ozkan, S.; Dimitrov, I.; et al. Prenatal and Early Postnatal behavioural programming in laying hens, with possible implications for the development of injurious pecking. Front. Vet. Sci. 2021, 8, 678500. [Google Scholar] [CrossRef]
- Rieke, L.; Spindler, B.; Zylka, I.; Kemper, N.; Giersberg, M.F. Pecking behaviour in conventional layer hybrids and dual-purpose hens throughout the laying period. Front. Vet. Sci. 2021, 8, 660400. [Google Scholar] [CrossRef]
- Kuenzel, W.J. Neurobiological basis of sensory perception: Welfare implications of beak trimming. Poult. Sci. 2007, 86, 1273–1282. [Google Scholar] [CrossRef]
- Marchant-Forde, R.M.; Fahey, A.G.; Cheng, H.W. Comparative effects of infrared and one-third hot-blade trimming on beak topography, behaviour, and growth. Poult. Sci. 2008, 87, 1474–1483. [Google Scholar] [CrossRef]
- Coton, J.; Guinebretière, M.; Guesdon, V.; Chiron, G.; Mindus, C.; Laravoire, A.; Pauthier, G.; Balaine, L.; Descamps, M.; Bignon, L.; et al. Feather pecking in laying hens housed in free-range or furnished-cage systems on French farms. Br. Poult. Sci. 2019, 60, 617–627. [Google Scholar] [CrossRef]
- Schreiter, R.; Damme, K.; von Borell, E.; Vogt, I.; Klunker, M.; Freick, M. Effects of litter and additional enrichment elements on the occurrence of feather pecking in pullets and laying hens—A focused review. Vet. Med. Sci. 2019, 5, 500–507. [Google Scholar] [CrossRef] [PubMed]
- van Staaveren, N.; Ellis, J.; Baes, C.F.; Harlander-Matauschek, A. A meta-analysis on the effect of environmental enrichment on feather pecking and feather damage in laying hens. Poult. Sci. 2021, 100, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.; Hennessy, M.B.; Sachser, N. Domestication affects the structure, development and stability of biobehavioural profiles. Front. Zool. 2015, 12, S19. [Google Scholar] [CrossRef]
- Travis, J.; Reznick, D.N. Natural Selection: How Selection on behaviour interacts with selection on morphology. Curr. Biol. 2018, 28, R882–R884. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.C.; White, R.R. Breeding animals to feed people: The many roles of animal reproduction in ensuring global food security. Theriogenology 2020, 150, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R. Current and future reproductive technologies for avian species. Adv. Exp. Med. Biol. 2014, 752, 23–31. [Google Scholar]
- Webster, A.B. Welfare implications of avian osteoporosis. Poult. Sci. 2004, 83, 184–192. [Google Scholar] [CrossRef]
- Bain, M.M.; Nys, Y.; Dunn, I.C. Increasing persistency in lay and stabilising egg quality in longer laying cycles. What are the challenges? Br. Poult. Sci. 2016, 57, 330–338. [Google Scholar] [CrossRef]
- Moreira, G.C.M.; Salvian, M.; Boschiero, C.; Cesar, A.S.M.; Reecy, J.M.; Godoy, T.F.; Ledur, M.C.; Garrick, D.; Mourão, G.B.; Coutinho, L.L. Genome-wide association scan for QTL and their positional candidate genes associated with internal organ traits in chickens. BMC Genom. 2019, 20, 669. [Google Scholar] [CrossRef]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef]
- UEP (United Egg Producers). Animal Husbandry Guidelines for U.S. Egg-Laying Hens. 2017 Edition. Available online: https://uepcertified.com/wp-content/uploads/2019/09/CF-UEP-Guidelines_17-3.pdf (accessed on 20 December 2021).
- Cheng, H.W.; Muir, W.M. Mechanisms of aggression and production in chickens: Genetic variations in the functions of serotonin, catecholamine, and corticosterone. World’s Poult. Sci. J. 2007, 63, 233–254. [Google Scholar] [CrossRef]
- Veroude, K.; Zhang-James, Y.; Fernàndez-Castillo, N.; Bakker, M.J.; Cormand, B.; Faraone, S.V. Genetics of aggressive behaviour: An overview. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2016, 171, 3–43. [Google Scholar] [CrossRef] [PubMed]
- Foister, S.; Doeschl-Wilson, A.; Roehe, R.; Arnott, G.; Boyle, L.; Turner, S. Social network properties predict chronic aggression in commercial pig systems. PLoS ONE 2018, 13, e0205122. [Google Scholar] [CrossRef] [PubMed]
- Bennewitz, J.; Bögelein, S.; Stratz, P.; Rodehutscord, M.; Piepho, H.P.; Kjaer, J.B.; Bessei, W. Genetic parameters for feather pecking and aggressive behaviour in a large F2-cross of laying hens using generalized linear mixed models. Poult. Sci. 2014, 93, 810–817. [Google Scholar] [CrossRef]
- UEP (Unite Egg Producers). Facts and Stats. 2020. Available online: https://unitedegg.com/facts-stats/ (accessed on 12 January 2022).
- Blatchford, R.A.; Fulton, R.M.; Mench, J.A. The utilization of the Welfare Quality® assessment for determining laying hen condition across three housing systems. Poult. Sci. 2016, 95, 154–163. [Google Scholar] [CrossRef]
- Widmar, N.; Bir, C.; Wolf, C.; Lai, J.; Liu, Y. #Eggs: Social and online media-derived perceptions of egg-laying hen housing. Poult. Sci. 2020, 99, 5697–5706. [Google Scholar]
- Appleby, M.C.; Walker, A.W.; Nicol, C.J.; Lindberg, A.C.; Freire, R.; Hughes, B.O.; Elson, H.A. Development of furnished cages for laying hens. Br. Poult. Sci. 2002, 43, 489–500. [Google Scholar] [CrossRef]
- Campbell, D.L.M.; Hinch, G.N.; Downing, J.A.; Lee, C. Early enrichment in free-range laying hens: Effects on ranging behaviour, welfare and response to stressors. Animal 2018, 12, 575–584. [Google Scholar] [CrossRef]
- Brantsæter, M.; Nordgreen, J.; Hansen, T.B.; Muri, K.; Nødtvedt, A.; Moe, R.O.; Janczak, A.M. Problem behaviours in adult laying hens—Identifying risk factors during rearing and egg production. Poult. Sci. 2018, 97, 2–16. [Google Scholar] [CrossRef]
- Zepp, M.; Louton, H.; Erhard, M.; Schmidt, P.; Helmer, F.; Schwarzer, A. The influence of stocking density and enrichment on the occurrence of feather pecking and aggressive pecking behaviour in laying hen chicks. J. Vet. Behav. 2018, 24, 9–18. [Google Scholar] [CrossRef]
- Emous, R.A.V.; Krimpen, M.M.V. Effects of Nutritional Interventions on Feathering of Poultry—A Review; Olukosi, O.A., Olori, V.E., Helmbrecht, A., Lambton, S., French, N.A., Eds.; CAB International: Wallingford, UK, 2019; pp. 133–150. [Google Scholar]
- Hocking, P.M. Unexpected consequences of genetic selection in broilers and turkeys: Problems and solutions. Br. Poult. Sci. 2014, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, T.B.; Van Krimpen, M.M.; De Jong, I.C.; De Haas, E.N.; Kops, M.S.; Riedstra, B.J.; Nordquist, R.E.; Wagenaar, J.P.; Bestman, M.; Nicol, C.J. The prevention and control of feather pecking in laying hens: Identifying the underlying principles. Worlds Poult. Sci. J. 2013, 69, 361–373. [Google Scholar] [CrossRef]
- Frizzell, K.M.; Jendral, M.J.; Maclean, I.M.; Dixon, W.T.; Putman, C.T. Physicochemical determinants of pH in pectoralis major of three strains of laying hens housed in conventional and furnished cages. Br. Poult. Sci. 2018, 59, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.M.; Duncan, I.J.H.; Mason, G. What’s in a peck? Using fixed action pattern morphology to identify the motivational basis of abnormal feather-pecking behaviour. Anim. Behav. 2008, 76, 1035–1042. [Google Scholar] [CrossRef]
- Hughes, A.L.; Buitenhuis, A.J. Reduced variance of gene expression at numerous loci in a population of chickens selected for high feather pecking. Poult. Sci. 2010, 89, 1858–1869. [Google Scholar] [CrossRef] [PubMed]
- Savory, C.J. Feather pecking and cannibalism. World’s Poult. Sci. J. 1995, 51, 215–219. [Google Scholar] [CrossRef]
- Lambton, S.L.; Nicol, C.J.; Friel, M.; Main, D.C.; McKinstry, J.L.; Sherwin, C.M.; Walton, J.; Weeks, C.A. A bespoke management package can reduce levels of injurious pecking in loose-housed laying hen flocks. Vet. Rec. 2013, 172, 423. [Google Scholar] [CrossRef]
- Blokhuis, H.J. Feather-pecking in poultry: Its relation with ground-pecking. Appl. Anim. Behav. Sci. 1986, 16, 63–67. [Google Scholar] [CrossRef]
- Rudkin, C. Feather pecking and foraging uncorrelated—The redirection hypothesis revisited. Br. Poult. Sci. 2021, 13. [Google Scholar] [CrossRef]
- Huber-Eicher, B.; Wechsler, B. Feather pecking in domestic chicks: Its relation to dustbathing and foraging. Anim. Behav. 1997, 54, 757–768. [Google Scholar] [CrossRef]
- Lay, D.C., Jr.; Fulton, R.M.; Hester, P.Y.; Karcher, D.M.; Kjaer, J.B.; Mench, J.A.; Mullens, B.A.; Newberry, R.C.; Nicol, C.J.; O’Sullivan, N.P.; et al. Hen welfare in different housing systems. Poult. Sci. 2011, 90, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Kops, M.S.; de Haas, E.N.; Rodenburg, T.B.; Ellen, E.D.; Korte-Bouws, G.A.; Olivier, B.; Güntürkün, O.; Bolhuis, J.E.; Korte, S.M. Effects of feather pecking phenotype (severe feather peckers, victims and non-peckers) on serotonergic and dopaminergic activity in four brain areas of laying hens (Gallus gallus domesticus). Physiol. Behav. 2013, 120, 77–82. [Google Scholar] [CrossRef] [PubMed]
- García-Cabrerizo, R.; Carbia, C.; O’Riordan, K.J.; Schellekens, H.; Cryan, J.F. Microbiota-gut-brain axis as a regulator of reward processes. J. Neurochem. 2020, 157, 1495–1524. [Google Scholar] [CrossRef]
- Cronin, G.M.; Hopcroft, R.L.; Groves, P.J.; Hall, E.J.S.; Phalen, D.N.; Hemsworth, P.H. Why did severe feather pecking and cannibalism outbreaks occur? An unintended case study while investigating the effects of forage and stress on pullets during rearing. Poult. Sci. 2018, 97, 1484–1502. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Aggression, rank and power: Why hens (and other animals) do not always peck according to their strength. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2022, 377, 20200434. [Google Scholar] [CrossRef] [PubMed]
- Majolo, B.; de Bortoli Vizioli, A.; Lehmann, J. The effect of intergroup competition on intragroup affiliation in primates. Anim. Behav. 2016, 114, 13–19. [Google Scholar] [CrossRef]
- Nandy, B.; Dasgupta, P.; Halder, S.; Verma, T. Plasticity in aggression and the correlated changes in the cost of reproduction in male Drosophila melanogaster. Anim. Behav. 2016, 114, 3–9. [Google Scholar] [CrossRef]
- Strauss, E.D.; Curley, J.P.; Shizuka, D.; Hobson, E.A. The centennial of the pecking order: Current state and future prospects for the study of dominance hierarchies. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2022, 377, 20200432. [Google Scholar] [CrossRef]
- Muir, W.M. Group selection for adaptation to multiple-hen cages: Selection program and direct responses. Poult. Sci. 1996, 75, 447–458. [Google Scholar] [CrossRef]
- Sherwin, E.; Bordenstein, S.R.; Quinn, J.L.; Dinan, T.G.; Cryan, J.F. Microbiota and the social brain. Science 2019, 366, eaar2016. [Google Scholar] [CrossRef]
- Papini, M.R.; Penagos-Corzo, J.C.; Pérez-Acosta, A.M. Avian emotions: Comparative perspectives on fear and frustration. Front. Psychol. 2018, 9, 2707. [Google Scholar] [CrossRef] [PubMed]
- Duncan, I.J. Behaviour and behavioural needs. Poult. Sci. 1998, 77, 1766–1772. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.S.; Mahmud, A.; Mehmood, S.; Pasha, T.N.; Khan, M.T.; Hussain, J. Assessing behaviour in Aseel pullets under free-range, part-time free-range, and cage system during growing phase. Poult. Sci. 2018, 97, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.W.; Dillworth, G.; Singleton, P.; Chen, Y.; Muirt, W.M. Effects of group selection for productivity and longevity on blood concentrations of serotonin, catecholamines, and corticosterone of laying hens. Poult. Sci. 2001, 80, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.W.; Eicher, S.D.; Chen, Y.; Singleton, P.; Muirt, W.M. Effect of genetic selection for group productivity and longevity on immunological and hematological parameters of chickens. Poult. Sci. 2001, 80, 1079–1086. [Google Scholar] [CrossRef]
- Cheng, H.W.; Muir, W.M. The effects of genetic selection for survivability and productivity on chicken physiological homeostasis. Worlds Poult. Sci. J. 2005, 61, 383–397. [Google Scholar] [CrossRef]
- Kriegseis, I.; Bessei, W.; Meyer, B.; Zentek, J.; Würbel, H.; Harlander-Matauschek, A. Feather-pecking response of laying hens to feather and cellulose-based rations fed during rearing. Poult. Sci. 2012, 91, 1514–1521. [Google Scholar] [CrossRef]
- Appleby, M.C. Modification of laying hen cages to improve behaviour. Poult. Sci. 1998, 77, 1828–1832. [Google Scholar] [CrossRef]
- Hartcher, K.M.; Tran, K.T.; Wilkinson, S.J.; Hemsworth, P.H.; Thomson, P.C.; Cronin, G.M. The effects of environmental enrichment and beak-trimming during the rearing period on subsequent feather damage due to feather-pecking in laying hens. Poult. Sci. 2015, 94, 852–859. [Google Scholar] [CrossRef]
- Iqbal, A.; Moss, A.F. Review: Key tweaks to the chicken’s beak: The versatile use of the beak by avian species and potential approaches for improvements in poultry production. Animal 2021, 15, 100119. [Google Scholar] [CrossRef]
- Struthers, S.; Classen, H.L.; Gomis, S.; Schwean-Lardner, K. The effect of beak tissue sloughing and post-treatment beak shape on the productivity of infrared beak-treated layer pullets and hens. Poult. Sci. 2019, 98, 3637–3646. [Google Scholar] [CrossRef] [PubMed]
- Riber, A.B.; Hinrichsen, L.K. Welfare consequences of omitting beak trimming in barn layers. Front. Vet. Sci. 2017, 4, 222. [Google Scholar] [CrossRef] [PubMed]
- Muir, W.M.; Cheng, H.W. Genetic Influences on the Behaviour of Chickens Associated with Welfare and Productivity. In Genetics and the Behavior of Domestic Animals, 2nd ed.; Temple, G., Mark, J.D., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 317–359. [Google Scholar]
- Falker-Gieske, C.; Iffland, H.; Preuß, S.; Bessei, W.; Drögemüller, C.; Bennewitz, J.; Tetens, J. Meta-analyses of genome wide association studies in lines of laying hens divergently selected for feather pecking using imputed sequence level genotypes. BMC Genet. 2020, 21, 114. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.J.; Wu, X.L.; Chen, W.Q.; Li, Y.F.; Zhang, G.F.; Chao, L.M.; Zhou, J.H.; Guo, A.; Liu, C.; Guo, S.N. The gut microbiome modulates the changes in liver metabolism and in inflammatory processes in the brain of chronic unpredictable mild stress rats. Oxid. Med. Cell. Longev. 2019, 2019, 7902874. [Google Scholar] [CrossRef]
- Thomann, A.K.; Mak, J.W.Y.; Zhang, J.W.; Wuestenberg, T.; Ebert, M.P.; Sung, J.J.Y.; Bernstein, Ç.N.; Reindl, W.; Ng, S.C. Review article: Bugs, inflammation and mood-a microbiota-based approach to psychiatric symptoms in inflammatory bowel diseases. Aliment. Pharmacol. Ther. 2020, 52, 247–266. [Google Scholar] [CrossRef] [PubMed]
- Münger, E.; Montiel-Castro, A.J.; Langhans, W.; Pacheco-López, G. Reciprocal interactions between gut microbiota and host social behaviour. Front. Integr. Neurosci. 2018, 12, 21. [Google Scholar] [CrossRef]
- Kraimi, N.; Dawkins, M.; Gebhardt-Henrich, S.G.; Velge, P.; Rychlik, I.; Volf, J.; Creach, P.; Smith, A.; Colles, F.; Leterrier, C. Influence of the microbiota-gut-brain axis on behaviour and welfare in farm animals: A review. Physiol. Behav. 2019, 210, 112658. [Google Scholar] [CrossRef]
- Sarkar, A.; Harty, S.; Johnson, K.V.; Moeller, A.H.; Carmody, R.N.; Lehto, S.M.; Erdman, S.E.; Dunbar, R.I.M.; Burnet, P.W.J. The role of the microbiome in the neurobiology of social behaviour. Biol. Rev. Camb. Philos. Soc. 2020, 95, 1131–1166. [Google Scholar] [CrossRef]
- Bindari, Y.R.; Gerber, P.F. Centennial Review: Factors affecting the chicken gastrointestinal microbial composition and their association with gut health and productive performance. Poult. Sci. 2022, 101, 101612. [Google Scholar] [CrossRef]
- Bosi, A.; Banfi, D.; Bistoletti, M.; Giaroni, C.; Baj, A. Tryptophan metabolites along the microbiota-gut-brain axis: An interkingdom communication system influencing the gut in health and disease. Int. J. Tryptophan Res. 2020, 13. [Google Scholar] [CrossRef]
- Chen, S.; Luo, S.; Yan, C. Gut Microbiota Implications for Health and Welfare in Farm Animals: A Review. Animals 2021, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.A.; Yalçın, S.; Latorre, J.D.; Basiouni, S.; Attia, Y.A.; Abd El-Wahab, A.; Visscher, C.; El-Seedi, H.R.; Huber, C.; Hafez, H.M.; et al. Probiotics, Prebiotics, and Phytogenic Substances for Optimizing Gut Health in Poultry. Microorganisms 2022, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- El Sabry, M.I.; Hassan, S.S.A.; Zaki, M.M.; Stino, F.K.R. Stocking density: A clue for improving social behaviour, welfare, health indices along with productivity performances of quail (Coturnix coturnix)-a review. Trop. Anim. Health Prod. 2022, 54, 83. [Google Scholar] [CrossRef] [PubMed]
- Ricke, S.C.; Rothrock, M.J., Jr. Gastrointestinal microbiomes of broilers and layer hens in alternative production systems. Poult. Sci. 2020, 99, 660–669. [Google Scholar] [CrossRef]
- Sun, L.J.; Li, J.N.; Nie, Y.Z. Gut hormones in microbiota-gut-brain cross-talk. Chin. Med. J. 2020, 133, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Gautron, J.; Dombre, C.; Nau, F.; Feidt, C.; Guillier, L. Review: Production factors affecting the quality of chicken table eggs and egg products in Europe. Animal 2022, 16 (Suppl. 1), 100425. [Google Scholar] [CrossRef] [PubMed]
- Adedokun, S.A.; Olojede, O.C. Optimizing Gastrointestinal Integrity in Poultry: The role of nutrients and feed additives. Front. Vet. Sci. 2018, 5, 348. [Google Scholar] [CrossRef]
- Villageliu, D.N.; Lyte, M. Microbial endocrinology: Why the intersection of microbiology and neurobiology matters to poultry health. Poult. Sci. 2017, 96, 2501–2508. [Google Scholar] [CrossRef]
- Cao, C.; Chowdhury, V.S.; Cline, M.A.; Gilbert, E.R. The Microbiota-Gut-Brain Axis during Heat Stress in Chickens: A Review. Front. Physiol. 2021, 12, 752265. [Google Scholar] [CrossRef]
- Lyte, J.M.; Martinez, D.A.; Robinson, K.; Donoghue, A.M.; Daniels, K.M.; Lyte, M. A neurochemical biogeography of the broiler chicken intestinal tract. Poult. Sci. 2022, 101, 101671. [Google Scholar] [CrossRef]
- Mandal, R.K.; Jiang, T.; Wideman, R.F., Jr.; Lohrmann, T.; Kwon, Y.M. Microbiota Analysis of Chickens Raised Under Stressed Conditions. Front. Vet. Sci. 2020, 7, 482637. [Google Scholar] [CrossRef] [PubMed]
- Molina-Torres, G.; Rodriguez-Arrastia, M.; Roman, P.; Sanchez-Labraca, N.; Cardona, D. Stress and the gut microbiota-brain axis. Behav. Pharmacol. 2019, 30, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Van Goor, A.; Redweik, G.A.J.; Stromberg, Z.R.; Treadwell, C.G.; Xin, H.; Mellata, M. Microbiome and biological blood marker changes in hens at different laying stages in conventional and cage free housings. Poult. Sci. 2020, 99, 2362–2374. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, C.E.; Martin, J.A.; Manriquez, F.V.; Dinan, T.G.; Cryan, J.F.; Clarke, G. Focus on the essentials: Tryptophan metabolism and the microbiome-gut-brain axis. Curr. Opin. Pharmacol. 2019, 48, 137–145. [Google Scholar] [CrossRef]
- Kaur, A.; Chen, T.; Green, S.J.; Mutlu, E.; Martin, B.R.; Rumpagaporn, P.; Patterson, J.A.; Keshavarzian, A.; Hamaker, B.R. Physical inaccessibility of a resistant starch shifts mouse gut microbiota to butyrogenic firmicutes. Mol. Nutr. Food Res. 2019, 63, e1801012. [Google Scholar] [CrossRef]
- Gao, K.; Mu, C.-L.; Farzi, A.; Zhu, W.-Y. Tryptophan metabolism: A link between the gut microbiota and brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef]
- Hoglund, E.; Overli, O.; Winberg, S. Tryptophan metabolic pathways and brain serotonergic activity: A comparative review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of tryptophan and serotonin on mood and cognition with a possible role of the gut-brain axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef]
- Comai, S.; Bertazzo, A.; Brughera, M.; Crotti, S. Tryptophan in health and disease. Adv. Clin. Chem. 2020, 95, 165–218. [Google Scholar]
- Mindus, C.; van Staaveren, N.; Fuchs, D.; Gostner, J.M.; Kjaer, J.B.; Kunze, W.; Mian, M.F.; Shoveller, A.K.; Forsythe, P.; Harlander-Matauschek, A.L. Rhamnosus improves the immune response and tryptophan catabolism in laying hen pullets. Sci. Rep. 2021, 11, 19538. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.B.; Voilque, G.; Odle, J.; Kim, S.W. Dietary L-tryptophan supplementation with reduced large neutral amino acids enhances feed efficiency and decreases stress hormone secretion in nursery pigs under social-mixing stress. J. Nutr. 2012, 142, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Morandini, L.; Roberto Ramallo, M.; Guimaraes Moreira, R.; Hoecht, C.; Manuel Somoza, G.; Silva, A.; Pandolfi, M. Serotonergic outcome, stress and sexual steroid hormones, and growth in a South American cichlid fish fed with an L-tryptophan enriched diet. Gen. Comp. Endocrinol. 2015, 223, 27–37. [Google Scholar] [CrossRef] [PubMed]
- de Haas, E.N.; van der Eijk, J.A.J. Where in the serotonergic system does it go wrong? Unravelling the route by which the serotonergic system affects feather pecking in chickens. Neurosci. Biobehav. Rev. 2018, 95, 170–188. [Google Scholar] [CrossRef]
- Mikulska, J.; Juszczyk, G.; Gawrońska-Grzywacz, M.; Herbet, M. HPA Axis in the pathomechanism of depression and schizophrenia: New therapeutic strategies based on its participation. Brain Sci. 2021, 11, 1298. [Google Scholar] [CrossRef]
- Kulikova, E.A.; Kulikov, A.V. Tryptophan hydroxylase 2 as a therapeutic target for psychiatric disorders: Focus on animal models. Expert. Opin. Ther. Targets 2019, 23, 655–667. [Google Scholar] [CrossRef]
- Pereira, G.R.C.; Tavares, G.D.B.; de Freitas, M.C.; De Mesquita, J.F. In silico analysis of the tryptophan hydroxylase 2 (TPH2) protein variants related to psychiatric disorders. PLoS ONE 2020, 15, e0229730. [Google Scholar] [CrossRef]
- Karakula-Juchnowicz, H.; Rog, J.; Juchnowicz, D.; Łoniewski, I.; Skonieczna-Żydecka, K.; Krukow, P.; Futyma-Jedrzejewska, M.; Kaczmarczyk, M. The study evaluating the effect of probiotic supplementation on the mental status, inflammation, and intestinal barrier in major depressive disorder patients using gluten-free or gluten-containing diet (SANGUT study): A 12-week, randomized, double-blind, and placebo-controlled clinical study protocol. Nutr. J. 2019, 18, 50. [Google Scholar]
- Endres, K.; Schäfer, K.H. Influence of commensal microbiota on the enteric nervous system and its role in neurodegenerative diseases. J. Innate Immun. 2018, 10, 172–180. [Google Scholar] [CrossRef]
- Grosso, G. Nutritional Psychiatry: How Diet Affects Brain through Gut Microbiota. Nutrients 2021, 13, 1282. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Geurts, L.; Hoyles, L.; Iozzo, P.; Kraneveld, A.D.; La Fata, G.; Miani, M.; Patterson, E.; Pot, B.; Shortt, C.; et al. The microbiota-gut-brain axis: Pathways to better brain health. Perspectives on what we know, what we need to investigate and how to put knowledge into practice. Cell. Mol. Life Sci. 2022, 79, 80. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef] [PubMed]
- Knuesel, T.; Mohajeri, M.H. The role of the gut microbiota in the development and progression of major depressive and bipolar disorder. Nutrients 2021, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The microbiota-gut-brain axis: From motility to mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Kraimi, N.; Calandreau, L.; Biesse, M.; Rabot, S.; Guitton, E.; Velge, P.; Leterrier, C. Absence of Gut Microbiota Reduces Emotional Reactivity in Japanese Quails (Coturnix japonica). Front. Physiol. 2018, 9, 603. [Google Scholar] [CrossRef]
- Mignon-Grasteau, S.; Chantry-Darmon, C.; Boscher, M.Y.; Sellier, N.; Le Bihan-Duval, E.; Bertin, A. Genetic Determinism of Fearfulness, General Activity and Feeding Behaviour in Chickens and Its Relationship with Digestive Efficiency. Behav. Genet. 2017, 47, 114–124. [Google Scholar] [CrossRef]
- Meyer, B.; Zentek, J.; Harlander-Matauschek, A. Differences in intestinal microbial metabolites in laying hens with high and low levels of repetitive feather-pecking behaviour. Physiol. Behav. 2013, 110–111, 96–101. [Google Scholar] [CrossRef]
- Birkl, P.; Bharwani, A.; Kjaer, J.B.; Kunze, W.; McBride, P.; Forsythe, P.; Harlander-Matauschek, A. Differences in cecal microbiome of selected high and low feather-pecking laying hens. Poult. Sci. 2018, 97, 3009–3014. [Google Scholar] [CrossRef]
- van der Eijk, J.A.J.; de Vries, H.; Kjaer, J.B.; Naguib, M.; Kemp, B.; Smidt, H.; Rodenburg, T.B.; Lammers, A. Differences in gut microbiota composition of laying hen lines divergently selected on feather pecking. Poult. Sci. 2019, 98, 7009–7021. [Google Scholar] [CrossRef]
- van der Eijk, J.A.J.; Rodenburg, T.B.; de Vries, H.; Kjaer, J.B.; Smidt, H.; Naguib, M.; Kemp, B.; Lammers, A. Early-life microbiota transplantation affects behavioural responses, serotonin and immune characteristics in chicken lines divergently selected on feather pecking. Sci. Rep. 2020, 10, 2750. [Google Scholar] [CrossRef]
- Fu, Y. Targeting Gut Microbiota to Reduce Aggression and Improve Welfare in Poultry; InnovatED, Purdue University: West Lafayette, IN, USA, 2022; pp. 10–11. [Google Scholar]
- Yan, C.; Xiao, J.; Li, Z.; Liu, H.; Zhao, X.; Liu, J.; Chen, S.; Zhao, X. Exogenous Fecal Microbial Transplantation Alters Fearfulness, Intestinal Morphology, and Gut Microbiota in Broilers. Front. Vet. Sci. 2021, 8, 706987. [Google Scholar] [CrossRef] [PubMed]
- van Staaveren, N.; Krumma, J.; Forsythe, P.; Kjaer, J.B.; Kwon, I.Y.; Mao, Y.K.; West, C.; Kunze, W.; Harlander-Matauschek, A. Cecal motility and the impact of Lactobacillus in feather pecking laying hens. Sci. Rep. 2020, 10, 12978. [Google Scholar] [CrossRef] [PubMed]
- Borda-Molina, D.; Iffland, H.; Schmid, M.; Müller, R.; Schad, S.; Seifert, J.; Tetens, J.; Bessei, W.; Bennewitz, J.; Camarinha-Silva, A. Gut Microbial Composition and Predicted Functions Are Not Associated with Feather pecking and Antagonistic Behaviour in Laying Hens. Life 2021, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (direct-fed microbials) in poultry nutrition and their effects on nutrient utilization, growth and laying performance, and gut health: A systematic review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Krysiak, K.; Konkol, D.; Korczyński, M. Overview of the Use of Probiotics in Poultry Production. Animals 2021, 11, 1620. [Google Scholar] [CrossRef]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The Gut Microbiota of Laying Hens and Its Manipulation with Prebiotics and Probiotics To Enhance Gut Health and Food Safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef]
- Sylvia, K.E.; Demas, G.E. A gut feeling: Microbiome-brain-immune interactions modulate social and affective behaviours. Horm. Behav. 2018, 99, 41–49. [Google Scholar] [CrossRef]
- Ren, C.C.; Sylvia, K.E.; Munley, K.M.; Deyoe, J.E.; Henderson, S.G.; Vu, M.P.; Demas, G.E. Photoperiod modulates the gut microbiome and aggressive behaviour in Siberian hamsters. J. Exp. Biol. 2020, 223, jeb212548. [Google Scholar]
- Pirbaglou, M.; Katz, J.; de Souza, R.J.; Stearns, J.C.; Motamed, M.; Ritvo, P. Probiotic supplementation can positively affect anxiety and depressive symptoms: A systematic review of randomized controlled trials. Nutr. Res. 2016, 36, 889–898. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Rincel, M.; Darnaudery, M. Maternal separation in rodents: A journey from gut to brain and nutritional perspectives. Proc. Nutr. Soc. 2020, 79, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Slykerman, R.F.; Hood, F.; Wickens, K.; Thompson, J.M.D.; Barthow, C.; Murphy, R.; Kang, J.; Rowden, J.; Stone, P.; Crane, J.; et al. Effect of Lactobacillus rhamnosus HN001 in pregnancy on postpartum symptoms of depression and anxiety: A randomised double-blind placebo-controlled trial. EBioMedicine 2017, 24, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.; Rackers, H.; Kimmel, M. A role for the microbiome in mother-infant interaction and perinatal depression. Int. Rev. Psychiatry 2019, 31, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Kantak, P.A.; Bobrow, D.N.; Nyby, J.G. Obsessive-compulsive-like behaviours in house mice are attenuated by a probiotic (Lactobacillus rhamnosus GG). Behav. Pharmacol. 2014, 25, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Latalova, K.; Hajda, M.; Prasko, J. Can gut microbes play a role in mental disorders and their treatment? Psychiatr. Danub. 2017, 29, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, V.L.; Hall, M.R.B.; Hall, L.J.; Cleare, A.J.; Stone, J.M.; Young, A.H. Perturbations in gut microbiota composition in psychiatric disorders: A review and meta-analysis. JAMA Psychiatry 2021, 78, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; O’Riordan, K.J.; Lee, Y.K.; Wang, G.; Zhao, J.; Zhang, H.; Cryan, J.F.; Chen, W. Towards a psychobiotic therapy for depression: Bifidobacterium breve CCFM1025 reverses chronic stress-induced depressive symptoms and gut microbial abnormalities in mice. Neurobiol. Stress 2020, 12, 100216. [Google Scholar] [CrossRef]
- Yadav, M.; Mandeep; Shukla, P. Probiotics of diverse origin and their therapeutic applications: A review. J. Am. Coll. Nutr. 2020, 39, 469–479. [Google Scholar] [CrossRef]
- Zagórska, A.; Marcinkowska, M.; Jamrozik, M.; Wiśniowska, B.; Paśko, P. From probiotics to psychobiotics—The gut-brain axis in psychiatric disorders. Benef. Microbes 2020, 11, 717–732. [Google Scholar] [CrossRef]
- Ligezka, A.N.; Sonmez, A.I.; Corral-Frias, M.P.; Golebiowski, R.; Lynch, B.; Croarkin, P.E.; Romanowicz, M. A systematic review of microbiome changes and impact of probiotic supplementation in children and adolescents with neuropsychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 108, 110187. [Google Scholar] [CrossRef]
- Ruiz-Gonzalez, C.; Roman, P.; Rueda-Ruzafa, L.; Rodriguez-Arrastia, M.; Cardona, D. Effects of probiotics supplementation on dementia and cognitive impairment: A systematic review and meta-analysis of preclinical and clinical studies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 108, 110189. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Gupta, D.; Mehrotra, R.; Mago, P. Psychobiotics: The next-generation probiotics for the brain. Curr. Microbiol. 2021, 78, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Cryan, J.F. Microbiota-targeted interventions for mental health. Curr. Opin. Psychiatry 2022, 35, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Deidda, G.; Biazzo, M. Gut and Brain: Investigating physiological and pathological interactions between microbiota and brain to gain new therapeutic avenues for brain diseases. Front. Neurosci. 2021, 15, 753915. [Google Scholar] [CrossRef]
- Ellis, R.J.; Bruce, K.D.; Jenkins, C.; Stothard, J.R.; Ajarova, L.; Mugisha, L.; Viney, M.E. Comparison of the distal gut microbiota from people and animals in Africa. PLoS ONE 2013, 8, e54783. [Google Scholar] [CrossRef]
- Reese, A.T.; Chadaideh, K.S.; Diggins, C.E.; Schell, L.D.; Beckel, M.; Callahan, P.; Ryan, R.; Emery Thompson, M.; Carmody, R.N. Effects of domestication on the gut microbiota parallel those of human industrialization. elife 2021, 10, e60197. [Google Scholar] [CrossRef]
- Mindus, C.; van Staaveren, N.; Bharwani, A.; Fuchs, D.; Gostner, J.M.; Kjaer, J.B.; Kunze, W.; Mian, M.F.; Shoveller, A.K.; Forsythe, P.; et al. Ingestion of Lactobacillus rhamnosus modulates chronic stress-induced feather pecking in chickens. Sci. Rep. 2021, 11, 17119. [Google Scholar] [CrossRef]
- Wurtman, R.J.; Hefti, F.; Melamed, E. Precursor control of neurotransmitter synthesis. Pharmacol. Rev. 1980, 32, 315–335. [Google Scholar]
- Mindus, C.; Ellis, J.; van Staaveren, N.; Harlander-Matauschek, A. Lactobacillus-Based Probiotics Reduce the Adverse Effects of Stress in Rodents: A Meta-analysis. Front. Behav. Neurosci. 2021, 15, 642757. [Google Scholar] [CrossRef]
- Parois, S.; Calandreau, L.; Kraimi, N.; Gabriel, I.; Leterrier, C. The influence of a probiotic supplementation on memory in quail suggests a role of gut microbiota on cognitive abilities in birds. Behav. Brain Res. 2017, 331, 47–53. [Google Scholar] [CrossRef]
- Mohammed, A.; Mahmoud, M.; Murugesan, R.; Cheng, H.W. Effect of a Synbiotic Supplement on Fear Response and Memory Assessment of Broiler Chickens Subjected to Heat Stress. Animals 2021, 11, 427. [Google Scholar] [CrossRef] [PubMed]
- Rushen, J. The peck orders of chickens—How do they develop and why are they linear. Anim. Behav. 1982, 30, 1129–1137. [Google Scholar] [CrossRef]
- Wang, W.C.; Yan, F.F.; Hu, J.Y.; Amen, O.A.; Cheng, H.W. Supplementation of Bacillus subtilis-based probiotic reduces heat stress-related behaviours and inflammatory response in broiler chickens. J. Anim. Sci. 2018, 96, 1654–1666. [Google Scholar] [CrossRef]
- Abdel-Azeem, N.M. Do probiotics affect the behaviour of turkey poults? J. Vet. Med. Anim. Health 2013, 5, 144–148. [Google Scholar]
- Joerger, R.D.; Ganguly, A. Current status of the preharvest application of pro- and prebiotics to farm animals to enhance the microbial safety of animal products. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Jeżewska-Frąckowiak, J.; Seroczyńska, K.; Banaszczyk, J.; Jedrzejczak, G.; Żylicz-Stachula, A.; Skowron, P.M. The promises and risks of probiotic Bacillus species. Acta Biochim. Pol. 2018, 65, 509–519. [Google Scholar] [CrossRef]
- Campbell, D.L.M.; Dickson, E.J.; Lee, C. Application of open field, tonic immobility, and attention bias tests to hens with different ranging patterns. PeerJ 2019, 7, e8122. [Google Scholar] [CrossRef]
- Paytuví-Gallart, A.; Sanseverino, W.; Winger, A.M. Daily intake of probiotic strain Bacillus subtilis DE111 supports a healthy microbiome in children attending day-care. Benef. Microbes 2020, 11, 611–620. [Google Scholar] [CrossRef]
- Trotter, R.E.; Vazquez, A.R.; Grubb, D.S.; Freedman, K.E.; Grabos, L.E.; Jones, S.; Gentile, C.L.; Melby, C.L.; Johnson, S.A.; Weir, T.L. Bacillus subtilis DE111 intake may improve blood lipids and endothelial function in healthy adults. Benef. Microbes 2020, 11, 621–630. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Shafi, M.E.; Qattan, S.Y.A.; Batiha, G.E.; Khafaga, A.F.; Abdel-Moneim, A.E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1835–1850. [Google Scholar] [CrossRef]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yan, F.F.; Hu, J.Y.; Mohammed, A.; Cheng, H.W. Bacillus subtilis-based probiotic improves skeletal health and immunity in broiler chickens exposed to heat stress. Animals 2021, 11, 1494. [Google Scholar] [CrossRef] [PubMed]
- Neveling, D.P.; Dicks, L.M.T. Probiotics: An antibiotic replacement strategy for healthy broilers and productive rearing. Probiotics Antimicrob. Proteins 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cartman, S.T.; La Ragione, R.M.; Woodward, M.J. Bacillus subtilis spores germinate in the chicken gastrointestinal tract. Appl. Environ. Microbiol. 2008, 74, 5254–5258. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Samanya, M.; Yamauchi, K. Histological alterations of intestinal villi in chickens fed dried Bacillus subtilis var. natto. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 95–104. [Google Scholar] [CrossRef]
- Fernandez-Alarcon, M.F.; Trottier, N.; Steibel, J.P.; Lunedo, R.; Campos, D.M.B.; Santana, A.M.; Pizauro, J.M., Jr.; Furlan, R.L.; Furlan, L.R. Interference of age and supplementation of direct-fed microbial and essential oil in the activity of digestive enzymes and expression of genes related to transport and digestion of carbohydrates and proteins in the small intestine of broilers. Poult. Sci. 2017, 96, 2920–2930. [Google Scholar] [CrossRef]
- Bar Shira, E.; Friedman, A. Innate immune functions of avian intestinal epithelial cells: Response to bacterial stimuli and localization of responding cells in the developing avian digestive tract. PLoS ONE 2018, 13, e0200393. [Google Scholar] [CrossRef]
- Abudabos, A.M.; Aljumaah, M.R.; Alkhulaifi, M.M.; Alabdullatif, A.; Suliman, G.M.; Al Sulaiman, A.R. Comparative effects of Bacillus subtilis and Bacillus licheniformis on live performance, blood metabolites and intestinal features in broiler inoculated with Salmonella infection during the finisher phase. Microb. Pathog. 2019, 139, 103870. [Google Scholar] [CrossRef]
- Galagarza, O.A.; Smith, S.A.; Drahos, D.J.; Eifert, J.D.; Williams, R.C.; Kuhn, D.D. Modulation of innate immunity in Nile tilapia (Oreochromis niloticus) by dietary supplementation of Bacillus subtilis endospores. Fish Shellfish Immunol. 2018, 83, 171–179. [Google Scholar] [CrossRef]
- Lyte, M. Probiotics function mechanistically as delivery vehicles for neuroactive compounds: Microbial endocrinology in the design and use of probiotics. Bioessays 2011, 33, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.-J.; Li, Y.-T.; Chen, J.-J.J.; Chen, S.-C.; Lin, Y.S.; Kou, Y.R.; Peng, C.-W. Sexually dimorphic urethral activity in response to pharmacological activation of 5-HT1A receptors in the rat. Am. J. Physiol. Renal Physiol. 2013, 305, F1332–F1342. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gong, Q.; Liu, L.; Janowski, M. Dopaminergic and serotoninergic signaling in black fur BL6 and albino, rag2-/-, immunodeficient BL6 mice subjected to lens-induced (LIM) and form-deprivation myopia (FDM). Investig. Ophthalmol. Vis. Sci. 2018, 59, 704. [Google Scholar]
- El Aidy, S.; Dinan, T.G.; Cryan, J.F. Gut Microbiota: The conductor in the orchestra of immune-neuroendocrine communication. Clin. Ther. 2015, 37, 954–967. [Google Scholar] [CrossRef]
- Sarsero, J.P.; Merino, E.; Yanofsky, C. A Bacillus subtilis operon containing genes of unknown function senses tRNA(Trp) charging and regulates expression of the genes of tryptophan biosynthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 2656–2661. [Google Scholar] [CrossRef]
- Gollnick, P.; Babitzke, P.; Antson, A.; Yanofsky, C. Complexity in regulation of tryptophan biosynthesis in Bacillus subtilis. Ann. Rev. Genet. 2005, 39, 47–68. [Google Scholar] [CrossRef]
- Bjerre, K.; Cantor, M.D.; Norgaard, J.V.; Poulsen, H.D.; Blaabjerg, K.; Canibe, N.; Jensen, B.B.; Stuer-Lauridsen, B.; Nielsen, B.; Derkx, P.M. Development of Bacillus subtilis mutants to produce tryptophan in pigs. Biotechnol. Lett. 2017, 39, 289–295. [Google Scholar] [CrossRef]
- Porter, R.J.; Phipps, A.J.; Gallagher, P.; Scott, A.; Stevenson, P.S.; O’Brien, J.T. Effects of acute tryptophan depletion on mood and cognitive functioning in older recovered depressed subjects. Am. J. Geriatr. Psychiatry 2005, 13, 607–615. [Google Scholar] [CrossRef]
- van Veen, J.F.; van Vliet, I.M.; de Rijk, R.H.; van Pelt, J.; Mertens, B.; Fekkes, D.; Zitman, F.G. Tryptophan depletion affects the autonomic stress response in generalized social anxiety disorder. Psychoneuroendocrinology 2009, 34, 1590–1594. [Google Scholar] [CrossRef]
- Duan, K.-M.; Ma, J.-H.; Wang, S.-Y.; Huang, Z.; Zhou, Y.; Yu, H. The role of tryptophan metabolism in postpartum depression. Metab. Brain Dis. 2018, 33, 647–660. [Google Scholar] [CrossRef]
- Kaluzna-Czaplinska, J.; Gatarek, P.; Chirumbolo, S.; Chartrand, M.S.; Bjorklund, G. How important is tryptophan in human health? Crit. Rev. Food Sci. Nutr. 2019, 59, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Aune, T.M.; Pogue, S.L. Inhibition of tumor-cell growth by interferon-gamma is mediated by 2 distinct mechanisms dependent upon oxygen-tension-induction of tryptophan degradation and depletion of intracellular nicotinamide adenine-dinucleotide. J. Clin. Investig. 1989, 84, 863–875. [Google Scholar] [CrossRef]
- Waider, J.; Araragi, N.; Gutknecht, L.; Lesch, K.-P. Tryptophan hydroxylase-2 (TPH2) in disorders of cognitive control and emotion regulation: A perspective. Psychoneuroendocrinology 2011, 36, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Shabalina, A.T. Dominance rank, fear scores and reproduction in cockerels. Br. Poult. Sci. 1984, 25, 297–301. [Google Scholar] [CrossRef] [PubMed]
- D’Eath, R.B.; Keeling, L.J. Social discrimination and aggression by laying hens in large groups: From peck orders to social tolerance. Appl. Anim. Behav. Sci. 2003, 84, 197–212. [Google Scholar] [CrossRef]
- Fox, A.S.; Kalin, N.H. A Translational neuroscience approach to understanding the development of social anxiety disorder and its pathophysiology. Am. J. Psychiatry 2014, 171, 1162–1173. [Google Scholar] [CrossRef]
- Jupp, B.; Murray, J.E.; Jordan, E.R.; Xia, J.; Fluharty, M.; Shrestha, S.; Robbins, T.W.; Dalley, J.W. Social dominance in rats: Effects on cocaine self-administration, novelty reactivity and dopamine receptor binding and content in the striatum. Psychopharmacology 2016, 233, 579–589. [Google Scholar] [CrossRef]
- Larrieu, T.; Sandi, C. Stress-induced depression: Is social rank a predictive risk factor? Bioessays 2018, 40, e1800012. [Google Scholar] [CrossRef]
- Guhl, A.M.; Craig, J.V.; Mueller, C.D. Selective breeding for aggressiveness in chickens. Poult. Sci. 1960, 39, 970–980. [Google Scholar] [CrossRef]
- Dennis, R.L.; Lay, D.C., Jr.; Cheng, H.W. Effects of early serotonin programming on behaviour and central monoamine concentrations in an avian model. Behav. Brain Res. 2013, 253, 290–296. [Google Scholar] [CrossRef]
- Hollis, F.; Kabbaj, M. Social defeat as an animal model for depression. ILAR J. 2014, 55, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Coppens, C.M.; de Boer, S.F.; Buwalda, B.; Meerlo, P.; Timmermans, P.J.A. The resident-intruder paradigm: A standardized test for aggression, violence and social stress. J. Vis. Exp. 2013, 4, e4367. [Google Scholar] [CrossRef] [PubMed]
- Jager, A.; Maas, D.A.; Fricke, K.; de Vries, R.B.; Poelmans, G.; Glennon, J.C. Aggressive behaviour in transgenic animal models: A systematic review. Neurosci. Biobehav. Rev. 2018, 91, 198–217. [Google Scholar] [CrossRef]
- Masis-Calvo, M.; Schmidtner, A.K.; de Moura Oliveira, V.E.; Grossmann, C.P.; de Jong, T.R.; Neumann, I.D. Animal models of social stress: The dark side of social interactions. Stress 2018, 21, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, J.E.; Ellen, E.D.; Van Reenen, C.G.; De Groot, J.; Ten Napel, J.; Koopmanschap, R.E.; Reilingh, G.D.; Uitdehaag, K.A.; Kemp, B.; Rodenburg, T.B. Effects of genetic group selection against mortality on behaviour and peripheral serotonin in domestic laying hens with trimmed and intact beaks. Physiol. Behav. 2009, 97, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, T.E.; Brammer, G.L.; Caspi, A.; Fawcett, J.P.; Raleigh, M.; Yuwiler, A.; Silva, P. Whole blood serotonin relates to violence in an epidemiological study. Biol. Psychiatry 1998, 43, 446–457. [Google Scholar] [CrossRef]
- Hercigonja Novkovic, V.; Rudan, V.; Pivac, N.; Nedic, G.; Muck-Seler, D. Platelet serotonin concentration in children with attention-deficit/hyperactivity disorder. Neuropsychobiology 2009, 59, 17–22. [Google Scholar] [CrossRef]
- Rosado, B.; Garcia-Belenguer, S.; Palacio, J.; Chacon, G.; Villegas, A.; Alcalde, A.I. Serotonin transporter activity in platelets and canine aggression. Vet. J. 2010, 186, 104–105. [Google Scholar] [CrossRef]
- Brown, G.L.; Goodwin, F.K.; Bunney, W.E., Jr. Human aggression and suicide: Their relationship to neuropsychiatric diagnoses and serotonin metabolism. Adv. Biochem. Psychopharmacol. 1982, 34, 287–307. [Google Scholar]
- Mann, J.J.; Brent, D.A.; Arango, V. The neurobiology and genetics of suicide and attempted suicide: A focus on the serotonergic system. Neuropsychopharmacology 2001, 24, 467–477. [Google Scholar] [CrossRef]
- McDonald, M.D.; Gonzalez, A.; Sloman, K.A. Higher levels of aggression are observed in socially dominant toadfish treated with the selective serotonin reuptake inhibitor, fluoxetine. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Pietraszek, M.H.; Takada, Y.; Yan, D.; Urano, T.; Serizawa, K.; Takada, A. Relationship between Serotonergic Measures in Periphery and the Brain of Mouse. Life Sci. 1992, 51, 75–82. [Google Scholar] [CrossRef]
- Sarrias, M.J.; Martinez, E.; Celada, P.; Udina, C.; Alvarez, E.; Artigas, F. Plasma free 5HT and platelet 5HT in depression: Case-control studies and the effect of antidepressant therapy. Adv. Exp. Med. Biol. 1991, 294, 653–658. [Google Scholar] [PubMed]
- Yan, F.F.; Wang, W.C.; Cheng, H.W. Bacillus subtilis based probiotic improved bone mass and altered brain serotoninergic and dopaminergic systems in broiler chickens. J. Funct. Foods 2018, 49, 501–509. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Effects | Conclusion | Reference |
|---|---|---|---|
| H & L birds of the HFP & LFP lines | Selection caused line’s differences in intestinal microbial metabolites. | These changes could relate to lines’ differences in behaviors via the MGB axis. | [142] |
| Birds of the HFP & LFP Lines | HFP birds had ↓cecal microbial beta diversity with ↑relative abundance of Clostridiae but ↓Lactobaccillacae. | Selection causes differences in the cecal microbial profile; cecal microbiota may involve in FP behavior. | [143] |
| Genotypes: Selected HFP & LFP lines | HFP birds had ↑genera of Clostridiales, ↓Staphylococcus & Lactobacillus in LM. HFP birds had ↑diversity & evenness for both cecal MAM & LM. HFP neutral birds had ↑genera of Clostridiales, ↓Lactobacillus with ↑diversity & evenness in LM but not MAM. | Feather pecking genotype but not phenotype affects LM composition; but the correlation between FP and microbiota composition remains to be elucidated. | [144] |
| Phenotypes: Feather peckers & neutrals | |||
| HFP & LFP recipients received microbiota from the same line (HMT) during the 1st two weeks | HMT influenced immune characteristics in both lines. HMT influenced active behavior and peripheral serotonin in the LFP line. HMT without effects on gut microbiota composition, stress response, and FP. | Early MT may influence the development of FP due to its effects on FP-associated behavioral and physiological characteristics. | [145] |
| Birds of the HFP & LFP lines | HFP birds had ↓genera Lactobacillus, ↑Escherichia in ileum digesta, and ↓Faecalibacturium & Blautia in cecal digesta & mucoa. | Gut microbial composition and its functions are not associated with FP & antagonistic behavior. | [149] |
| DXL recipients received CMT from lines 63, 72 or saline orally from day 1 to day 10, then boosted once from week 3 to week 5 | HFP birds had ↑tryptophan metabolism & lysine degradation in digesta & mucosa. There were no line effects on microbial abilities. | [146] | |
| 63-CMT recipients displayed less aggressive behavior during paired aggression test with higher concentrations of serotonin. | Early-life CMT has the potential to reduce aggressive behavior through the GMB axis. |
| Birds/Treatment | Effects | Conclusion | Reference | |
|---|---|---|---|---|
| Layers | L. rhamnosus (5 × 109 CFU/mL) in-drinking water of stressed HFP birds, LFP birds, & a unselected pullets | Pecks (phenotypic and genotypic) had lower plasma TRP. L. rhamnosus caused a short-term increase in plasma TRYP and the TRP:(PHE + TYR) ratio and all subsets of T cell proportions. | A transient effect on the immune and TRP catabolism with minimal changes in behavior in pullets. | [126] |
| L. rhamnosus (5 × 109/mL) in-drinking water of chronic stressed adult HPF & LPF hens | L. rhamnosus prevented stress-induced FP. L. rhamnosus increased T cells in the spleen and cecal tonsil. L. rhamosus reduced cecal microbiota dysbiosis. | Reduces stress-induced FP; and improves hen welfare. | [173] | |
| L. rhamnosus (5 × 108 CFU/mL) orally fed peckers & non-peckers | L. rhamosus caused ↑cecal contractions and their amplitude; It positively correlated with the number of FP of peckers. | Impacted gut motility with FP phenotypic effects. | [148] | |
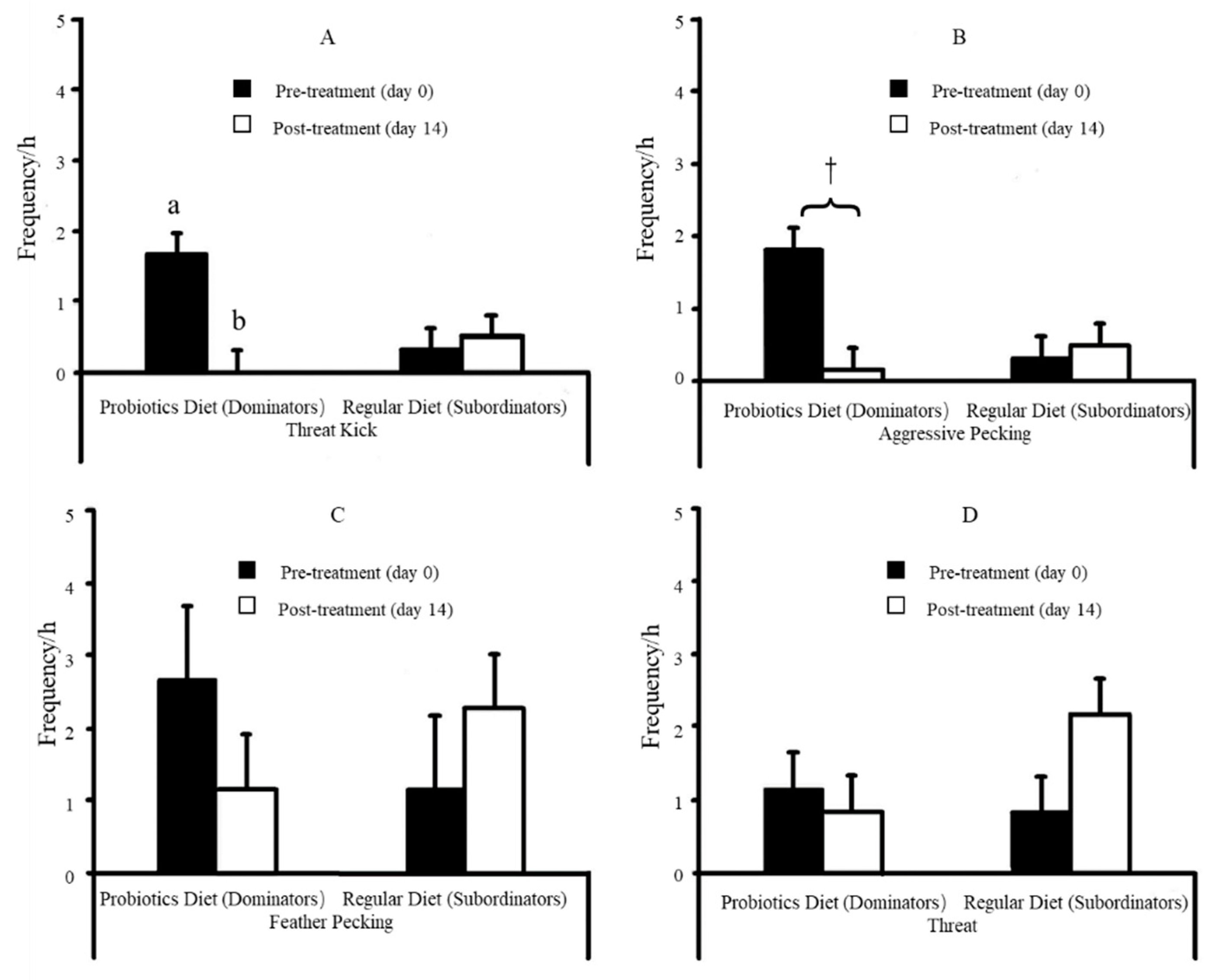
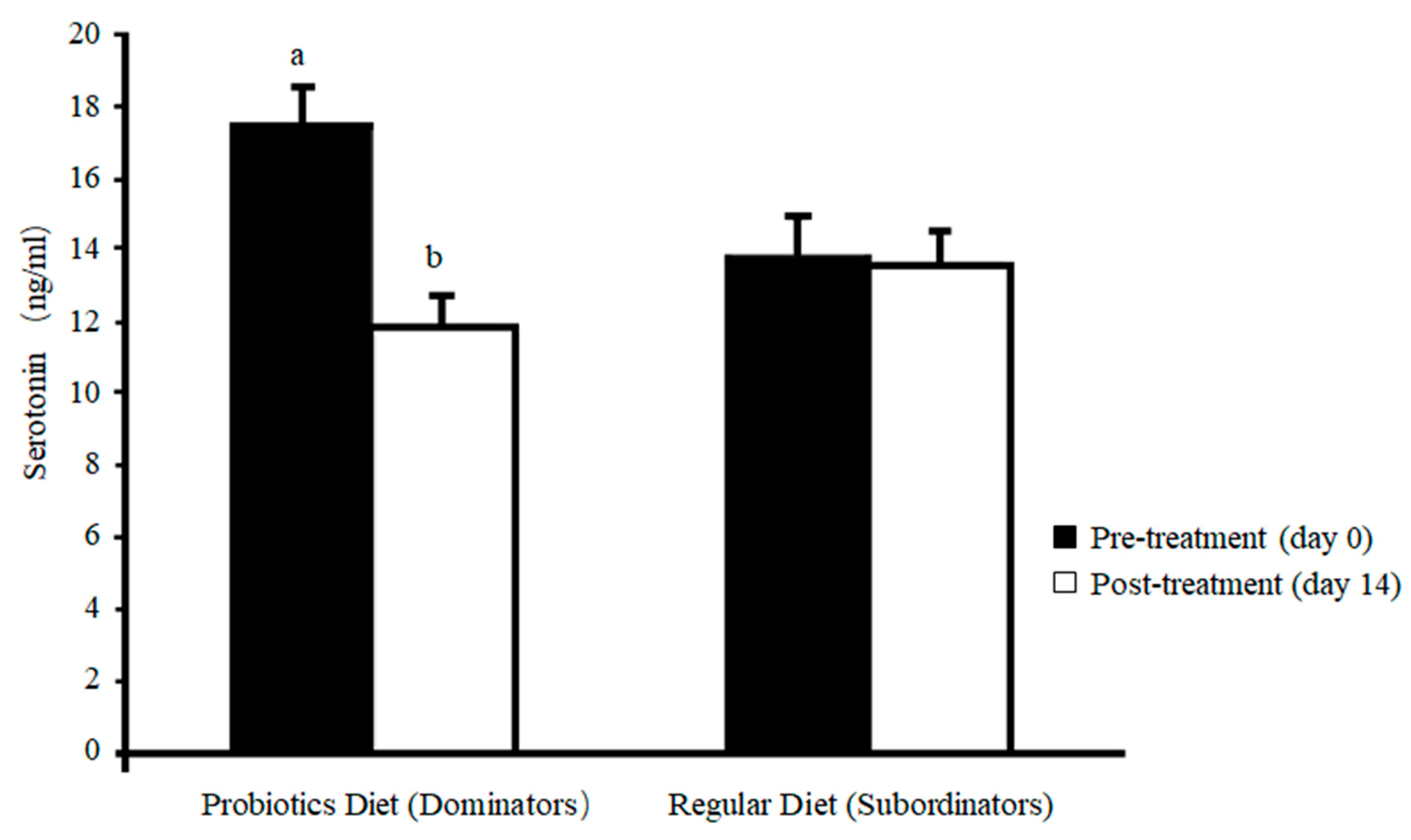
| B. subtilis (1 × 106 CFU/g) fed dominant & subordinate hens | B. subtilis caused ↓threat kick and ↓aggressive pecking during paired aggression test, and ↓plasma serotonin. B. subtilis caused ↓HS-associated behavior but ↑eating, foraging, standing, and walking. B. subtilis led to ↓hepatic IL-6, HSP70, cecal IgA & IgY but ↑hepatic IL-10. | Dietary probiotic could be a suitable strategy for controlling aggression in chickens. | [178] | |
| Broilers | B. subtilis (1 × 106 CFU/g) fed HS broilers | Reduces HS-induced inflammatory reactions via the microbiota-immune axis, while increases broilers to copy HS more effectively. | [179] | |
| * A synbiotic fed HS broilers | Snybiotic fed birds had a shorter latency to make the first vocalization, with higher vocalization rates during the isolation test and a greater number of birds reached the observer during the touch test. | The synbiotic can reduce the fear response and stress state of HS broilers. | [177] | |
| Turkeys | B. amyloliquefaciens fed turkey poults | Probiotic increased the feeding time and decreased distress call and aggressive behaviors. | Probiotics regulates behavior in turkey poults via modulation of gut microbiota. | [180] |
| Quails | P. acidilactici (2.54 × 106/g) fed STI & LTI quails | Probiotic reduces immobility duration of STI birds during TI test. | The probiotic affected host behavior and memory via the effects on gut microbiota | [176] |
| Behavior | Description |
|---|---|
| Feather Pecking | One bird pecking at feathers of another bird can be (a) gentle peck (nibbling or gentle pecking in which feathers are not removed or pulled) or (b) severe peck (vigorous pecking to feathers in which feathers are often pulled, broken, or removed). |
| Threat | One bird standing with its neck erect and hackle feathers raised in front of another bird. |
| Aggressive pecking | Forceful downward pecks directed at the head or neck of other birds |
| Threat Kick | One bird forcefully extending one or both legs such that the foot strikes another bird. |
| Lines | Corticosterone (ng/mL) | Dopamine (ng/mL) | Epinephrin (ng/mL) | Serotonin (ng/mL) | H/L Ratio 2 (×100) |
|---|---|---|---|---|---|
| KGB 1 | 1.87 + 0.19 | 0.59 + 0.08 a | 0.30 + 0.06 a | 11.8 + 0.07 a | 13.0 a |
| MBB | 1.49 + 0.21 | 2.42 + 0.76 b | 0.59 + 0.13 b | 14.3 + 0.06 b | 29.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Hu, J.-Y.; Cheng, H.-W. The Impact of Probiotic Bacillus subtilis on Injurious Behavior in Laying Hens. Animals 2022, 12, 870. https://doi.org/10.3390/ani12070870
Jiang S, Hu J-Y, Cheng H-W. The Impact of Probiotic Bacillus subtilis on Injurious Behavior in Laying Hens. Animals. 2022; 12(7):870. https://doi.org/10.3390/ani12070870
Chicago/Turabian StyleJiang, Sha, Jia-Ying Hu, and Heng-Wei Cheng. 2022. "The Impact of Probiotic Bacillus subtilis on Injurious Behavior in Laying Hens" Animals 12, no. 7: 870. https://doi.org/10.3390/ani12070870
APA StyleJiang, S., Hu, J.-Y., & Cheng, H.-W. (2022). The Impact of Probiotic Bacillus subtilis on Injurious Behavior in Laying Hens. Animals, 12(7), 870. https://doi.org/10.3390/ani12070870