
Release of DNA from Dermanyssus gallinae during the Biting Process

 ,
,  , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. Seminested PCR
2.3. Cloning and Nucleotide Sequence Determination
2.4. qPCR
2.5. Fluorescent In Situ Hybridization
3. Results
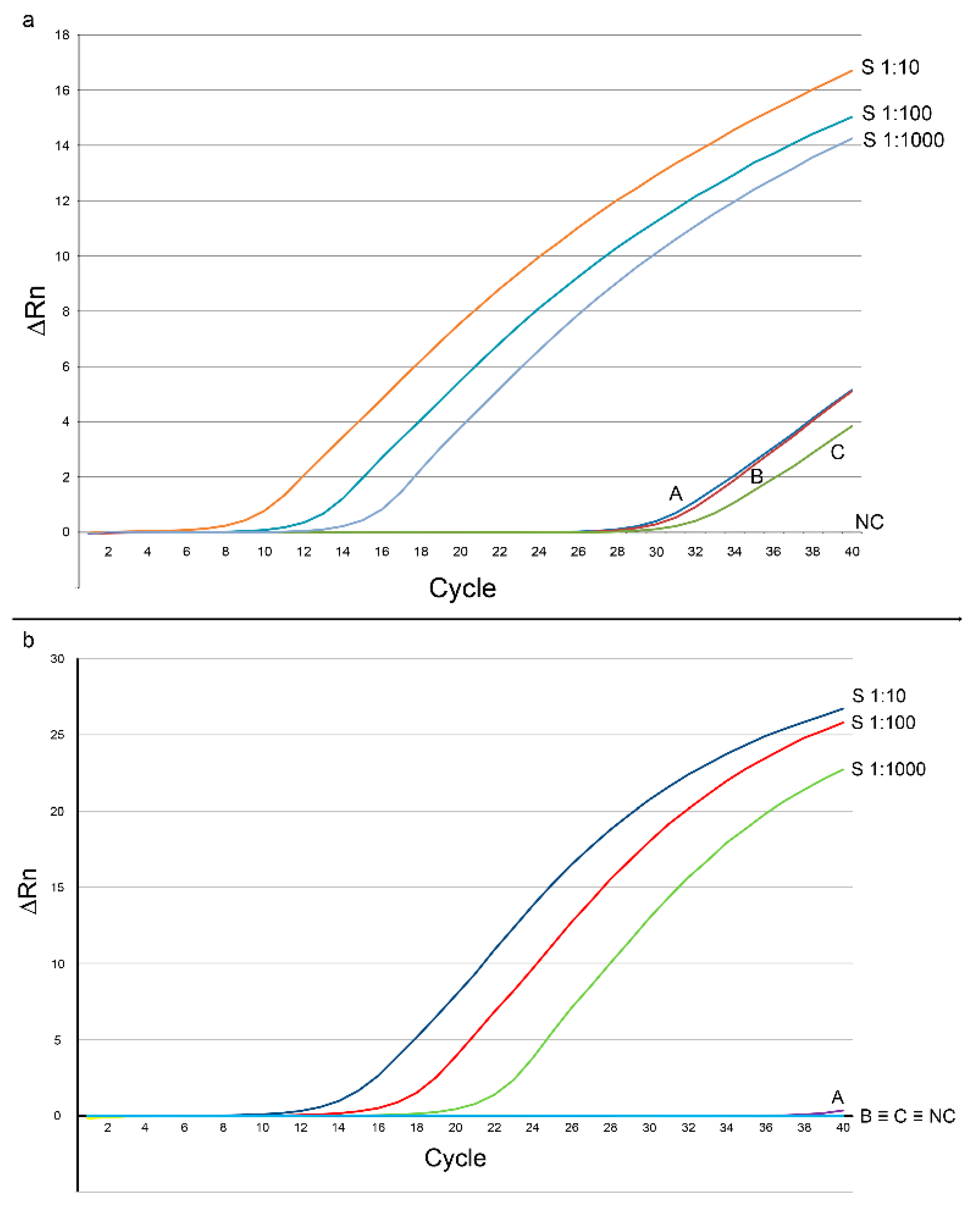
3.1. PCR Detection and Quantification of Mite DNA in Chicken Skin
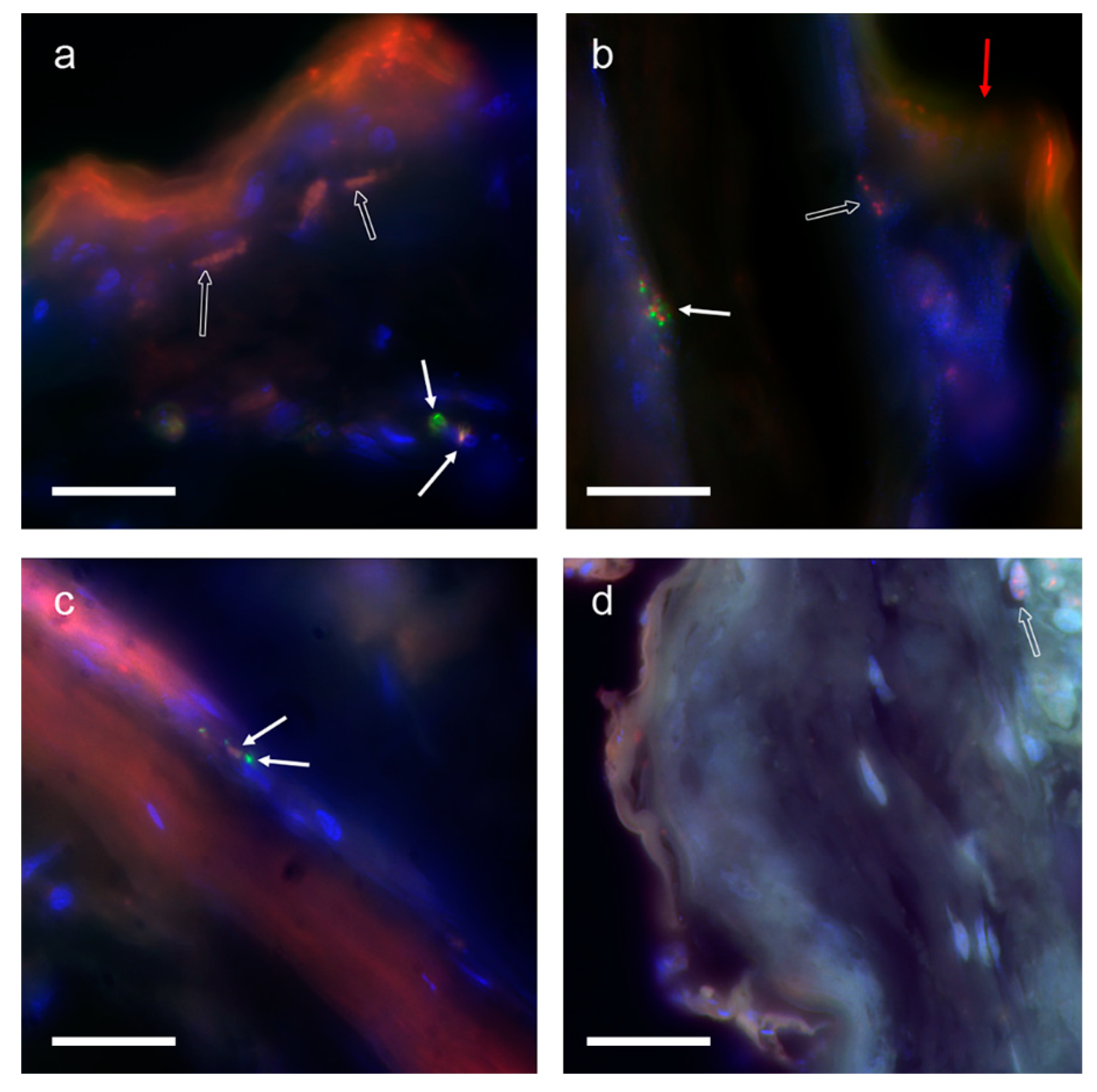
3.2. Fluorescent in Situ Hybridization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pritchard, J.; Kuster, T.; Sparagano, O.; Tomley, F. Understanding the biology and control of the poultry red mite Dermanyssus gallinae: A review. Avian Pathol. 2015, 44, 143–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axtell, R.C.; Arends, J.J. Ecology and management of arthropod pests of poultry. Annu. Rev. Entomol. 1990, 35, 101–126. [Google Scholar] [CrossRef] [PubMed]
- Sikes, R.K.; Chamberlain, R.W. Laboratory observations on three species of bird mites. J. Parasitol. 1954, 40, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Kilpinen, O.; Roepstorff, A.; Permin, A.; Nørgaard-Nielsen, G.; Lawson, L.G.; Simonsen, H.B. Influence of Dermanyssus gallinae and Ascaridia galli infections on behavior and health of laying hens (Gallus gallus domesticus). Br. Poult Sci. 2005, 46, 26–34. [Google Scholar] [CrossRef]
- Koziatek, S.; Sokół, R. Dermanyssus gallinae still poses a serious threat for the rearing of laying hens. Pol. J. Natur. Sci. 2015, 30, 451–463. [Google Scholar]
- Sigognault Flochlay, A.; Thomas, E.; Sparagano, O. Poultry red mite (Dermanyssus gallinae) infestation: A broad impact parasitological disease that still remains a significant challenge for the egg-laying industry in Europe. Parasit. Vectors 2017, 10, 357. [Google Scholar] [CrossRef]
- Cafiero, M.A.; Galante, D.; Camarda, A.; Giangaspero, A.; Sparagano, O. Why dermanyssosis should be listed as an occupational hazard. Occup. Environ. Med. 2011, 68, 628. [Google Scholar] [CrossRef]
- Cafiero, M.A.; Barlaam, A.; Camarda, A.; Radeski, M.; Mul, M.; Sparagano, O.; Giangaspero, A. Dermanyssus gallinae attacks humans. Mind the gap! Avian Pathol. 2019, 48, S22–S34. [Google Scholar] [CrossRef] [Green Version]
- Gavrilović, P.; Kecman, V.; Jovanović, M. Diagnosis of skin lesions caused by Dermanyssus gallinae in five patients. Int. J. Dermatol. 2015, 54, 207–210. [Google Scholar] [CrossRef]
- Cafiero, M.A.; Camarda, A.; Circella, E.; Galante, D.; Lomuto, M. An urban outbreak of red mite dermatitis in Italy. Int. J. Dermatol. 2009, 48, 1119–1121. [Google Scholar] [CrossRef]
- Raele, D.A.; Galante, D.; Pugliese, N.; La Salandra, G.; Lomuto, M.; Cafiero, M.A. First report of Coxiella burnetii and Borrelia burgdorferi sensu latu in poultry red mites, Dermanyssus gallinae (Mesostigmata, Acari), related to urban outbreaks of dermatitis in Italy. New Microbes New Infect. 2018, 23, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Cocciolo, G.; Circella, E.; Pugliese, N.; Lupini, C.; Mescolini, G.; Catelli, E.; Borchert-Stuhlträger, M.; Zoller, H.; Thomas, E.; Camarda, A. Evidence of vector borne transmission of Salmonella enterica enterica serovar Gallinarum and fowl typhoid disease mediated by the poultry red mite, Dermanyssus gallinae (De Geer, 1778). Parasit. Vectors 2020, 13, 513. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; Pugliese, N.; Circella, E.; Camarda, A. Association between the poultry red mite Dermanyssus gallinae and potential avian pathogenic Escherichia coli (APEC). Vet. Parasitol. 2020, 284, 109198. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; Pugliese, N.; Otranto, D.; Samarelli, R.; Circella, E.; De Virgilio, C.; Camarda, A. Dermanyssus gallinae: The long journey of the poultry red mite to become a vector. Parasit. Vectors 2022, 15, 29. [Google Scholar] [CrossRef]
- Sokoł, R.; Rotkiewicz, T. Histopathological changes of the skin in hens infested with Dermanyssus gallinae. Pol. J. Vet. Sci. 2010, 13, 385–387. [Google Scholar]
- Kowalska, M.; Kupis, B. Gamasoidosis (gamasidiosis)—Not infrequent skin reactions, frequently unrecognized. Pol. Med. Sci. Hist. Bull. 1975, 16, 391–394. [Google Scholar]
- Harrington, D.W.J.; Robinson, K.; Sparagano, O.A.E. Immune responses of the domestic fowl to Dermanyssus gallinae under laboratory conditions. Parasitol. Res. 2010, 106, 1425–1434. [Google Scholar] [CrossRef]
- Xu, X.; Wang, C.; Huang, Y.; Zhang, S.; Yu, H.; Meng, J.; Pan, B. Evaluation of the vaccine efficacy of three digestive protease antigens from Dermanyssus gallinae using an in vivo rearing system. Vaccine 2020, 38, 7842–7849. [Google Scholar] [CrossRef]
- Harrington, D.; Robinson, K.; Guy, J.; Sparagano, O. Characterization of the immunological response to Dermanyssus gallinae infestation in domestic fowl. Transbound. Emerg. Dis. 2010, 57, 107–110. [Google Scholar] [CrossRef]
- Makert, G.R.; Vorbrüggen, S.; Krautwald-Junghanns, M.E.; Voss, M.; Sohn, K.; Buschmann, T.; Ulbert, S. A method to identify protein antigens of Dermanyssus gallinae for the protection of birds from poultry mites. Parasitol. Res. 2016, 115, 2705–2713. [Google Scholar] [CrossRef]
- Bartley, K.; Huntley, J.F.; Wright, H.W.; Nath, M.; Nisbet, A.J. Assessment of cathepsin D and L-like proteinases of poultry red mite, Dermanyssus gallinae (De Geer), as potential vaccine antigens. Parasitology 2012, 139, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Bartley, K.; Wright, H.W.; Huntley, J.F.; Manson, E.D.T.; Inglis, N.F.; McLean, K.; Nath, M.; Bartley, Y.; Nisbet, A.J. Identification and evaluation of vaccine candidate antigens from the poultry red mite (Dermanyssus gallinae). Int. J. Parasitol. 2015, 45, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariizumi, T.; Murata, S.; Fujisawa, S.; Isezaki, M.; Sato, T.; Oishi, E.; Taneno, A.; Ichii, O.; Maekawa, N.; Okagawa, T.; et al. In vitro evaluation of a cysteine protease from poultry red mites, Demanyssus gallinae, as a vaccine antigen for chickens. Poult. Sci. 2022, 101, 101638. [Google Scholar] [CrossRef] [PubMed]
- Mul, M.F.; van Riel, J.W.; Meerburg, B.G.; Dicke, M.; George, D.R.; Groot Koerkamp, P.W.G. Validation of an automated mite counter for Dermanyssus gallinae in experimental laying hen cages. Exp. Appl. Acarol. 2015, 66, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Varma, M.G.R. Ticks and Mites (Acari) in Medical Insects and Arachnids; Lane, R.P., Crosskey, R.W., Eds.; University Press: Cambridge, UK, 1993; pp. 597–658. [Google Scholar]
- Baker, A.S. Mites and Ticks of Domestic Animals. An Identification Guide and Information Source; The Natural History Museum, The Stationery Office: London, UK, 2004. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Potenza, L.; Cafiero, M.A.; Camarda, A.; La Salandra, G.; Cucchiarini, L.; Dachà, M. Characterization of Dermanyssus gallinae (Acarina: Dermanyssidae) by sequence analysis of the ribosomal internal transcribed spacer regions. Vet. Res. Commun. 2009, 33, 611–618. [Google Scholar] [CrossRef]
- Lichter, P.; Tang, C.J.; Call, K.; Hermanson, G.; Evans, G.A.; Housman, D.; Ward, D.C. High-resolution mapping of human chromosome 11 by in situ hybridization with cosmid clones. Science 1990, 247, 64–69. [Google Scholar] [CrossRef]
- Capozzi, O.; Carbone, L.; Stanyon, R.R.; Marra, A.; Yang, F.; Whelan, C.W.; de Jong, P.J.; Rocchi, M.; Archidiacono, N. A comprehensive molecular cytogenetic analysis of chromosome rearrangements in gibbons. Genome Res. 2012, 22, 2520–2528. [Google Scholar] [CrossRef] [Green Version]
- Pugliese, N.; Circella, E.; Pazzani, C.; Pupillo, A.; Camarda, A. Validation of a seminested PCR approach for rapid detection of Salmonella enterica subsp. enterica serovar Gallinarum. J. Microbiol. Methods 2011, 85, 22–27. [Google Scholar] [CrossRef]
- Zhao, L. Mitochondrial DNA degradation: A quality control measure for mitochondrial genome maintenance and stress response. Enzymes 2019, 45, 311–341. [Google Scholar]
- Farkaš, R.; Ďatková, Z.; Mentelová, L.; Löw, P.; Beňová-Liszeková, D.; Beňo, M.; Sass, M.; Řehulka, P.; Řehulková, H.; Raška, O.; et al. Apocrine secretion in Drosophila salivary glands: Subcellular origin, dynamics, and identification of secretory proteins. PLoS ONE 2014, 9, e947383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shatrov, A.B.; Soldatenko, E.V. Dermal glands in freshwater mites Limnesia undulata (O.F. Müller, 1776) and Limnesia fulgida (C.L. Koch, 1836) (Acariformes, Limnesiidae). Arthropod Struct. Dev. 2016, 45, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Harrington, D.; El Din, H.M.; Guy, J.; Robinson, K.; Sparagano, O. Characterization of the immune response of domestic fowl following immunization with proteins extracted from Dermanyssus gallinae. Vet. Parasitol. 2009, 160, 285–294. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence | Target | Reference |
|---|---|---|---|
| ITSDGF | ATCCWTTCACTCACKCAGAG | Internal transcribed spacer 1 | This study |
| ITS2-R | GGGGTCGTCACACTTGATTT | Internal transcribed spacer 2 | [28] |
| ITSDGR | GTGAGTACGCGATACRAATYTAG | Internal transcribed spacer 1 | This study |
| FCOIDG | CATTAATATTAACTGCACCTGACA | Cytochrome oxidase subunit I | This study |
| RCOIDG | CCCGTGGAGTGTTGAAATTCA | Cytochrome oxidase subunit I | This study |
| COIDGSN | AAATTGYRGTAATTAAAATAGAYCATG | Cytochrome oxidase subunit I | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pugliese, N.; Raele, D.A.; Schiavone, A.; Cafiero, M.A.; Potenza, L.; Samarelli, R.; Circella, E.; Vasco, I.; Pennuzzi, G.; Camarda, A. Release of DNA from Dermanyssus gallinae during the Biting Process. Animals 2022, 12, 1084. https://doi.org/10.3390/ani12091084
Pugliese N, Raele DA, Schiavone A, Cafiero MA, Potenza L, Samarelli R, Circella E, Vasco I, Pennuzzi G, Camarda A. Release of DNA from Dermanyssus gallinae during the Biting Process. Animals. 2022; 12(9):1084. https://doi.org/10.3390/ani12091084
Chicago/Turabian StylePugliese, Nicola, Donato Antonio Raele, Antonella Schiavone, Maria Assunta Cafiero, Lucia Potenza, Rossella Samarelli, Elena Circella, Ilaria Vasco, Germana Pennuzzi, and Antonio Camarda. 2022. "Release of DNA from Dermanyssus gallinae during the Biting Process" Animals 12, no. 9: 1084. https://doi.org/10.3390/ani12091084
APA StylePugliese, N., Raele, D. A., Schiavone, A., Cafiero, M. A., Potenza, L., Samarelli, R., Circella, E., Vasco, I., Pennuzzi, G., & Camarda, A. (2022). Release of DNA from Dermanyssus gallinae during the Biting Process. Animals, 12(9), 1084. https://doi.org/10.3390/ani12091084