
Morphological Characterization and Molecular Phylogenetic Analysis of Kudoa iwatai from Large Yellow Croaker (Larimichthys crocea) as a New Host in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Procession
2.2. Morphological and Histological Examination
2.3. Ultrastructure Observation
2.4. Molecular and Phylogenetic Analysis
3. Results
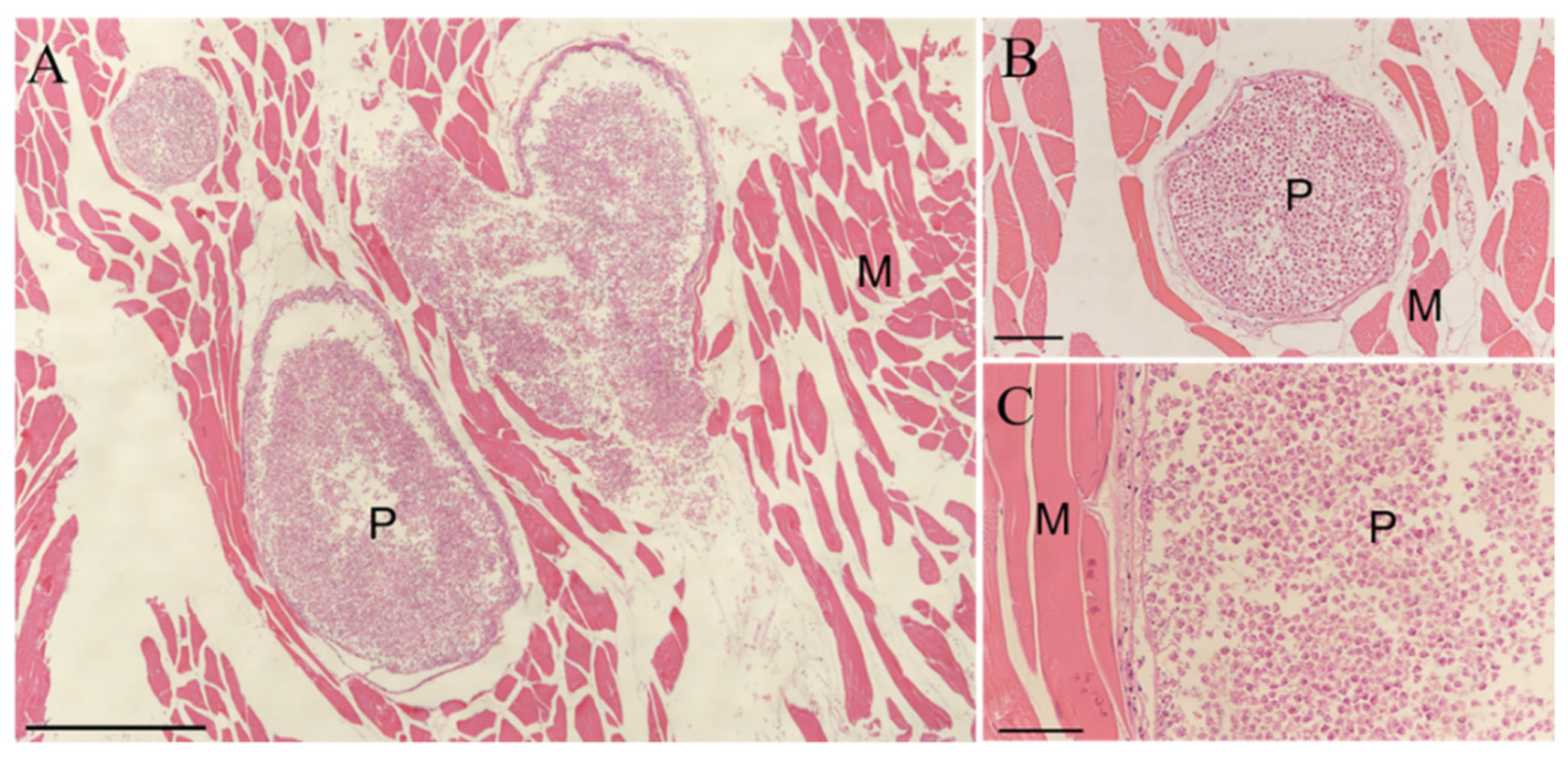
3.1. Morphological and Histopathology Examination
3.2. Ultrastructure Observation
3.3. Phylogenetic Analysis of the SSU and LSU
3.4. Phylogenetic Analysis of the Mitochondrial DNA Genes (cox-1 and rnl)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, A.; Li, W.; Wang, W.; Ye, K.; Gong, S.; Wang, Z. Analysis of DNA methylation differences in gonads of the large yellow croaker. Gene 2020, 749, 144–754. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wang, C.; Ruan, X.; Li, G.; Zhao, Y.; Wang, Y. Preservation of large yellow croaker (Pseudosciaena crocea) by Coagulin L1208, a novel bacteriocin produced by Bacillus coagulans L1208. Int. J. Food Microbiol. 2018, 266, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Su, Y.; Hong, W. Aquaculture of the Large Yellow Croaker. Aquac. China 2018, 10, 297–308. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, H.; Ke, Q.; Li, B.; Zhou, Z.; Wang, H.; Chen, B.; Pu, F.; Zhou, T.; Xu, P. Genomic selection for parasitic ciliate Cryptocaryon irritans resistance in large yellow croaker. Aquaculture 2021, 531, 735–786. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, J.; Su, Y.; Quan, C.; Zhou, H.; Yang, W. Random amplified polymorphic DNA analysis (RAPD) of Neobenedenia melleni from cultured marine fishes. Acta Oceanol. Sin. 2002, 24, 135–139. [Google Scholar] [CrossRef]
- Brazenor, A.K.; Hutson, K.S. Effects of temperature and salinity on the life cycle of Neobenedenia sp. (Monogenea: Capsalidae) infecting farmed barramundi (Lates calcarifer). Parasitol. Res. 2015, 114, 1875–1886. [Google Scholar] [CrossRef]
- Stilwell, J.; Yanong, R.P.E. Myxosporidiosis (Myxozoan Infections) in Warmwater Fish. EDIS 2017, 1–8. [Google Scholar] [CrossRef]
- Santos, W.; Silva, D.T.D.; Santos, P.; Matos, E.R.; Hamoy, I.G. New occurrence of Kudoa orbicularis parasitizing the freshwater catfish Trachelyopterus galeatus (Siluriformes: Auchenipteridae) in the Brazlian Amazon region. Rev. Bras. Parasitol. Vet. 2019, 28, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Chen, Z.; Ding, H.; Xie, J.; Wang, W.; Wang, G.; He, J.; Xu, W. Preliminary study on the histopathology and detection methods of a Myxosporea parasite that causes white-gill disease in cultured Larimichthys crocea. J. Fish. Sci. China 2019, 26, 203–213. [Google Scholar] [CrossRef]
- Shin, S.P.; Shirakashi, S.; Hamano, S.; Kato, K.; Lasso, L.T.; Yokoyama, H. Phylogenetic study of the genus Kudoa (Myxozoa: Multivalvulida) with a description of Kudoa rayformis sp. nov. from the trunk muscle of Pacific sierra Scomberomorus sierra. Mol. Phylogenet. Evol. 2016, 98, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Kasai, A.; Li, Y.C.; Mafie, E.; Sato, H. Morphological and molecular genetic characterization of two Kudoa spp., K. musculoliquefaciens, and K. pleurogrammi n. sp. (Myxosporea: Multivalvulida), causing myoliquefaction of commercial marine fish. Parasitol. Res. 2016, 115, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.S.; Hoffman, L.C.; Manley, M. A review of Kudoa-induced myoliquefaction of marine fish species in South Africa and other countries. S. Afr. J. Sci. 2013, 109, 1–5. [Google Scholar] [CrossRef]
- Whipps, C.M.; Kent, M.L. Phylogeography of the cosmopolitan marine parasite Kudoa thyrsites (Myxozoa: Myxosporea). J. Eukaryot. Microbiol. 2006, 53, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Whipps, C.M.; Grossel, G.; Adlard, R.D.; Yokoyama, H.; Bryant, M.S.; Munday, B.L.; Kent, M.L. Phylogeny of the Multivalvulidae (Myxozoa: Myxosporea) based on comparative ribosomal DNA sequence analysis. J. Parasitol. 2004, 90, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Dyková, I.; Buron, I.d.; Fiala, I.; Roumillat, W.A. Kudoa inornata sp. n. (Myxosporea: Multivalvulida) from the skeletal muscles of Cynoscion nebulosus (Teleostei: Sciaenidae). Folia Parasitol. 2009, 56, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neto, J.P.A.; Cardim, J.; Silva, D.T.D.; Hamoy, I.; Matos, E.; Abrunhosa, F. Kudoa ajurutellus n. sp. (Multivalvulida: Kudoidae), a parasite of the skeletal musculature of the Bressou sea catfish, Aspistor quadriscutis, in northeastern of the State of Para. Zootaxa 2020, 4718, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Lom, J.; Arthur, J.R. A guideline for the preparation of species descriptions in Myxosporea. J. Fish Dis. 1989, 12, 151–156. [Google Scholar] [CrossRef]
- Abdel-Baki, A.-A.S.; Al-Quraishy, S.; Omar, S.Y.A.; Mansour, L. Ultrastructure and phylogeny of Kudoa barracudai sp. nov. infecting Sphyraena putnamae in the Red Sea. Parasitol. Res. 2016, 115, 1799–1805. [Google Scholar] [CrossRef]
- Giulietti, L.; Karlsbakk, E.; Cipriani, P.; Shayo, S.D.; Storesund, J.E.; Levsen, A. Molecular Characterization of the Myoliquefactive Fish Parasite Kudoa mirabilis (Cnidaria, Kudoidae) from SE Indian Ocean and Its Phylogenetic Relationship with the Kudoa thyrsites Species Complex. Microorganisms 2020, 8, 1352. [Google Scholar] [CrossRef]
- Jeon, C.H.; Do, J.W.; Nam, U.H.; Kim, W.S.; Kim, J.H. Development of PCR method for detecting Kudoa iwatai (Myxozoa: Multivalvulida) from rock bream Oplegnathus fasciatus. Parasitol. Res. 2017, 116, 789–796. [Google Scholar] [CrossRef]
- Li, Y.C.; Inoue, K.; Zhang, J.Y.; Sato, H. Phylogenetic relationships of three Kudoa spp. with morphologically similar myxospores (K. iwatai, K. lutjanus, and K. bora), with the redescription of K. uncinata and K. petala and description of a new species (K. fujitai n. sp.) in fishes in the South China Sea. Parasitol. Res. 2020, 119, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Matsukane, Y.; Sato, H.; Tanaka, S.; Kamata, Y.; Sugita-Konishi, Y. Kudoa iwatai and two novel Kudoa spp., K. trachuri n. sp. and K. thunni n. sp. (Myxosporea: Multivalvulida), from daily consumed marine fish in western Japan. Parasitol. Res. 2011, 108, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Diamant, A.; Ucko, M.; Paperna, I.; Colorni, A.; Lipshitz, A. Kudoa iwatai (Myxosporea: Multivalvulida) in Wild and Cultured Fish in the Rea Sea: Redescription and Molecular Phylogeny. J. Parasitol. 2005, 91, 1175–1189. [Google Scholar] [CrossRef] [PubMed]
- Eiras, J.C.; Saraiva, A.l.; Cruz, C. Synopsis of the species of Kudoa Meglitsch, 1947 (Myxozoa: Myxosporea: Multivalvulida). Syst. Parasitol. 2014, 87, 153–180. [Google Scholar] [CrossRef] [PubMed]
- Egusa, S.; Shiomit, T. Two New Species of the Genus Kudoa (Myxosporea: Multivalvulida) from Marine Cultured Fishes in Japan. Fish Pathol. 1983, 18, 163–171. [Google Scholar] [CrossRef]
- Burger, M.A.; Cribb, T.H.; Adlard, R.D. Patterns of relatedness in the Kudoidae with descriptions of Kudoa chaetodoni n. sp. and K. lethrini n. sp. (Myxosporea: Multivalvulida). Parasitology 2007, 134, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Burger, M.A.A.; Adlard, R.D. Low host specificity in the Kudoidae (Myxosporea: Multivalvulida) including seventeen new host records for Kudoa thalassomi. Folia Parasitol. 2011, 58, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Moran, D.W.; Whitaker, D.J.; Kent, M.L. A review of the myxosporean genus Kudoa Meglitsch, 1947, and its impact on the international aquaculture industry and commercial fisheries. Aquaculture 1999, 172, 163–196. [Google Scholar] [CrossRef]
- Fujita, T. On a New Myxosporidia in the Muscle of the Gray-Mullet Chloromyxum bora nov. sp. Zool. Soc. Japan 1930, 42, 45–48. [Google Scholar]
- Xie, X.; Chen, Q. A New Genus and Species of Family Tetracapsulidae from Marine Fishes in South China Sea (Myxosporea: Multivalvulida). Oceanol. Limnol. Sin. 1988, 19, 431–434. [Google Scholar]
- Wang, P.C.; Huang, J.P.; Tsai, M.A.; Cheng, S.Y.; Tsai, S.S.; Chen, S.D.; Chen, S.P.; Chiu, S.H.; Liaw, L.L.; Chang, L.T.; et al. Systemic infection of Kudoa lutjanus n. sp. (Myxozoa: Myxosporea) in red snapper Lutjanus erythropterus from Taiwan. Dis. Aquat. Org. 2005, 67, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y. Taxonomic Study and Geographical Distribution of Myxosporean (Prorozoa: Myxozoa) from Some Marine Fishes of Coasts off China Waters. Ph.D. Thesis, Chongqing Normal University, Chongqing, China, 2007. [Google Scholar]
- Li, Y.C.; Inoue, K.; Tanaka, S.; Zhang, J.Y.; Sato, H. Identification of four new Kudoa spp. (Myxozoa: Myxosporea: Multivalvulida) in commercial fishes collected from South China Sea, Atlantic Ocean, and Bering Sea by integrated taxonomic approach. Parasitol. Res. 2020, 119, 2113–2128. [Google Scholar] [CrossRef] [PubMed]
- Kristmundsson, A.; Freeman, M.A. Negative effects of Kudoa islandica n. sp. (Myxosporea: Kudoidae) on aquaculture and wild fisheries in Iceland. Int. J. Parasitol. Parasites Wildl. 2014, 3, 135–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervio, D.M.L.; Khattra, J.; Devlin, R.H.; Kent, M.L.; Sakanari, J.; Yokoyama, H. Taxonomy of Kudoa species (Myxosporea), using a small-subunit ribosomal DNA sequence. Can. J. Zool. 1997, 75, 2112–2119. [Google Scholar] [CrossRef]
- Kawai, T.; Sekizuka, T.; Yahata, Y.; Kuroda, M.; Kumeda, Y.; Iijima, Y.; Kamata, Y.; Sugita-Konishi, Y.; Ohnishi, T. Identification of Kudoa septempunctata as the Causative Agent of Novel Food Poisoning Outbreaks in Japan by Consumption of Paralichthys olivaceus in Raw Fish. Clin. Infect. Dis. 2012, 54, 1046–1052. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, T.; Fujiwara, M.; Tomaru, A.; Yoshinari, T.; Sugita-Konishi, Y. Survivability of Kudoa septempunctata in human intestinal conditions. Parasitol. Res. 2016, 115, 2519–2522. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences (5′–3′) | Annealing Temperature | Length of Seq. | Reference |
|---|---|---|---|---|
| Ksp18S-F | GGATAACTGTGGTAAATCTAGAGC | 64 °C | 1300 bp | [19] |
| Ksp18S-R | GAGCAATTATTACAAGGCTCARTC | |||
| Ksp28S-R | CAGCTCCATACAAGTTTACAC | 64 °C | 1500 bp | [19] |
| Kt28S1-F | CAAGACTACCTGCTGAAC | |||
| Ki28S-F | CTATCAACCGGTGAGAACAT | 59 °C | 574 bp | [20] |
| Ki28S-R | ACGTCACACTACGCAGTTCATC | |||
| Cox1-F3 | ATGCTAATGATGGTATGGTTCCT | 52 °C | 471 bp | [21] |
| Cox1-R3 | TCTCCTCAGGAAGTATGGCT | |||
| RnL-F3 | TTCACGTGTTCAGGTTCCTT | 52 °C | 384 bp | [21] |
| RnL-R3 | ACCTTATCTTGCCGAATTCATCA |
| Species | Kudoa iwatai | Kudoa iwatai | Kudoa iwatai | Kudoa iwatai | Kudoa lutjanus | Kudoa bora | Kudoa petala |
|---|---|---|---|---|---|---|---|
| Host | Larimichthys crocea | Oplegnathus fasciatus | Acanthopagrus sclegelii | Sparus aurata | Acanthopagrus latus | Osteomugil perusii | Sillago sihama |
| Locality | Zhejiang Province, China | Korea | Japan | Israeli | Guangdong Province, China | Guangdong Province, China | Fujian Province, China |
| Reference | Present study | [20] | [22] | [23] | [21] | [21] | [21] |
| Number of examined spores | 100 | 30 | 20 | 30 | 20 | 20 | 16 |
| Thickness | 7.37–11.05 (9.38) | 6.7–8.3 (7.8) | 8.7–10.3 (10.1) | 5.0–7.0 (6.0) | 8.1–9.8 (8.9) | 8.7–10.2 (9.7) | 8.0–10.2 (9.2) |
| Sutural Thickness | 8.4–11.26 (10.01) | NA | NA | NA | 7.5–9.1 (8.2) | 8.1–9.6 (9.2) | 7.6–9.2 (8.5) |
| Width | 6.98–9.98 (8.68) | 9.8–11.8 (11.0) | NA | 6.0–7.5 (6.8) | 8.8–10.3 (9.5) | 9.6–10.4 (10.1) | 9.2–11.2 (10.4) |
| Length | 7.03–9.59 (8.15) | 6.5–8.3 (7.5) | 7.3–8.9 (8.2) | 5.0–7.0 (6.0) | 7.3–8.4 (7.9) | 8.3–9.4 (8.7) | 7.2–8.0 (7.5) |
| Polar capsule length | 2.02–5.25 (3.76) | 2.3–4.3 (3.2) | 3.7–5.0 (4.3) | 2.5–3.5 (2.8) | 3.6–4.5 (4.1) | 4.5–6.4 (5.1) | 4.0–5.2 (4.7) |
| Polar capsule width | 1.22–3.79 (2.37) | 1.3–2.5 (1.8) | 1.5–2.1 (2.2) | 1.5–2.0 (1.5) | 1.4–1.7 (1.6) | 1.9–2.5 (2.0) | 1.3–1.8 (1.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-B.; He, J.; Ma, R.-R.; Sun, F.-Y.; Wu, W.-X.; Luo, H.-M.; Bai, L.-H.; Qian, D. Morphological Characterization and Molecular Phylogenetic Analysis of Kudoa iwatai from Large Yellow Croaker (Larimichthys crocea) as a New Host in China. Animals 2022, 12, 1145. https://doi.org/10.3390/ani12091145
Li X-B, He J, Ma R-R, Sun F-Y, Wu W-X, Luo H-M, Bai L-H, Qian D. Morphological Characterization and Molecular Phylogenetic Analysis of Kudoa iwatai from Large Yellow Croaker (Larimichthys crocea) as a New Host in China. Animals. 2022; 12(9):1145. https://doi.org/10.3390/ani12091145
Chicago/Turabian StyleLi, Xiao-Bing, Jie He, Rong-Rong Ma, Fu-Ying Sun, Wen-Xin Wu, Hua-Ming Luo, Lu-Huai Bai, and Dong Qian. 2022. "Morphological Characterization and Molecular Phylogenetic Analysis of Kudoa iwatai from Large Yellow Croaker (Larimichthys crocea) as a New Host in China" Animals 12, no. 9: 1145. https://doi.org/10.3390/ani12091145
APA StyleLi, X.-B., He, J., Ma, R.-R., Sun, F.-Y., Wu, W.-X., Luo, H.-M., Bai, L.-H., & Qian, D. (2022). Morphological Characterization and Molecular Phylogenetic Analysis of Kudoa iwatai from Large Yellow Croaker (Larimichthys crocea) as a New Host in China. Animals, 12(9), 1145. https://doi.org/10.3390/ani12091145