
Physiological Changes in Chicken Embryos Inoculated with Drugs and Viruses Highlight the Need for More Standardization of this Animal Model

 ,
, 

 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Challenge of CE with Gammacoronavirus
2.2. Inoculation with DX and FG
2.2.1. Drug Dosages: Pilot Test
2.2.2. Drug Tests in CE
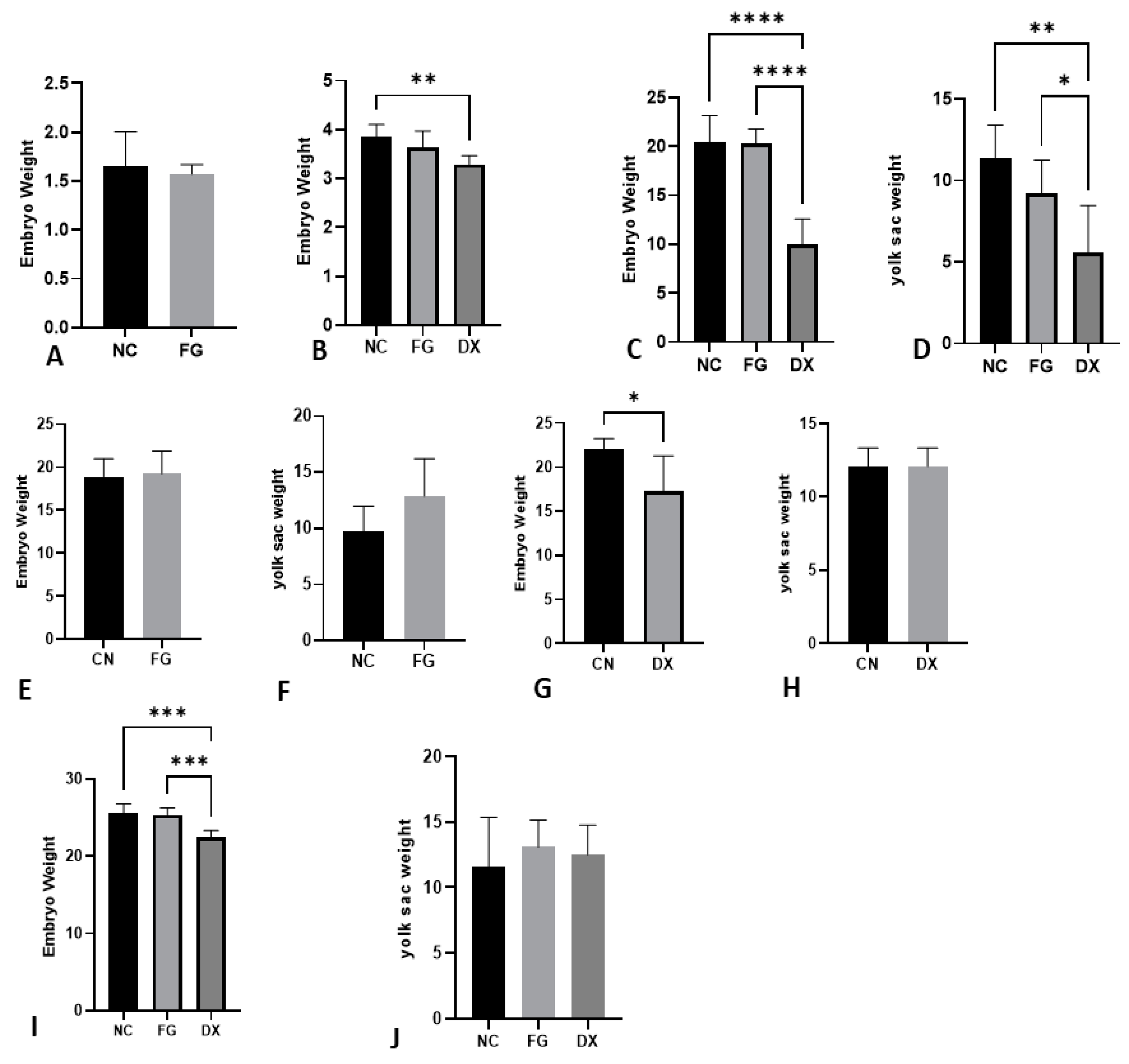
2.3. Weight of the CE and Annexes
2.4. Macroscopic Evaluations
2.5. Blood Cell Count, Hematocrit, Hemoglobin and Erythrocyte Indices
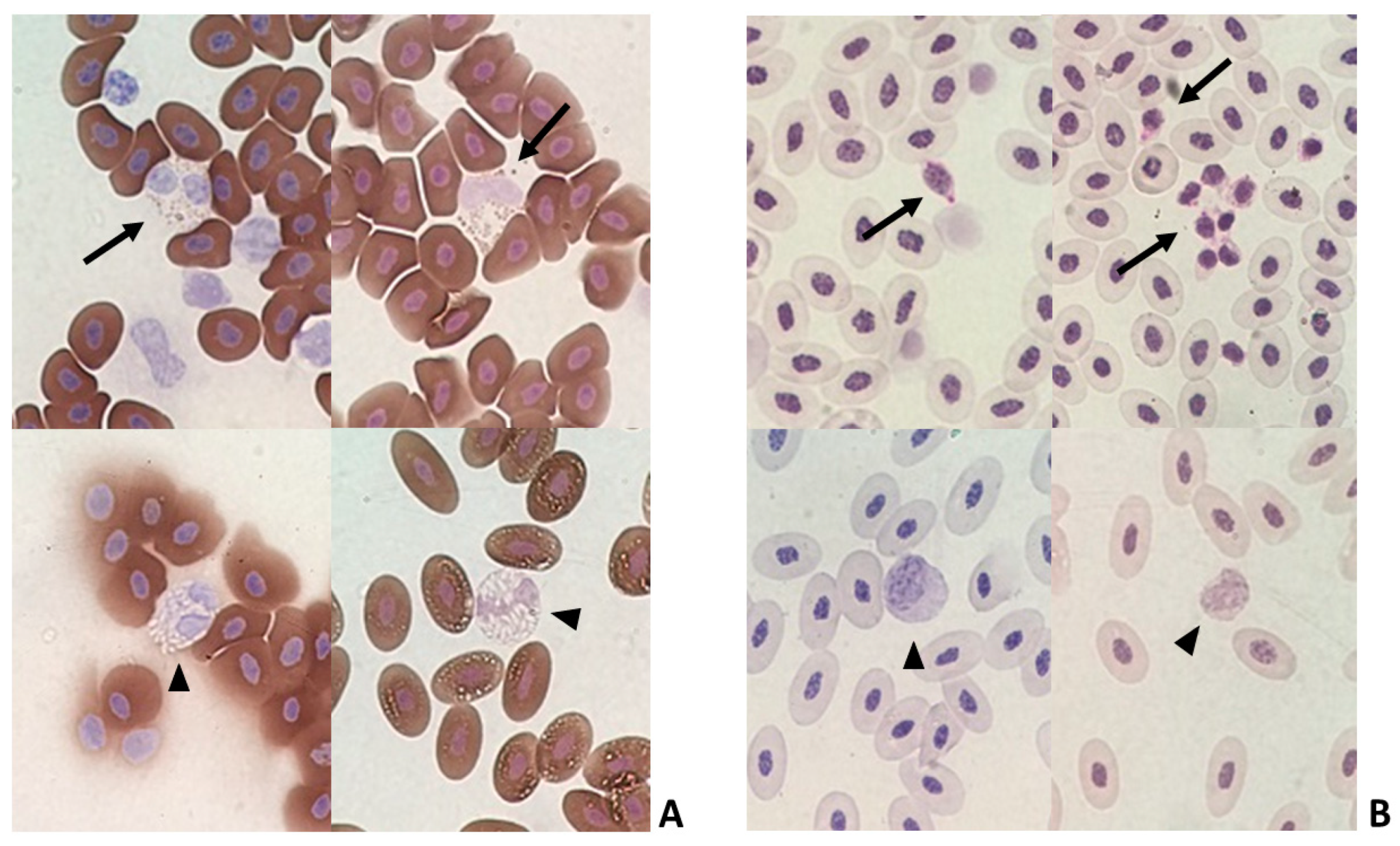
2.6. Characterisation of Blood Cells
2.7. Biochemical Analyses of the Serum and AF
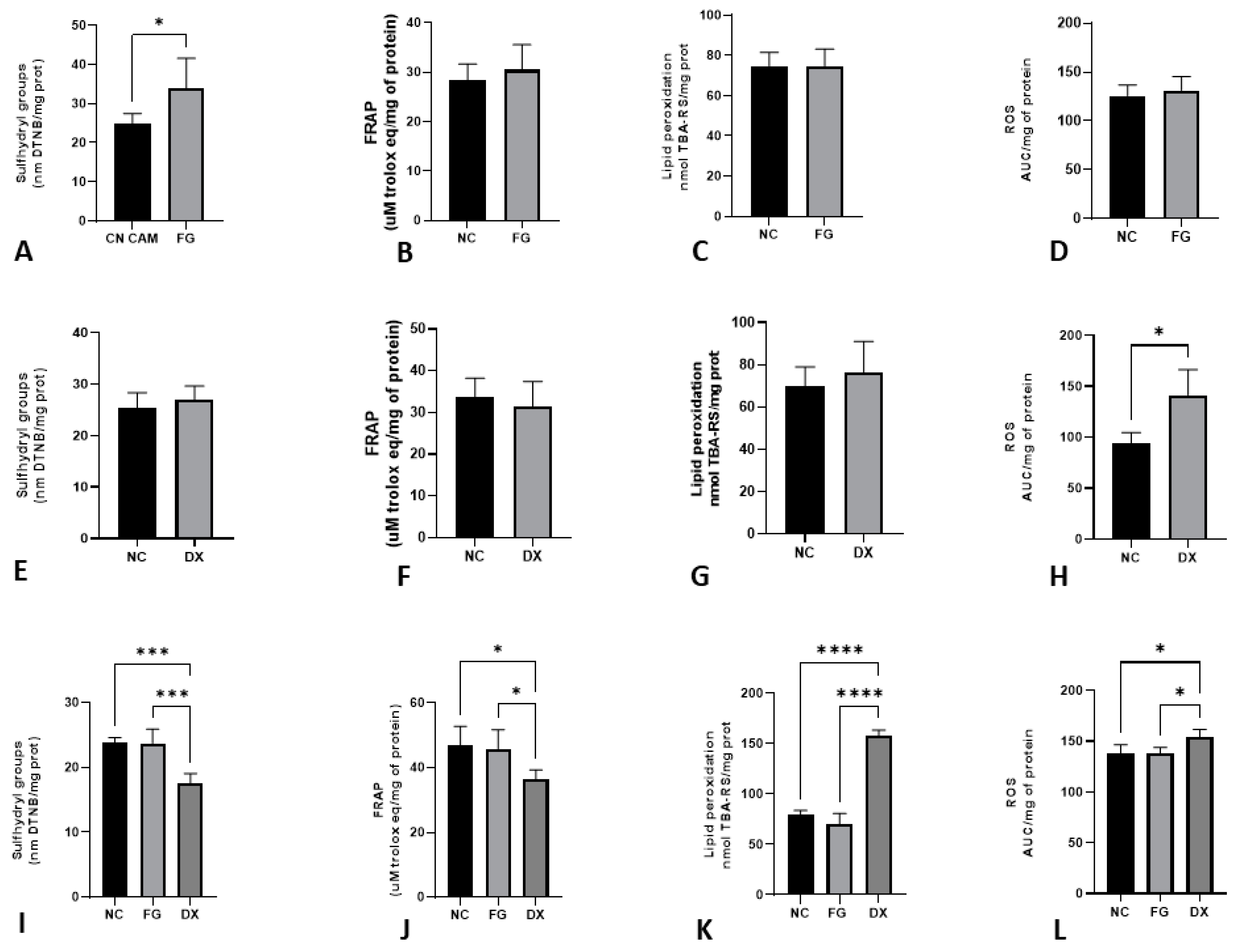
2.8. Oxidative Stress
2.8.1. Reactive Oxygen Species (ROS)
2.8.2. Lipid Peroxidation
2.8.3. Sulfhydryl Group
2.8.4. Total Antioxidant Capacity
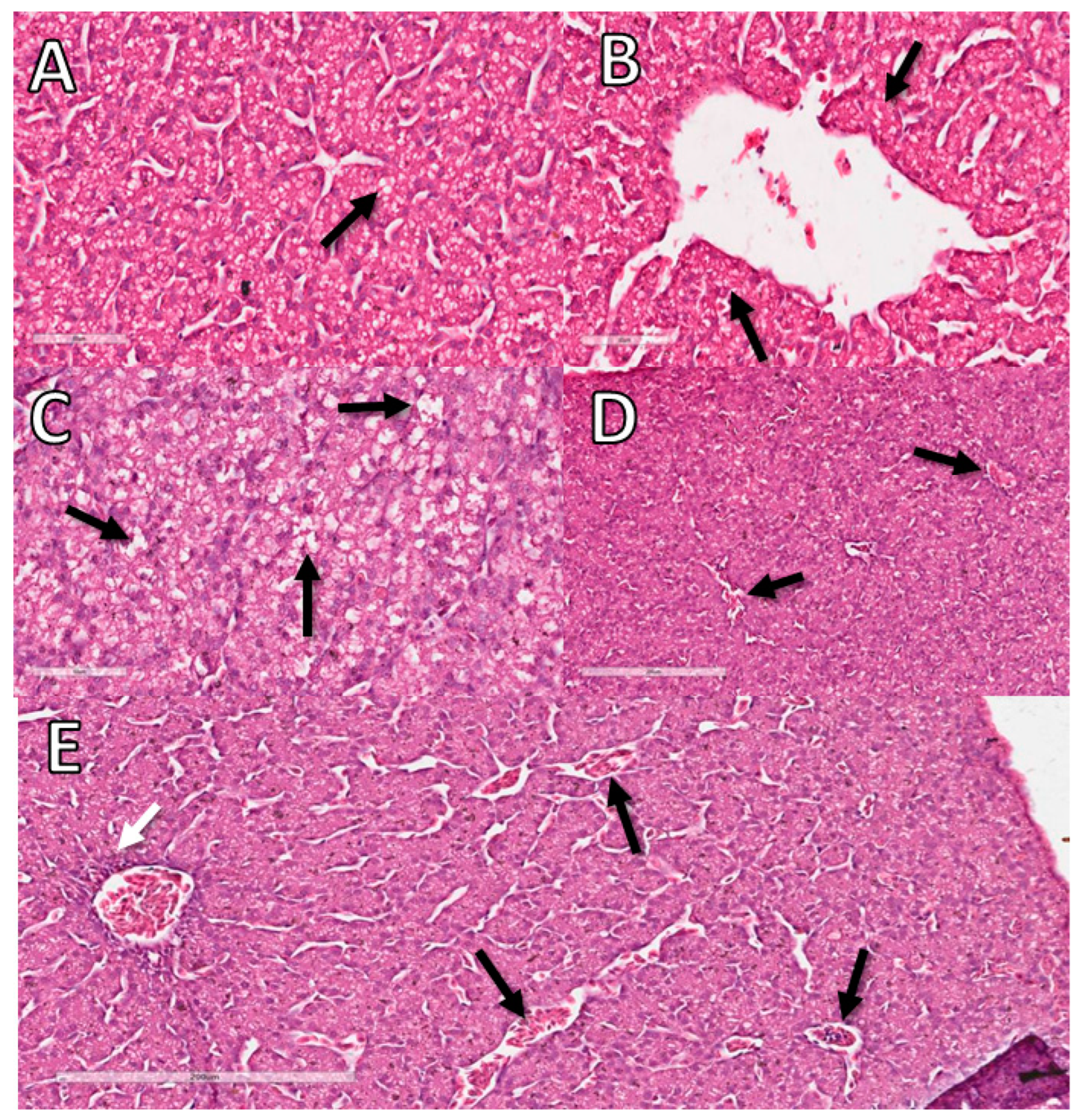
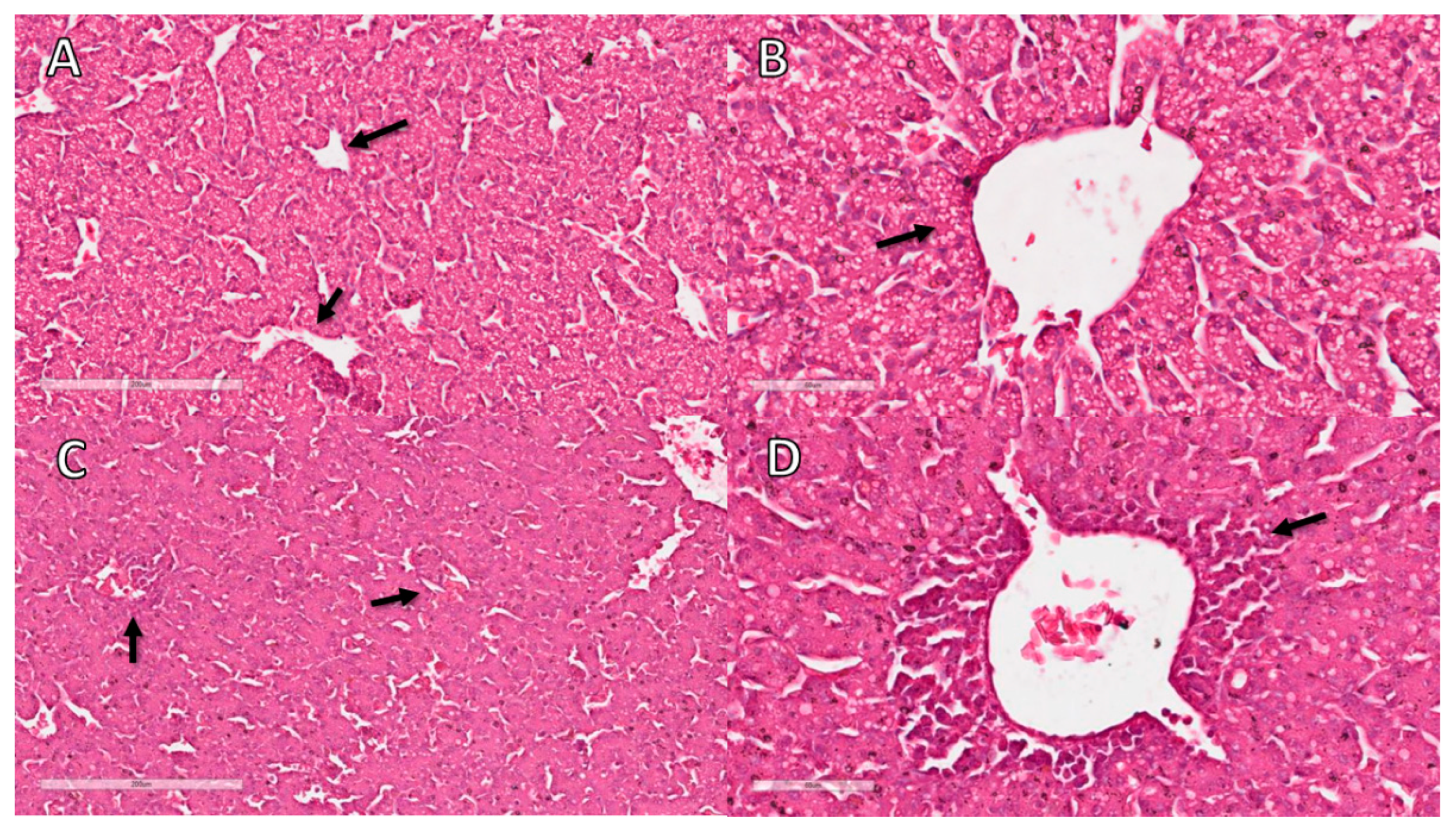
2.9. Histopathology
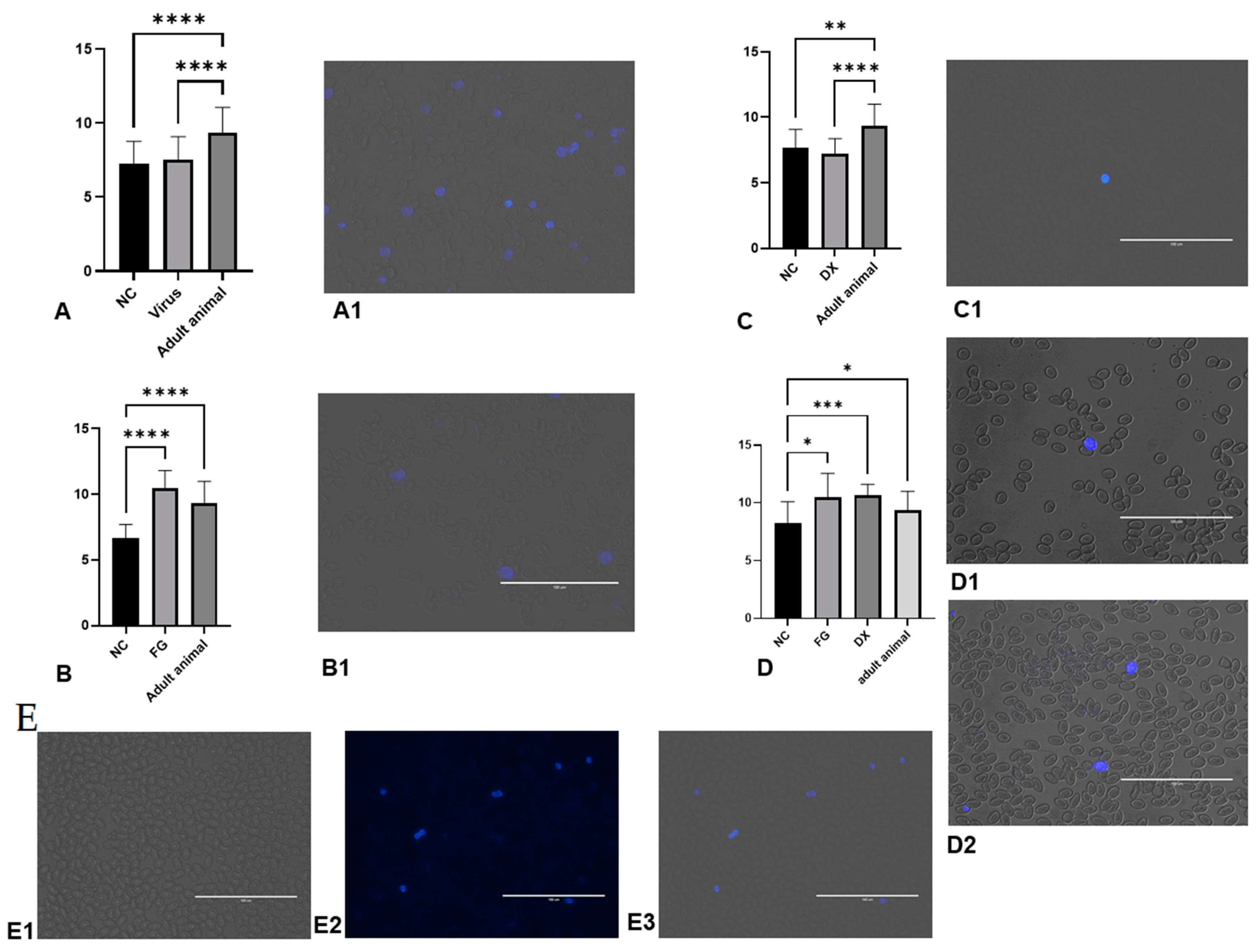
2.10. Angiogenesis
2.11. Statistical Analysis
3. Results
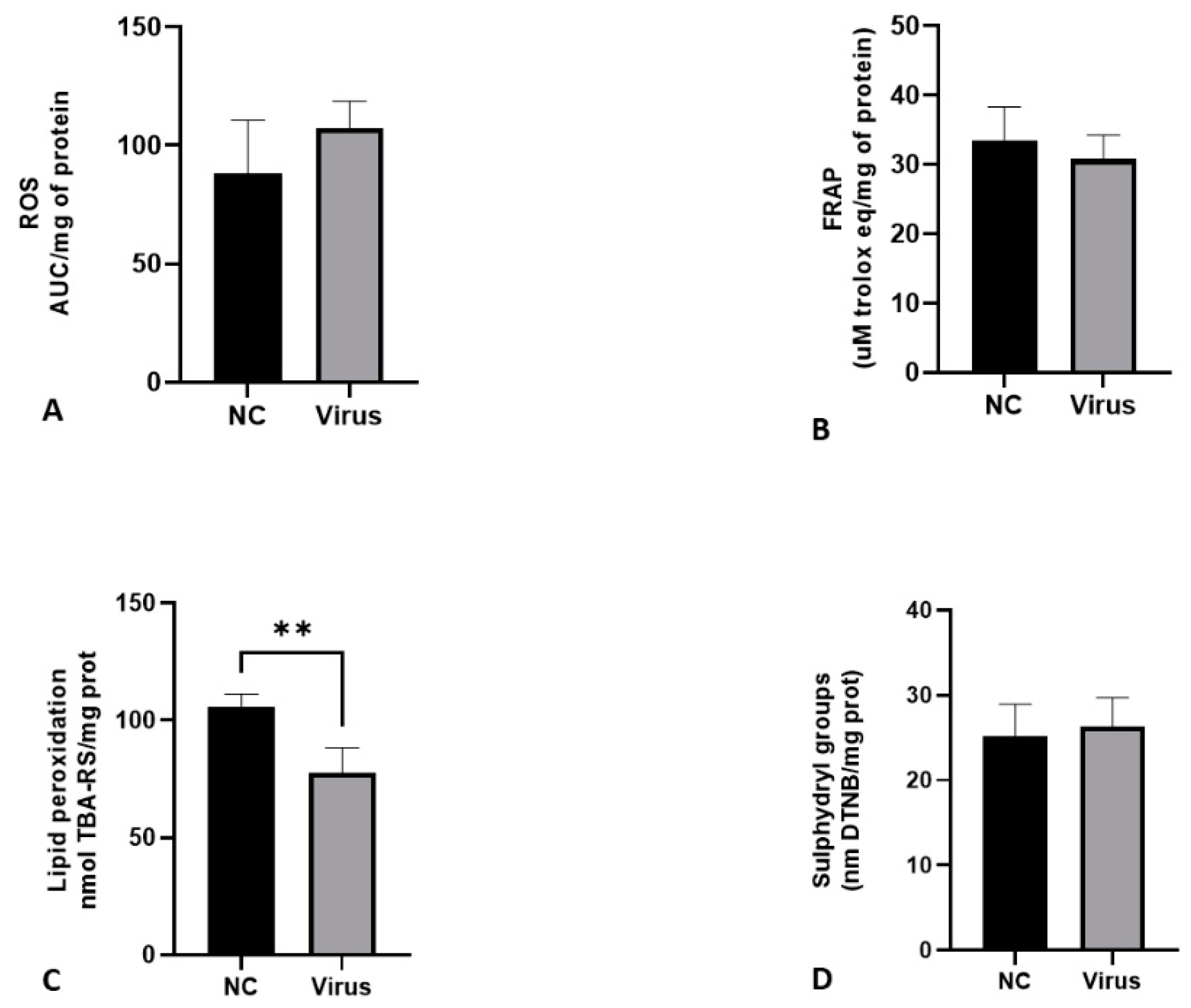
3.1. The Virus Can Change Blood Cell Count, Calcium and Lipid Peroxidation in the Liver
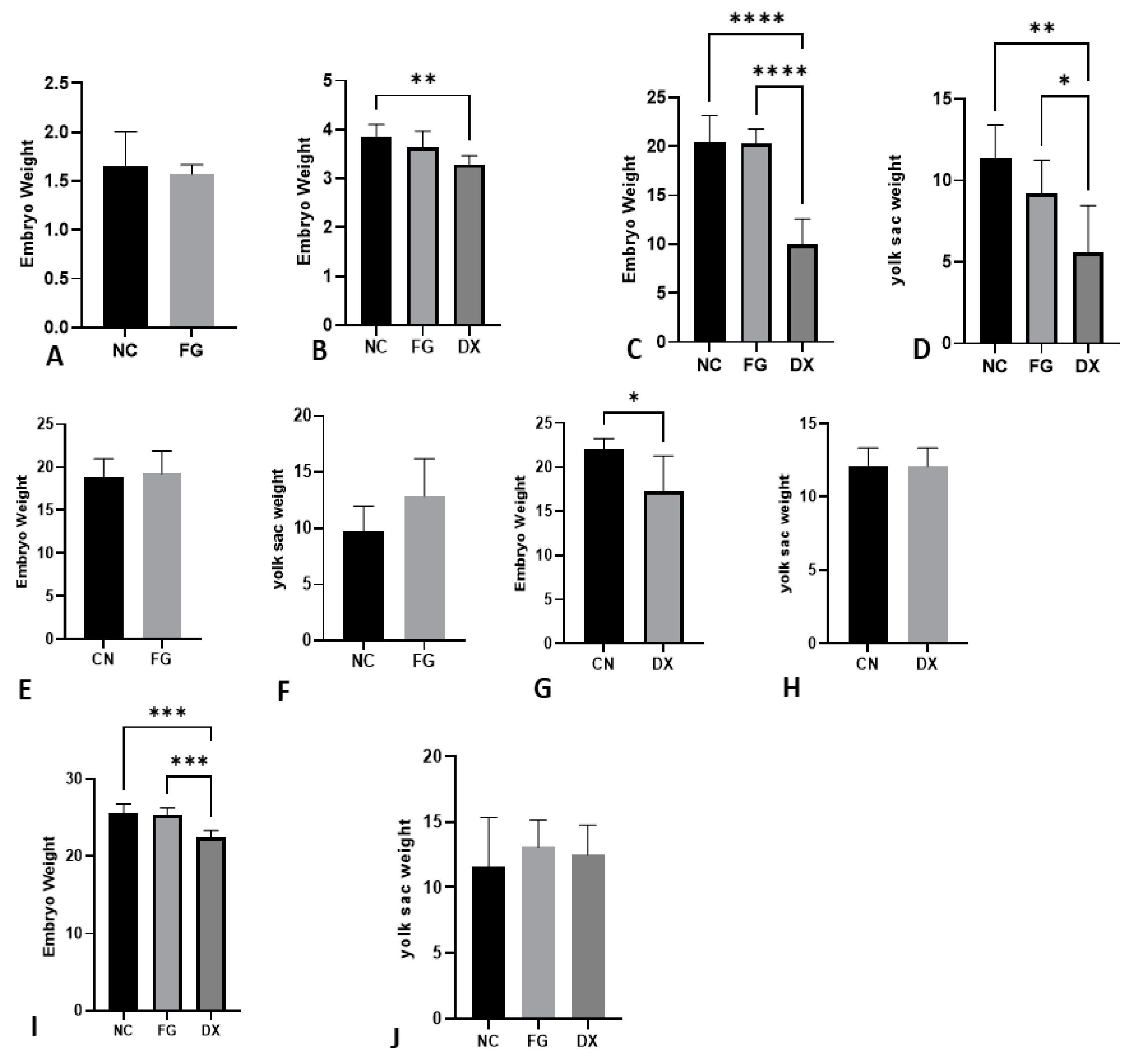
3.2. The Age of Inoculation and Type of Drug Result in Different Effects on Injury, Mortality or Embryo Viability
3.3. The Weight of CE Can Be a Drug Analysis Tool Depending on Age
3.4. The Blood Cell Count Can Change According to the Age of Inoculation
3.5. Not All Metabolites, Enzymes and/or Minerals Change Even in Liver-Damaged Embryos
3.6. The Amount of Metabolites, Enzymes and Minerals in Serum and Allantois Are Not Always Identical
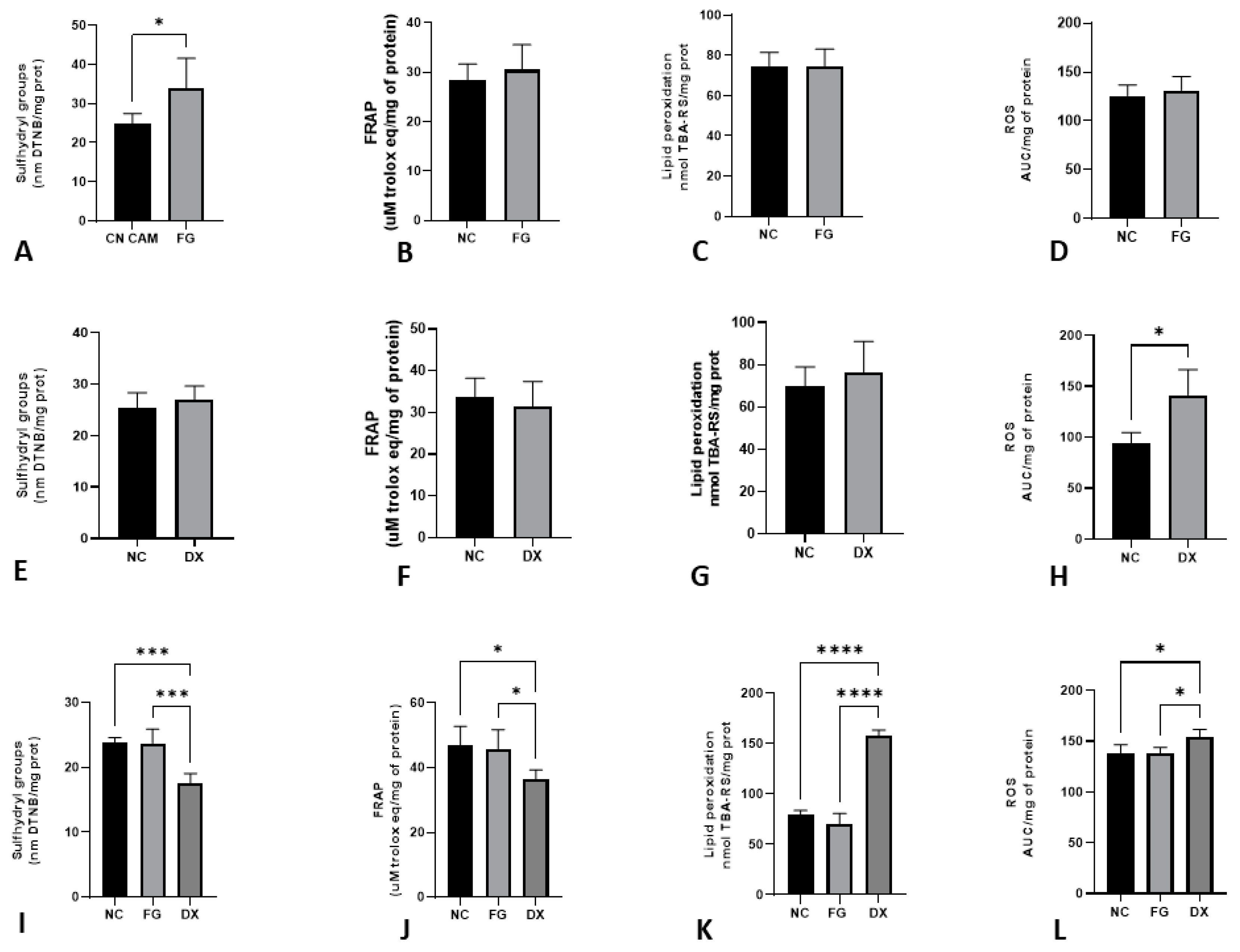
3.7. The Age of Inoculation Is an Essential Factor for Changing Oxidative Stress Parameters
3.8. There Is a Correlation between the Results for Haemoglobin Using a Drabkin Solution or Performing the Calculation of One Third of the Haematocrit
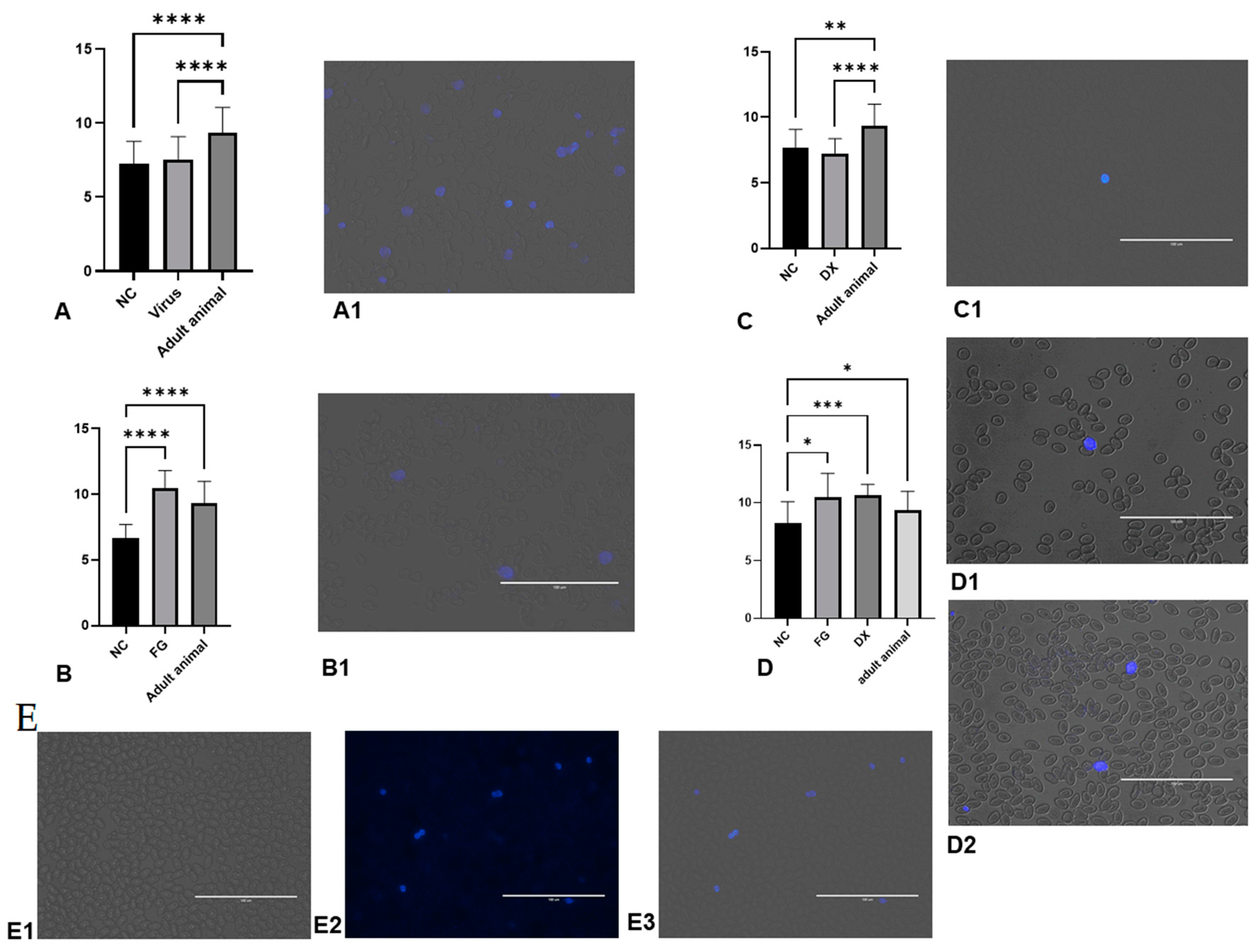
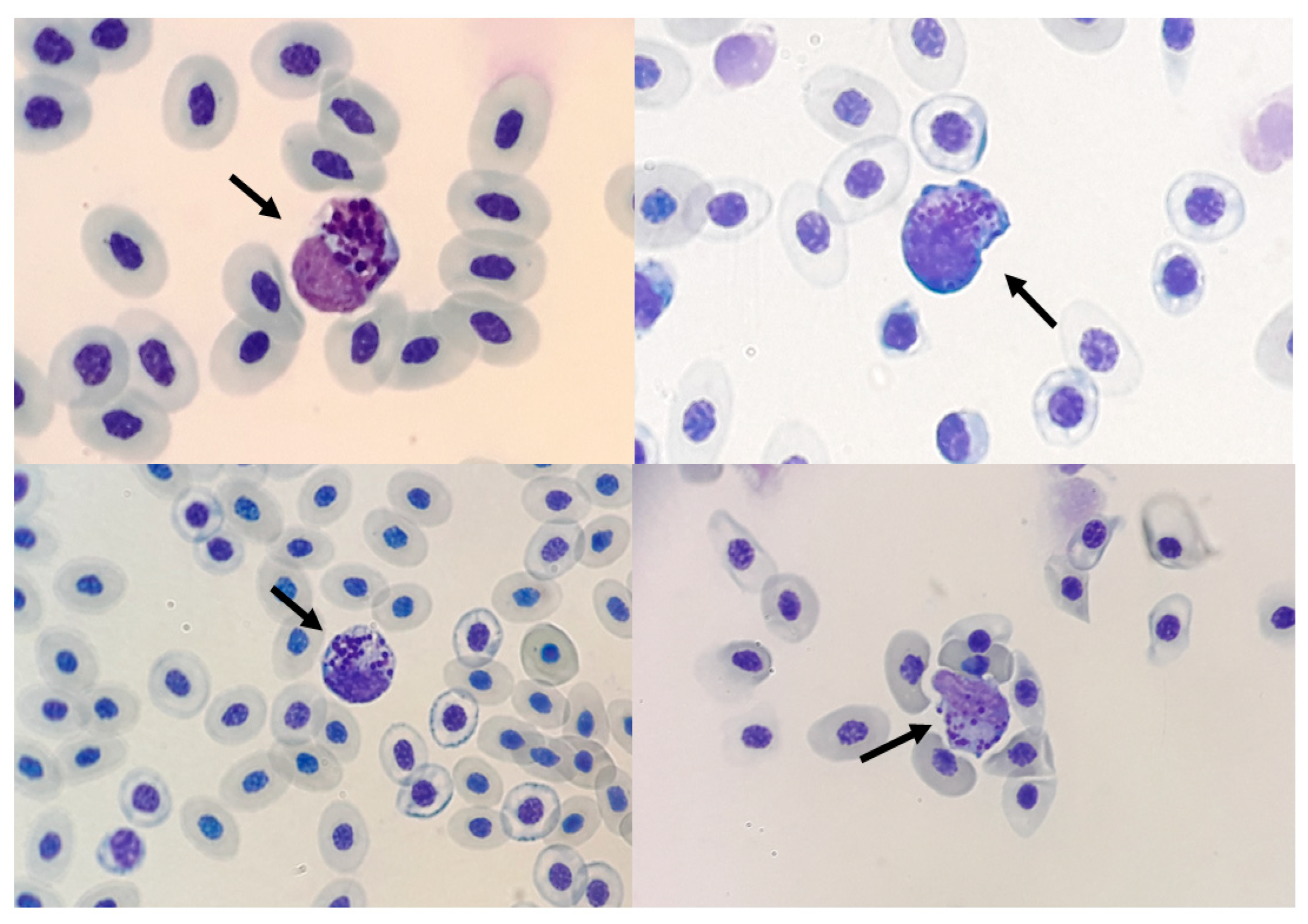
3.9. Characterisation of Granulocytes and Thrombocytes by Cytochemistry
3.10. Unidentified Granulocytes Were Found in Several Groups
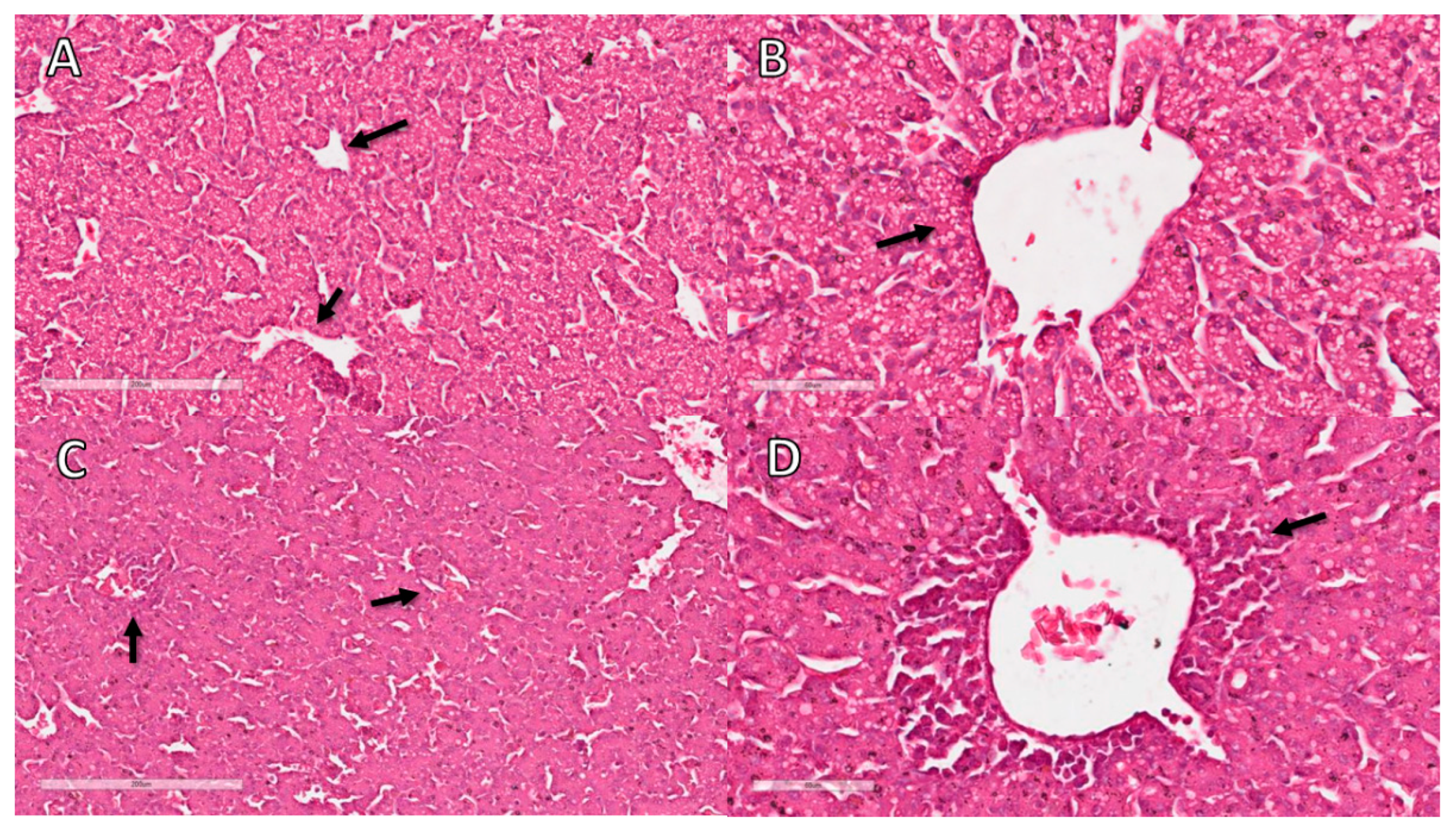
3.11. Liver Histopathological Analysis
3.12. FG in High Doses Does Not Cause Injury to Angiogenesis in CE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kain, K.H.; Miller, J.W.I.; Jones-Paris, C.R.; Thomason, R.T.; Lewis, J.D.; Bader, D.M.; Barnett, J.V.; Zijlstra, A. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. 2014, 243, 216–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokman, N.A.; Elder, A.S.F.; Ricciardelli, C.; Oehler, M.K. Chick chorioallantoic membrane (CAM) assay as an in vivo model to study the effect of newly identified molecules on ovarian cancer invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 9959–9970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halik, G.; Strojny, B.; Hotowy, A.; Grodzik, M.; Piast, R. Toxicity studies of six types of carbon nanoparticles in a chicken-embryo model. Int. J. Nanomed. 2017, 12, 2887–2898. [Google Scholar]
- Hruba, H.; Abdelsalam, E.E.E.; Anisimov, N.; Bandouchova, H.; Havelkova, B.; Heger, T.; Kanova, M.; Kovacova, V.; Nemcova, M.; Piacek, V.; et al. Reproductive toxicity of fluoroquinolones in birds. BMC Vet. Res. 2019, 15, 209. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, A.; Sharifi, I.; Tavakkoli, H.; Derakhshanfar, A.; Keyhani, A.R.; Salari, Z.; Mosallanejad, S.S.; Bamorovat, M. Embryonic toxico-pathological effects of meglumine antimoniate using a chick embryo model. PLoS ONE 2018, 13, e0196424. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Deng, G.; Guo, X.Q.; Kuang, J.; Zhang, C.; Cao, H.; Hu, G. Clinicopathology of Gout in Growing Layers Induced by Avian Nephrotrophic Strains of Infectious Bronchitis Virus. Pak. Vet. J. 2015, 35, 8318. [Google Scholar]
- de Morais Ribeiro, L.N.; de Paula, E.; Rossi, D.A.; Monteiro, G.P.; Junior, E.C.V.; Silva, R.R.; Franco, R.R.; Espindola, F.S.; Goulart, L.R.; Fonseca, B.B. Hybrid Pectin-Liposome Formulation against Multi-Resistant Bacterial Strains. Pharmaceutics 2020, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Haselgrübler, R.; Stübl, F.; Stadlbauer, V.; Lanzerstorfer, P.; Weghuber, J. An In Ovo Model for Testing Insulin-mimetic Compounds. J. Vis. Exp. 2018, 134, e57237. [Google Scholar] [CrossRef] [Green Version]
- Guy, J.S. Isolation and Propagation of Coronaviruses in Embryonated Eggs. Methods Mol. Biol. 2015, 454, 109–117. [Google Scholar]
- Fauzia, E.; Barbhuyan, T.K.; Shrivastava, A.K.; Kumar, M.; Garg, P.; Khan, M.A.; Robertson, A.A.B.; Raza, S.S. Chick Embryo: A Preclinical Model for Understanding Ischemia-Reperfusion Mechanism. Front. Pharmacol. 2018, 9, 1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Ko, M.L.; Huang, C.C.Y.; Park, S.Y.; Hong, M.P.; Wo, C.; Ko, G.Y.P. Chicken Embryos as a Potential New Model for Early Onset Type I Diabetes. J. Diabetes Res. 2014, 2014, 354094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronner-Fraser, M. Methods in Cell Biology: Avian Embryology, 2nd ed.; Elsevier: New York, NY, USA, 2017; pp. 1–432. [Google Scholar]
- Maier, H.J.; Bickerton, E.; Britton, P. Coronaviruses: Methods and protocols. Coronaviruses Methods Protoc. 2015, 1282, 1–282. [Google Scholar]
- Viana, F.A.B. Guia Terapêutico Veterinário, 3rd ed.; CEM: Lagoa Santa, Brazil, 2019; pp. 1–528. [Google Scholar]
- Collier, H.B. Standardization of Blood Haemoglobin Determinations. Can. Med. Assoc. J. 1944, 50, 550. [Google Scholar]
- Natt, M.P.; Herrick, C.A. A New Blood Diluent for Counting the Erythrocytes and Leucocytes of the Chicken. Poult. Sci. 1952, 31, 735–738. [Google Scholar] [CrossRef]
- Campbell, T.W. Hematology. In Avian Medicine: Principles and Application; Wingers Publishing: Lake Worth, FL, USA, 1997; p. 810. [Google Scholar]
- Thrall, M.A. Veterinary Hematology and Clinical Chemistry, 2nd ed.; Wiley: Hoboken, New Jersey, USA, 2012; p. 784. [Google Scholar]
- Swayne, D.E.; Stockham, S.L.; Johnson, G.S. Cytochemical Properties of Chicken Blood Cells Resembling Both Thrombocytes and Lymphocytes. Vet. Clin. Pathol. 1986, 15, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Justino, A.B.; Pereira, M.N.; Peixoto, L.G.; Vilela, D.D.; Caixeta, D.C.; Souza, A.V.; Teixeira, R.R.; Silva, H.C.G.; Moura, F.B.R.; Moraes, I.B.; et al. Hepatoprotective Properties of a Polyphenol-Enriched Fraction from Annona crassiflora Mart. Fruit Peel against Diabetes-Induced Oxidative and Nitrosative Stress. J. Agric. Food Chem. 2017, 65, 4428–4438. [Google Scholar] [CrossRef] [PubMed]
- Aksenov, M.Y.; Markesbery, W.R. Changes in thiol content and expression of glutathione redox system genes in the hippocampus and cerebellum in Alzheimer’s disease. Neurosci. Lett. 2001, 302, 141–145. [Google Scholar] [CrossRef]
- Franco, R.R.; Justino, A.B.; Martins, M.M.; Silva, C.G.; Campana, P.R.V.; Lopes, J.C.D.; Almeida, V.L.; Espindola, F.S. Phytoscreening of Vochysiaceae species: Molecular identification by HPLC-ESI-MS/MS and evaluating of their antioxidant activity and inhibitory potential against human α-amylase and protein glycation. Bioorg. Chem. 2019, 91, 103122. [Google Scholar] [CrossRef]
- Tolosa, E.M.C.; Rodrigues, C.J.; Behmer, O.A.; Freitas-Neto, A.G. Manual de Técnicas Para Histologia Normal e Patológica, 2nd ed.; Manole: São Paulo, Brazil, 2003; p. 331. [Google Scholar]
- Brunt, E.M.; Janney, C.G.; Di Bisceglie, A.M.; Neuschwander-tetri, B.A.; Bacon, B.R. Nonalcoholic steatohepatitis: A proposal for grading and staging the histological lesions. Am. J. Gastroenterol. Suppl. 1999, 94, 2467–2474. [Google Scholar] [CrossRef]
- Angulo, P. Nonalcoholic fatty liver disease. Encycl. Mol. Mech. Dis. 2009, 346, 1429. [Google Scholar]
- Fáncsi, T. Ultrastructural Studies of the Goose Embryo Liver. Anat. Histol. Embryol. 1982, 11, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Elfarnawany, M.H. Signal Processing Methods for Quantitative Power Doppler Microvascular Angiography. Ph.D. Thesis, The University of Western Ontario, London, ON, Canada, 2015. [Google Scholar]
- Decaro, N. Gammacoronavirus: Coronaviridae. In The Springer Index of Viruses; Springer: New York, NY, USA, 2011; Volume 100, p. 403. [Google Scholar]
- Schultze, B.; Cavanagh, D.; Herrler, G. Neuraminidase treatment of avian infectious bronchitis coronavirus reveals a hemagglutinating activity that is dependent on sialic acid-containing receptors on erythrocytes. Virology 1992, 189, 792–794. [Google Scholar] [CrossRef]
- Sheng, G. Primitive and definitive erythropoiesis in the yolk sac: A bird’s eye view. Int. J. Dev. Biol. 2010, 54, 1033–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Sun, M.; Deng, J.; Yu, J.; Yang, X.; Zhao, W.; Chen, G.; Wang, P. Zika virus induced more severe inflammatory response than dengue virus in chicken embryonic livers. Front. Microbiol. 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Frey, T.K.; Yang, J.J. Viral calciomics: Interplays between Ca2+ and virus. Cell Calcium 2009, 46, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Han, Z.; Shao, Y.; Geng, H.; Kong, X.; Liu, S. Proteomic analysis of chicken embryonic trachea and kidney tissues after infection in ovo by avian infectious bronchitis coronavirus. Proteome Sci. 2011, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiblum, R.; Arnon, E.; Chazan, G.; Robinzon, B.; Gvaryahu, G.; Snapir, N. Glucocorticoid Administration During Incubation: Embryo Mortality and Posthatch Growth in Chickens. Poult. Sci. 2001, 80, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, M.S.; Haug, A.; Torjesen, P.A.; Bakken, M. Prenatal exposure to corticosterone impairs embryonic development and increases fluctuating asymmetry in chickens (Gallus gallus domesticus). Br. Poult. Sci. 2003, 44, 690–697. [Google Scholar] [CrossRef]
- Mashaly, M.M. Effect of Exogenous Corticosterone on Chicken Embryonic Development. Poult. Sci. 1990, 70, 371–374. [Google Scholar] [CrossRef]
- Lin, H.; Decuypere, E.; Buyse, J. Oxidative stress induced by corticosterone administration in broiler chickens (Gallus gallus domesticus): 2. Short-term effect. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2004, 139, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Bartunek, P.; Karafiat, V.; Bartunkova, J.; Pajer, P.; Dvarakova, M.; Kralova, J.; Zenke, M.; Dvorak, M. Impact of chicken thrombopoietin and its receptor c-Mpl on hematopoietic cell development. Exp. Hematol. 2008, 36, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.K.; Cavey, M.J. Development of the liver in the chicken embryo. I. Hepatic cords and sinusoids. Anat. Rec. 1992, 234, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, M.H. Avian blood leucocyte responses to stress. Worlds. Poult. Sci. J. 1993, 49, 34–43. [Google Scholar] [CrossRef]
- Bellairs, R.; Osmond, M. The Atlas of Chick Development, 3rd ed.; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Vicuña, E.A.; Kuttappan, V.A.; Galarza-Seeber, R.; Latorre, J.D.; Faulkner, O.B.; Hargis, B.M.; Tellez, G.; Bielke, L.R. Effect of dexamethasone in feed on intestinal permeability, differential white blood cell counts, and immune organs in broiler chicks. Poult. Sci. 2012, 94, 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.C. Essentials of Veterinary Hematology, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 1993. [Google Scholar]
- Aengwanich, W. Effects of dexamethasone on physiological changes and productive performance in broilers. Asian J. Anim. Vet. Adv. 2007, 32, 157–161. [Google Scholar] [CrossRef]
- Elsayed, M.; Mohamed, N.; Hatab, M.; Elaroussi, M. Oxidative Stress of in-Ovo Ochratoxin A Administered during Chick Embryonic Development. Braz. J. Poult. Sci. 2019, 21, eRBCA-2019-0637. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Harrison, J.G.; Harrison, L.R. Avian Medicine: Principles and Application; Wingers Publishing: Lake Worth, FL, USA, 2009; p. 1384. [Google Scholar]
- Mandelker, L.; Vajdovich, P. Oxidative stress in applied basic research and clinical practice-Studies on experimental models. Eur. J. Histochem. 2012, 56, ejh.2012.br9. [Google Scholar]
- Lin, H.; Decuypere, E.; Buyse, J. Oxidative stress induced by corticosterone administration in broiler chickens (Gallus gallus domesticus): 1. Chronic exposure. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2004, 139, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M. A thermodynamic model for denaturation of granulocyte colony-stimulating factor: O-linked sugar chain suppresses not the triggering deprotonation but the succeeding denaturation. Biochim. Biophys. Acta Protein Struct. Mol. 1993, 1203, 295–297. [Google Scholar] [CrossRef]
- Mukaka, M.M. Statistics corner: A guide to appropriate use of correlation coefficient in medical research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar] [PubMed]
- Weiss, D.J.; Wardrop, K.J. Schalm’s Veterinary Hematology; John Wiley & Sons: Ames, IA, USA, 2010; p. 1206. [Google Scholar]
- Bussolino, F.; Ziche, M.; Wang, J.M.; Alessi, D.; Morbidelli, L.; Cremona, O.; Bosia, A.; Marchisio, P.C.; Mantovani, A. In Vitro and In Vivo Activation of Endothelial Cells by Colony-stimulating Factors. J. Clin. Investig. 1991, 87, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Bussolino, F.; Wang, J.M.; Defilippi, P.; Turrini, F.; Sanavio, F.; Edgell, C.-J.S.; Aglietta, M.; Arese, P.; Mantovani, A. Granulocyte and granulocyte-macrophage colony stimulating factors induce human endothelial cells to migrate and proliferate. Nature 1989, 337, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Bocchietto, E.; Guglielmetti, A.; Silvagno, F.; Taraboletti, G.; Pescarmona, G.P.; Mantovani, A.; Bussolino, F. Proliferative and Migratory Responses of Murine Microvascular Endothelial Cells to Granulocyte-Colony-Stimulating Factor. J. Cell. Physiol. 1993, 95, 89–95. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NC | Virus | |
|---|---|---|
| Ht (%) | 21.00 (±1.63) a | 15.60 (±2.30) b |
| Hg (g/dL) (1/3 Ht) | 7.00 (±0.55) a | 5.20 (±0.77) b |
| Hg (g/dL) (Cyanmethemoglobin) | 8.05 (±0.16) a | 6.23 (±0.99) b |
| Erythrocytes × 106/mm3 | 2.05 (±0.15) a | 1.59 (±0.23) b |
| MCH (pg) | 35.15 (±3.47) a | 40.08 (±9.95) a |
| MCV (fL) | 38.15 (±3.47) a | 40.08 (±9.95) a |
| MCHC (g/dL) | 96.96 (±17.07) a | 100.6 (±27.12) a |
| Thrombocytes × 104/mm3 | 2.21 (±0.51) a | 2.75 (±2.05) a |
| Leucocytes × 103/mm3 | 8 (±2.67) a | 20.4(±22.5) a |
| Monocytes/mm3 | 28 (±55) a | 254 (±465.5) a |
| Lymphocytes/mm3 | 650 (±730) a | 2170 (±2943) a |
| Heterophiles × 103/mm3 | 7.29 (±2.27) a | 17.6 (±18.8) a |
| Heterophile/lymphocyte | 24.09 (±18.53) a | 28.32 (±20.51) a |
| Eosinophils/mm3 | 0.00 (±0.00) a | 0.00 (±0.00) a |
| Unidentified granulocytes/mm3 | 23.75(±47.5) a | 259 (±459.1) a |
| Basophils/mm3 | 0.00 (±0.00) a | 0.00 (±0.00) a |
| NC (Serum) | Virus (Serum) | NC (AF) | Virus (AF) | |
|---|---|---|---|---|
| UA (mg/dL) | 21.76 (±23.13) a | 22.63 (±15.17) a | 18.53 (±21.53) a | 54.30 (±29.89) a |
| Creat (mg/dL) | 1.42 (±1.21) ab | 0.50 (±0.35) a | 3.21 (±1.41) b | 1.55 (±1.11) ab |
| ALP (U/L) | 1857 (±1108) a | 2258 (±1540) a | 80.40 (±26.37) b | 55.74 (±49.27) b |
| GGT (U/L) | 81.53 (±55.34) a | 252.4 (±121.5) a | 210.00 (±172.2) a | 184.00 (±119.40) a |
| AST (U/L) | 264.00 (±40.84) a | 451.00 (±281.1) a | 200.00 (±123.3) a | 592.00 (±283.10) a |
| ALT (U/L) | 57.50 (±233.6) a | 92.00 (±85.73) a | 63.00 (±38.1) a | 48.00 (±22.80) a |
| CRP (mg/L) | 24.55 (±10.31) a | 24.75 (±9.91) a | 59.50 (±32.55) a | 84.00 (±45.72) a |
| Ca (mg/dL) | 95.23 (±66.18) a | 10.23 (±8.08) b | 16.05 (±10.48) b | 16.60 (±13.77) b |
| P (mg/dL) | 6.82 (±3.70) a | 5.50 (±2.96) a | 19.19 (±8.57) b | 10.76 (±6.32) ab |
| NC | Virus | |
|---|---|---|
| Inflammation | 0.00 a (Mi: 0.00; Ma: 0.00) | 2.00 b (Mi: 0.00; Ma: 2.00) |
| Degeneration | 0.00 a (Mi: 0.00; Ma: 0.00) | 0.00 a (Mi: 0.00; Ma: 0.00) |
| Necrosis | 0.00 a (Mi: 0.00; Ma: 0.00) | 0.00 a (Mi: 0.00; Ma: 3.00) |
| Circulatory change | 0.00 a (Mi: 0.00; Ma: 0.00) | 1.00 b (Mi: 0.00; Ma: 3.00) |
| Alive (Normal) | Injured | Dead | Total | ||
|---|---|---|---|---|---|
| Zero EID | NC | 6 | 0 | 1 | 6 |
| FG | 5 | 0 | 1 | 6 | |
| DX | 0 | 2 | 4 | 6 | |
| 3 EID | NC | 6 | 0 | 0 | 6 |
| FG | 6 | 0 | 0 | 6 | |
| DX | 1 | 5 | 0 | 6 | |
| 7 EID | NC | 6 | 0 | 0 | 6 |
| FG | 6 | 0 | 0 | 6 | |
| DX | 0 | 3 | 3 | 6 | |
| 10 EID CAM | NC | 5 | 1 | 0 | 6 |
| FG | 6 | 1 | 0 | 7 | |
| 10 EID SM | CN | 6 | 0 | 0 | 6 |
| DX | 3 | 6 | 0 | 9 | |
| 12 EID | NC | 6 | 0 | 0 | 6 |
| FG | 6 | 0 | 0 | 6 | |
| DX | 3 | 3 | 0 | 6 |
| 10 EID | ||||
|---|---|---|---|---|
| NC (CAM) | FG (CAM) | NC (SM) | DX (SM) | |
| Ht (%) | 21.64 (±3.38) # | 18.43 (±3.60) # | 20.33 (±1.86) a | 17.13 (±2.35) b |
| Hg (g/dL) (1/3 ht) | 7.20 (±1.13) # | 6.14 (±1.20) # | 6.78 (±0.62) a | 5.71 (±0.78) b |
| Hg (g/dL) (Cyanmethemoglobin) | 8.12 (±0.66) # | 7.52 (±1.29) # | 7.83 (±0.65) a | 7.22 (±1.12) b |
| Erythrocytes × 106/mm3 | 1.76 (±0.53) # | 1.73 (±0.28) # | 2.05 (±0.27) a | 1.58 (±0.26) b |
| MCH (pg) | 50.34 (±14.07) # | 47.35 (±4.95) # | 38.35 (±2.04) a | 45.58 (±6.69) b |
| MCV (fL) | 107.1 (±13.82) # | 135.0 (±43.47) # | 99.73 (±8.53) a | 109.10 (±13.39) a |
| MCHC (g/dL) | 38.33 (±3.3) # | 41.00 (±3.48) # | 38.56 (±1.85) a | 42.12 (±2.65) b |
| Thrombocytes × 103/mm3 | 15.8 (±0.97) # | 13.3 (±13.1) # | 16.3 (±5.1) a | 8.75 (±5.75) b |
| Leucocytes × 103/mm3 | 3.68 (±2.06) # | 4.25 (±2.85) # | 4.15 (±0.74) a | 3.37 (±2.58) a |
| Monocytes/mm3 | 0.00 (±0.00) # | 14.17 (±33.09) # | 0.00 (±0.00) a | 0.00 (±0.00) a |
| Lymphocytes/mm3 | 159.01 (±97.72) # | 220.80 (±216.80) # | 318.5 (±326.3) a | 357 (±396.6) a |
| Heterophiles/mm3 | 3523 (±2020) # | 3973 (±2565) # | 3104 (±1251) a | 2453 (±1695) a |
| Heterophile/lymphocyte | 26.26 (±34.75) # | 32.85 (±31.77) # | 38.51 (±38.11) a | 36.80 (±43.29) a |
| Eosinophils/mm3 | 5.00 (±15.00) # | 25.53 (±69.48) # | 0.00 (±0.00) a | 1.18 (±6.03) a |
| Basophils/mm3 | 0.00 (±0.00) # | 0.00 (±0.00) # | 0.00 (±0.00) a | 0.00 (±0.00) a |
| 12 EID | |||
|---|---|---|---|
| CN (SM) | FG (SM) | DX (SM) | |
| Ht (%) | 27.80 (±3.56) A | 28.67 (±3.50) A | 23.40 (±3.71) A |
| Hg (g/dL) (1/3 ht) | 8.93 (±1.03) A | 9.55 (±1.17) A | 9.26 (±1.48) A |
| Hg (g/dL) (Cyanmethemoglobin) | 10.61 (±3.04) A | 10.55 (±1.48) A | 7.90 (±1.13) A |
| Erythrocytes × 106/mm3 | 2.59 (±0.48) A | 2.66 (±0.38) A | 1.90 (±0.26) B |
| MCH (pg) | 41.13 (±9.12) A | 39.65 (±2.12) A | 41.09 (±1.39) A |
| MCV (fL) | 102.8 (±25.82) A | 108.4 (±10.98) A | 128.6 (±10.29) A |
| MCHC (g/dL) | 37.75 (±7.09) A | 36.92 (±4.40) A | 33.28 (±3.18) A |
| Thrombocytes × 103/mm3 | 5.30 (±1.35) A | 5.66 (±1.08) A | 14.5 (±7.32) B |
| Leucocytes × 103/mm3 | 4.00 (±1.05) A | 4.96 (±1.78) A | 16.3 (±8.30) B |
| Monocytes/mm3 | 45.83 (±40.79) AB | 61.67 (±34.86) A | 40.00 (±132.70) B |
| Lymphocytes/mm3 | 457.60 (±114.80) A | 516.70 (±139.00) A | 271.90 (±139.80) B |
| Heterophiles/mm3 | 3498 (±940.50) A | 4132 (±1549) A | 16,900 (±8200) B |
| Heterophile/lymphocyte | 8.00 (±1.55) A | 7.00 (±3.30) A | 65.77 (±29.40) B |
| Eosinophils/mm3 | 0.00 (±0.00) A | 0.00 (±0.00) A | 0.00 (±0.00) A |
| Basophils/mm3 | 0.00 (±0.00) A | 0.00 (±0.00) A | 0.00 (±0.00) A |
| 10 EID | ||||
|---|---|---|---|---|
| CAM | ||||
| NC (Serum) | FG (Serum) | NC (AF) | FG (AF) | |
| UA (mg/dL) | 11.00 (±7.60) # | 16.49 (±14.55) # | 97.57 (±31.16) * | 87.88 (±51.40) * |
| Creat (mg/dL) | 1.31 (±0.68) # | 0.80 (±0.63) # | 2.49 (±0.85) * | 3.19 (±0.77) * |
| APL (U/L) | 1836 (±815.30) # | 2380 (±298.40) # | 47.05 (±32.58) * | 61.26 (±53.26) * |
| GGT (U/L) | 63.17 (±85.13) # | 67.00 (±81.56) # | 14.53 (±5.50) # | 12.49 (±6.98) # |
| AST (U/L) | 48.12 (±29.53) # | 49.18 (±15.01) # | 15.14 (±11.55) * | 9.33 (±6.02) * |
| ALT (U/L) | 54.02 (±51.67) #* | 158.00 (±150.10) # | 8.28 (±4.07) * | 11.71 (±8.51) * |
| Ca (mg/dL) | 18.80 (±17.03) # | 21.80 (±21.75) # | 12.50 (±7.05) # | 8.34 (±4.62) # |
| P (mg/dL) | 7.48 (±4.52) # | 4.52 (±2.58) # | 15.37 (±9.04) #* | 22.13 (±9.63) * |
| 10 EID | ||||
|---|---|---|---|---|
| SM | ||||
| NC (Serum) | DX (Serum) | NC (AF) | DX (AF) | |
| UA (mg/dL) | 13.86 (±10.37) ab | 12.04 (±9.84) a | 60.00 (±38.63) ab | 70.30 (±45.93) b |
| Creat (mg/dL) | 0.53 (±0.35) a | 1.12 (±1.03) a | 3.57 (±0.53) b | 2.62 (±1.29) b |
| APL (U/L) | 2125 (±1365) a | 4233 (±5830) a | 45.94 (±39.65) b | 77.81 (±54.46) b |
| GGT (U/L) | 81.20 (±94.30) a | 49.41 (±82.50) a | 12.72 (±11.31) a | 19.51 (±9.54) a |
| AST (U/L) | 37.55 (±20.84) a | 99.89 (±68.10) b | 39.87 (±55.81) a | 13.27 (±4.25) a |
| ALT (U/L) | 142.6 (±161.3) a | 142.8 (±138.7) a | 8.00 (±1.58) a | 13.00 (±8.69) a |
| Ca (mg/dL) | 9.15 (±10.65) a | 30.53 (±30.50) a | 6.50 (±2.80) a | 15.91 (±13.46) a |
| P (mg/dL) | 4.68 (±3.33) a | 5.80 (±4.21) a | 23.65 (±6.32) b | 26.90 (±11.91) b |
| 12 EID | |||
|---|---|---|---|
| SM | |||
| CN (Serum) | FG (Serum) | DX (Serum) | |
| UA (mg/dL) | 8.28 (±1.88) A | 12.53 (±10.17) A | 9.50 (±10.82) A |
| Creat (mg/dL) | 1.37 (±1.04) A | 0.70 (±0.53) A | 1.39 (±0.53) A |
| APL (U/L) | 2194 (±322.90) A | 2439 (±437.70) A | 1928 (±356.40) A |
| GGT (U/L) | 47.25 (±40.85) A | 59.84 (±64.05) A | 69.08 (±47.69) A |
| AST (U/L) | 123.1 (±81.82) A | 108.2 (±19.21) A | 556.1 (±27.61) A |
| ALT (U/L) | 56.68 (±59.33) A | 130.5 (±226.9) A | 73.70 (±25.15) A |
| Ca (mg/dL) | 17.98 (±9.35) A | 17.70 (±12.28) A | 13.63 (±10.32) A |
| P (mg/dL) | 5.22 (±3.89) A | 4.76 (±2.85) A | 9.52 (±3.28) A |
| 10 EID | 12 EID | ||||||
|---|---|---|---|---|---|---|---|
| NC (CAM) | FG (CAM) | NC (SM) | DX (SM) | NC | FG | DX | |
| Inflammation | 0 (Mi: 0; Ma: 0) # | 1 (Mi: 0; Ma: 2) * | 0 (Mi: 0; Ma: 0) a | 0 (Mi: 0; Ma: 2) b | 0 (Mi: 0; Ma: 0) A | 0 (Mi: 0; Ma: 1) AB | 1 (Mi: 0; Ma: 2) B |
| Degeneration | 0 (Mi: 0; Ma: 0) # | 0 (Mi: 0; Ma: 0) # | 0 (Mi: 0; Ma: 0) a | 0 (Mi: 0; Ma: 3) a | 0 (Mi: 0; Ma: 0) A | 0 (Mi: 1; Ma: 1) AB | 1 (Mi: 1; Ma: 3) B |
| Necrosis | 0 (Mi: 0; Ma: 0) # | 0 (Mi: 0; Ma: 0) # | 0 (Mi: 0; Ma: 0) a | 0 (Mi: 0; Ma: 2) a | 0 (Mi: 0; Ma: 0) A | 0 (Mi: 0; Ma: 0) A | 0 (Mi: 0; Ma: 0) A |
| Circulatory change | 0 (Mi: 0; Ma: 0) # | 0 (Mi: 0; Ma: 1) # | 0 (Mi: 0; Ma: 0) a | 1 (Mi: 0; Ma: 3) b | 0 (Mi: 0; Ma: 0) A | 0 (Mi: 0; Ma: 1) A | 0 (Mi: 0; Ma: 0) A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sommerfeld, S.; Mundim, A.V.; Silva, R.R.; Queiroz, J.S.; Rios, M.P.; Notário, F.O.; Medeiros Ronchi, A.A.; Beletti, M.E.; Franco, R.R.; Espindola, F.S.; et al. Physiological Changes in Chicken Embryos Inoculated with Drugs and Viruses Highlight the Need for More Standardization of this Animal Model. Animals 2022, 12, 1156. https://doi.org/10.3390/ani12091156
Sommerfeld S, Mundim AV, Silva RR, Queiroz JS, Rios MP, Notário FO, Medeiros Ronchi AA, Beletti ME, Franco RR, Espindola FS, et al. Physiological Changes in Chicken Embryos Inoculated with Drugs and Viruses Highlight the Need for More Standardization of this Animal Model. Animals. 2022; 12(9):1156. https://doi.org/10.3390/ani12091156
Chicago/Turabian StyleSommerfeld, Simone, Antonio Vicente Mundim, Rogério Reis Silva, Jéssica Santos Queiroz, Maisa Paschoal Rios, Fabiana Oliveira Notário, Alessandra Aparecida Medeiros Ronchi, Marcelo Emílio Beletti, Rodrigo Rodrigues Franco, Foued Salmen Espindola, and et al. 2022. "Physiological Changes in Chicken Embryos Inoculated with Drugs and Viruses Highlight the Need for More Standardization of this Animal Model" Animals 12, no. 9: 1156. https://doi.org/10.3390/ani12091156
APA StyleSommerfeld, S., Mundim, A. V., Silva, R. R., Queiroz, J. S., Rios, M. P., Notário, F. O., Medeiros Ronchi, A. A., Beletti, M. E., Franco, R. R., Espindola, F. S., Goulart, L. R., & Fonseca, B. B. (2022). Physiological Changes in Chicken Embryos Inoculated with Drugs and Viruses Highlight the Need for More Standardization of this Animal Model. Animals, 12(9), 1156. https://doi.org/10.3390/ani12091156