1. Introduction
Fibroblast growth factor 21 (FGF21) is a metabolic regulator produced mainly in the liver [
1], which is induced in response to multiple stressors, including energy deprivation, amino acid deprivation, exercise, inflammation, and metabolic disorders, such as obesity. As a consequence of these stress conditions, FGF21 has been demonstrated to preferentially stimulate metabolic pathways that play a central role in energy mobilization in human cells and rodent models, such as lipolysis, gluconeogenesis, and ketogenesis [
2,
3]. This indicates that the physiological function of FGF21 is to act as a stress hormone that aims to increase the availability of energy substrates in order to cope with the energy-consuming stress response [
4].
High-yield dairy cows commonly exhibit a pronounced negative energy balance (NEB) and are frequently exposed to different metabolic (e.g., non-esterified fatty acids (NEFA)) and inflammatory stimuli (e.g., bacterial compounds, inflammatory mediators) during the periparturient phase and early lactation [
5,
6,
7,
8]. In line with the role of FGF21 as a stress hormone, a dramatic induction of
FGF21 expression in the liver and marked increase in FGF21 concentration in plasma have been reported in cows at the day of parturition and during early lactation [
9,
10,
11,
12,
13,
14]. This suggests that FGF21 plays a particular physiological role during this phase and is involved in the metabolic adaptation to NEB and stress conditions in dairy cows. Induction of the expression of
FGF21 in the liver of dairy cows at the day of parturition and in the early lactation period is likely caused by increased hepatic uptake of NEFA released from white adipose tissue (WAT) into circulation, because NEFA are potent ligands of peroxisome proliferator-activated receptor α (PPARα), which acts as a transcriptional regulator of
FGF21 [
2,
15]. Likewise, overfeeding during the dry period, which stimulates lipolysis in WAT and thus increases NEFA levels in plasma during the postpartum phase, causes an increase in FGF21 plasma concentrations in dairy cows [
12,
16,
17]. In addition, evidence indicates that different cellular stress conditions, such as endoplasmic reticulum (ER) stress or inflammation, which frequently occur in the livers of cows during early lactation, cause the induction of hepatic
FGF21 expression [
18,
19]. In agreement with this, supplementation of polyphenols reduces not only hepatic ER stress but also the expression of
FGF21 in the liver of dairy cows during the first week of lactation [
20,
21,
22].
Several studies with obese and diabetic mouse models have consistently demonstrated that administration of murine or human recombinant FGF21 reduces the fat mass of the body and hepatic fat content by increasing energy expenditure in WAT and brown adipose tissue [
23,
24,
25], increasing the rate of β-oxidation of fatty acids, and reducing the rate of de novo-fatty acid synthesis [
26]. In addition, exogenous application of FGF21 to mice administered a high-fat diet improved oral glucose tolerance and insulin sensitivity [
27] due to increased secretion of adiponectin from WAT [
28]. Moreover, application of FGF21 improved the profile of plasma lipids (decline of low-density lipoprotein cholesterol and triacylglycerols, elevation of high-density lipoprotein cholesterol). While the function of FGF21 in pathologic rodent models has been extensively studied, knowledge about the function of FGF21 in dairy cows is limited, despite existing reports about the factors underlying induction of
FGF21 expression in dairy cows [
29].
Thus, in order to improve the understanding of the physiological function of FGF21 in dairy cows, the present study aimed to explore metabolic differences between high-yield dairy cows differing in their hepatic expression level of FGF21 (high vs. low hepatic FGF21 expression) in the early postpartum phase using liver transcriptomics, targeted plasma metabolomics, and supplemental analysis of parameters related to inflammation, the antioxidant system, and the occurrence of oxidative stress.
4. Discussion
In order to gain a deeper understanding of the role played by FGF21 in dairy cows during early lactation, this study compared performance and metabolic, inflammatory, and oxidative stress-related parameters in two groups of high-yield dairy cows that markedly differed in their hepatic expression levels of
FGF21 at week 1 postpartum. This time-point was chosen for two main reasons. First, the finding that the expression of
FGF21 is dramatically upregulated and plasma FGF21 concentrations are strongly increased during the first few days after parturition and strongly decline thereafter suggests that FGF21 exerts its main biological functions in dairy cows during the first week postpartum [
9,
10,
11,
12,
13,
45]. Second, based on the suggestion that FGF21 acts as a stress hormone released from the liver to cope with stress conditions [
4], we expected that the biological effects of FGF21 would be most prominent during the phase in which cellular stress is most pronounced. Previous studies have shown that stress conditions, such as ER stress or inflammation, are most enhanced during the first week after parturition and strongly decline thereafter [
46,
47]. At week 5 postpartum, stress conditions such as ER stress and inflammation are already nearly absent [
46,
47,
48].
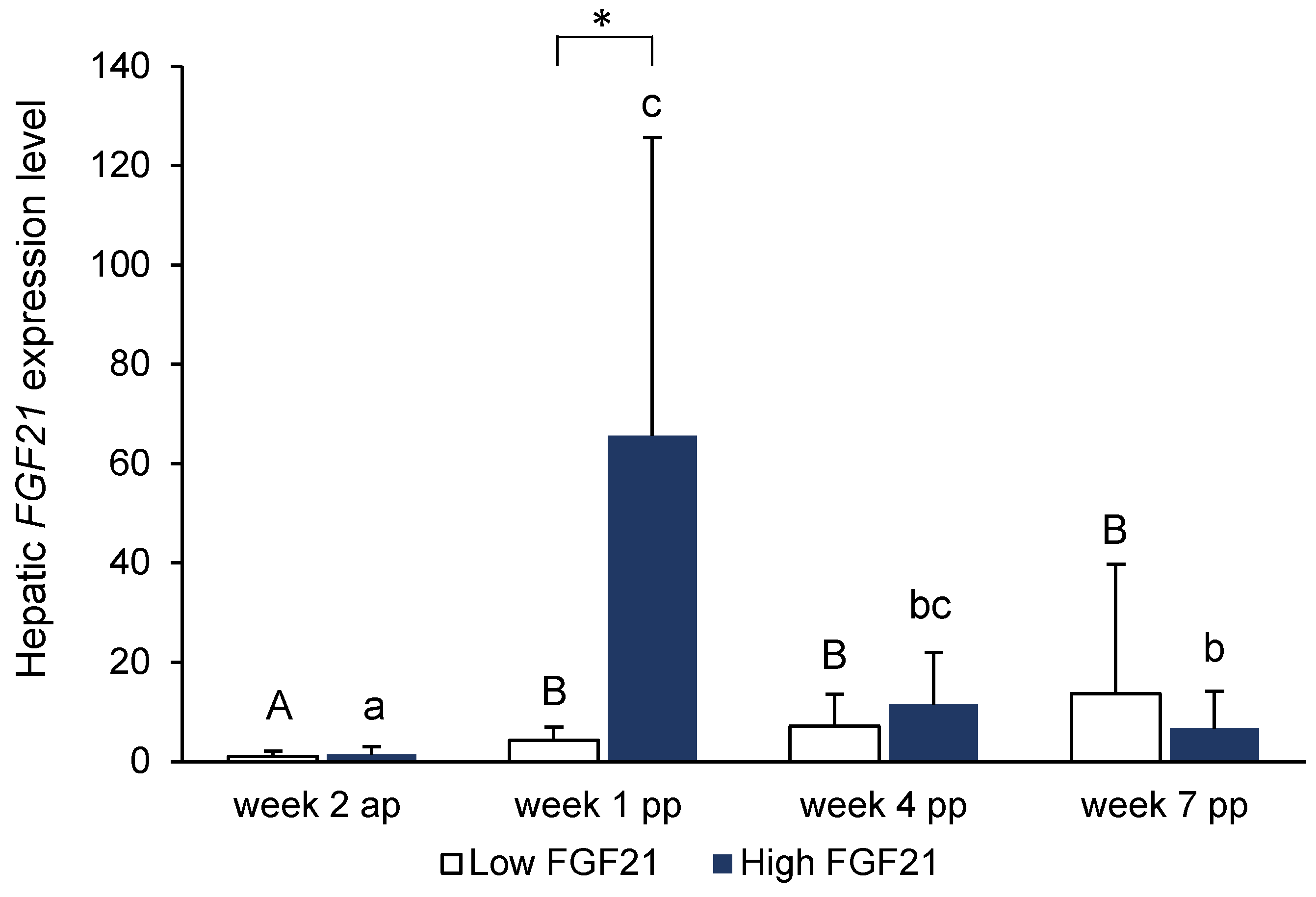
Analysis of
FGF21 expression in the liver in a cohort of 30 cows in our study demonstrated that the increase in
FGF21 expression at week 1 postpartum showed great individual variation. While some cows showed a dramatic increase compared to the antepartum level, hepatic
FGF21 expression in others remained nearly unchanged. Our study, however, confirmed the findings of other studies in which cows with strongly increased expression levels of
FGF21 in early lactation showed strongly declining expression levels towards later lactation [
10,
11]. In order to identify factors that could be involved in the expression of
FGF21 and to figure out a possible role of FGF21 in metabolic regulation, we selected the 8 cows with the highest hepatic
FGF21 expression and the 8 cows with the lowest expression at week 1 postpartum out of a cohort of 30 cows, and assigned them to two groups. Performance parameters, such as dry matter intake, net energy intake, milk yield, and ECM, body weights, and energy balance did not differ between these two groups at days 8–14 postpartum; thus, it can be excluded that the performance level or energy balance of the cows caused the differential expression of hepatic
FGF21 in these two groups. Although the strong induction of
FGF21 expression in the liver of dairy cows in early lactation is considered to be caused by the pronounced hepatic uptake of NEFA released from WAT during this phase [
10,
11,
13], the stronger induction of
FGF21 expression in the cows with high hepatic
FGF21 expression was probably not due to greater intrahepatic availability of NEFA because plasma concentrations of NEFA did not differ between the groups. Moreover, other metabolic parameters characterizing energy balance and hepatic lipid metabolism, such as plasma concentrations of BHBA, cholesterol, and TAG, and hepatic concentrations of cholesterol and TAG, did not differ between the two groups. This clearly indicated that the marked difference in hepatic
FGF21 expression between the two groups of cows was not caused by differences in energy mobilization and hepatic lipid metabolism. The finding that body weight loss during the early lactation period also did not differ between the two groups of cows supported the indication that the difference in hepatic
FGF21 expression was not due to a different rate of body fat mobilization.
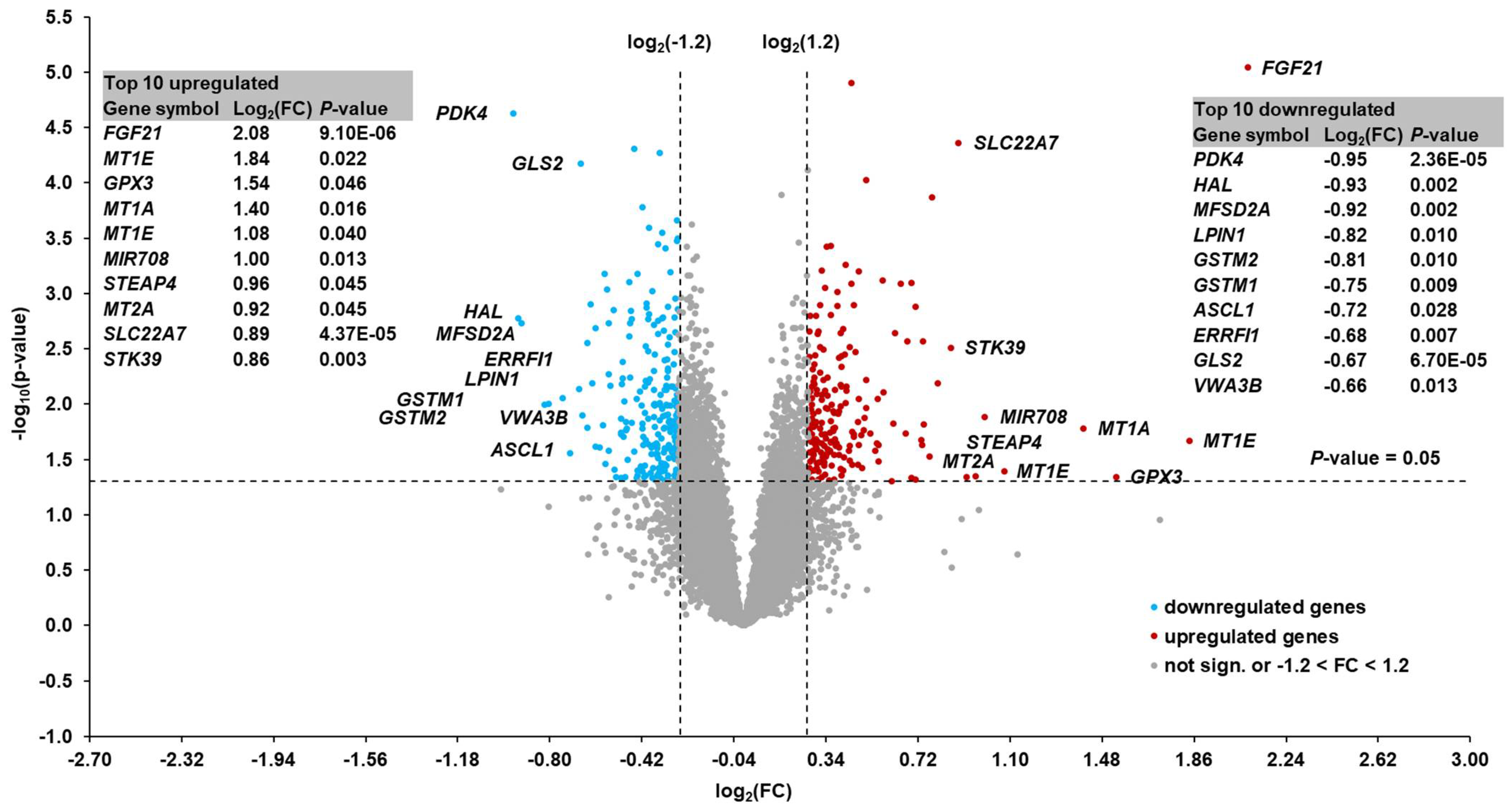
In order to identify metabolic pathways influenced by hepatic
FGF21 expression at week 1 postpartum, we performed transcriptomics analysis using microarray technology. The RNA integrity measurement indicates the presence of some kind of RNA degradation that can limit sensitivity (the rate of detection of true positives among all positives) and specificity (the rate of detection of true negatives among all negatives) of microarray performance [
49]. Nevertheless, the mean RIN value >6 suggested that the RNA integrity was sufficient to carry out microarray hybridizations. In addition, the microarray analysis was primarily used as a screening technique to identify changes in sets of pathway-specific genes, rather than as a tool to accurately quantify the expression of differentially regulated genes. To obtain deeper insight into the main metabolic pathways of energy metabolism that might differ between the two groups, the microarray expression levels of genes associated with β-oxidation and mitochondrial fatty acid import, gluconeogenesis, ketogenesis, and tricarboxylic acid were evaluated. Due to the low number of selected cows with low and high hepatic
FGF21 expression (
n = 8), this approach was preferred rather than carrying out principal component analysis or partial least-squares discrimination analysis, both of which require larger sample sizes to detect differences in discriminant genes. Our analysis showed that very few of these genes were differentially regulated between cows with high and low hepatic
FGF21 expression and the regulation of these genes was moderate [FC between −1.32 (
FBP1) and +1.34 (
CS)]. In order to determine whether
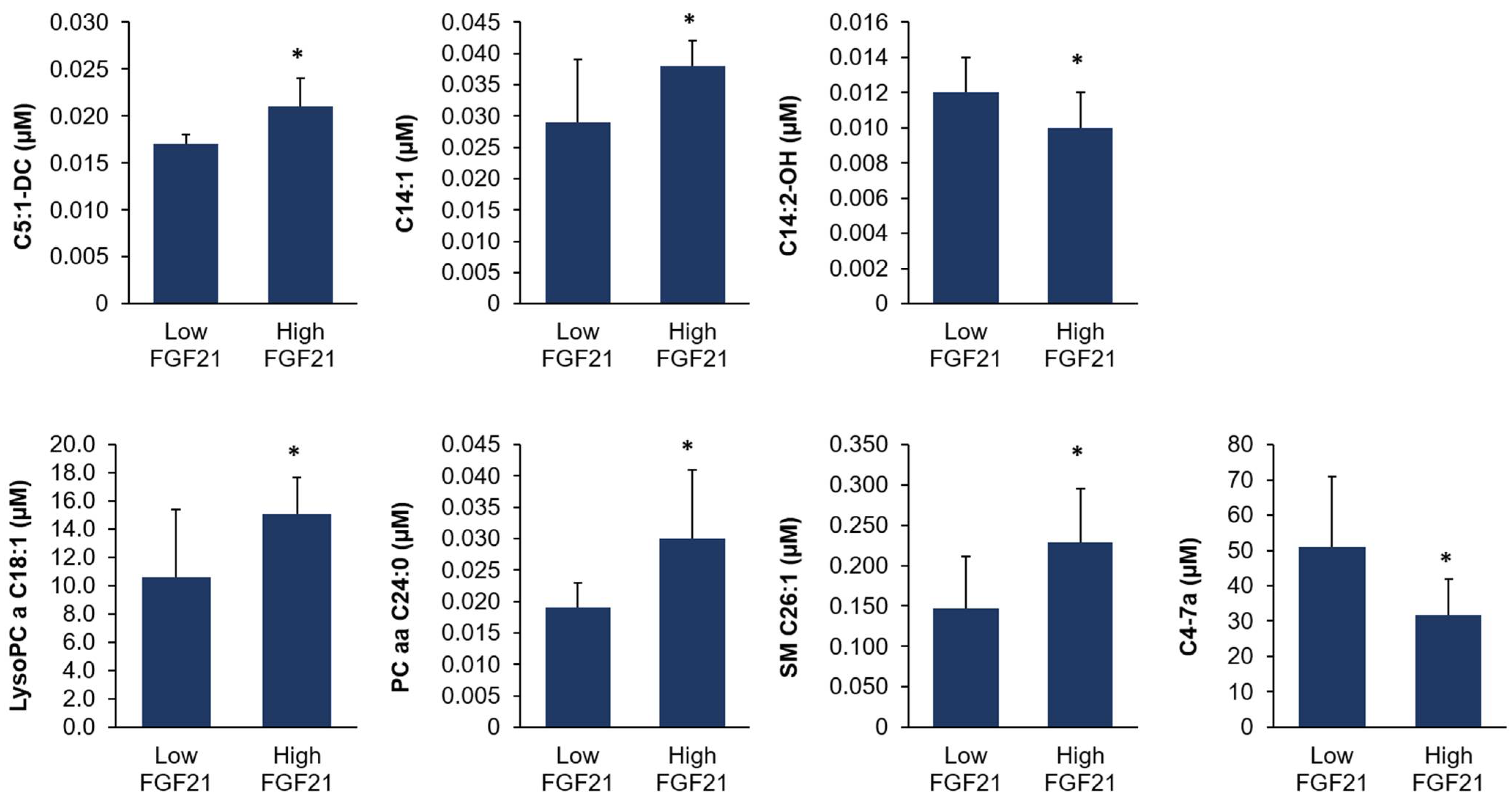
FGF21 expression in the liver was related to alterations on a metabolic level, we performed targeted plasma metabolomics analysis. We observed that the concentrations of only seven out of >200 metabolites were significantly different between the two groups of cows. These seven metabolites belonged to four different metabolite classes (acylcarnitines, glycerophospholipids, sphingolipids, and sterols). Acylcarnitines play a role in the β-oxidation of fatty acids within the mitochondrion. However, acylcarnitines represented only minor species with differing plasma concentrations between the two groups of cows and the concentrations of total carnitine and acetylcarnitine in plasma did not differ between the two group of cows. Therefore, there was no overall indication that
FGF21 expression affected the β-oxidation of fatty acids by influencing carnitine metabolism. In a similar manner, the three individual phospholipid species and sterol derivative with differing plasma concentrations between the two groups of cows were minor components, suggesting that
FGF21 expression was not linked to substantial effects on phospholipid and sterol metabolism. The observation that the small number of regulated plasma metabolites belonged to four different metabolite classes was a further indication that the hepatic expression level of
FGF21 was not associated with the consistent regulation of a specific metabolic pathway. This showed that cows with high and low hepatic
FGF21 expression did not differ with regard to hepatic energy metabolism, indicating that other factors must be responsible for the marked difference in hepatic
FGF21 expression. Likewise, the application of exogenous FGF21 had no effect on fatty acid metabolism in dairy cows [
50]. In addition, FGF21 administration in dairy cows did not affect plasma concentrations of insulin and glucose and insulin concentrations in a glucose tolerance test [
51], indicating that glucose metabolism was also not affected by FGF21 in dairy cows. An important reason for the lack of effect of exogenous FGF21 and lack of differences in the main metabolic pathways of energy metabolism between cows with high and low hepatic
FGF21 expression might be that the liver, unlike WAT, is not the primary tissue targeted by the action of FGF21. This is reflected by the fact that the physiologic FGF21 receptor FGFR1c, which is strongly expressed in WAT, is nearly absent in cattle liver whereas β-klotho, the co-activator of FGF21, is expressed in cattle liver at a markedly lower level than in WAT [
10].
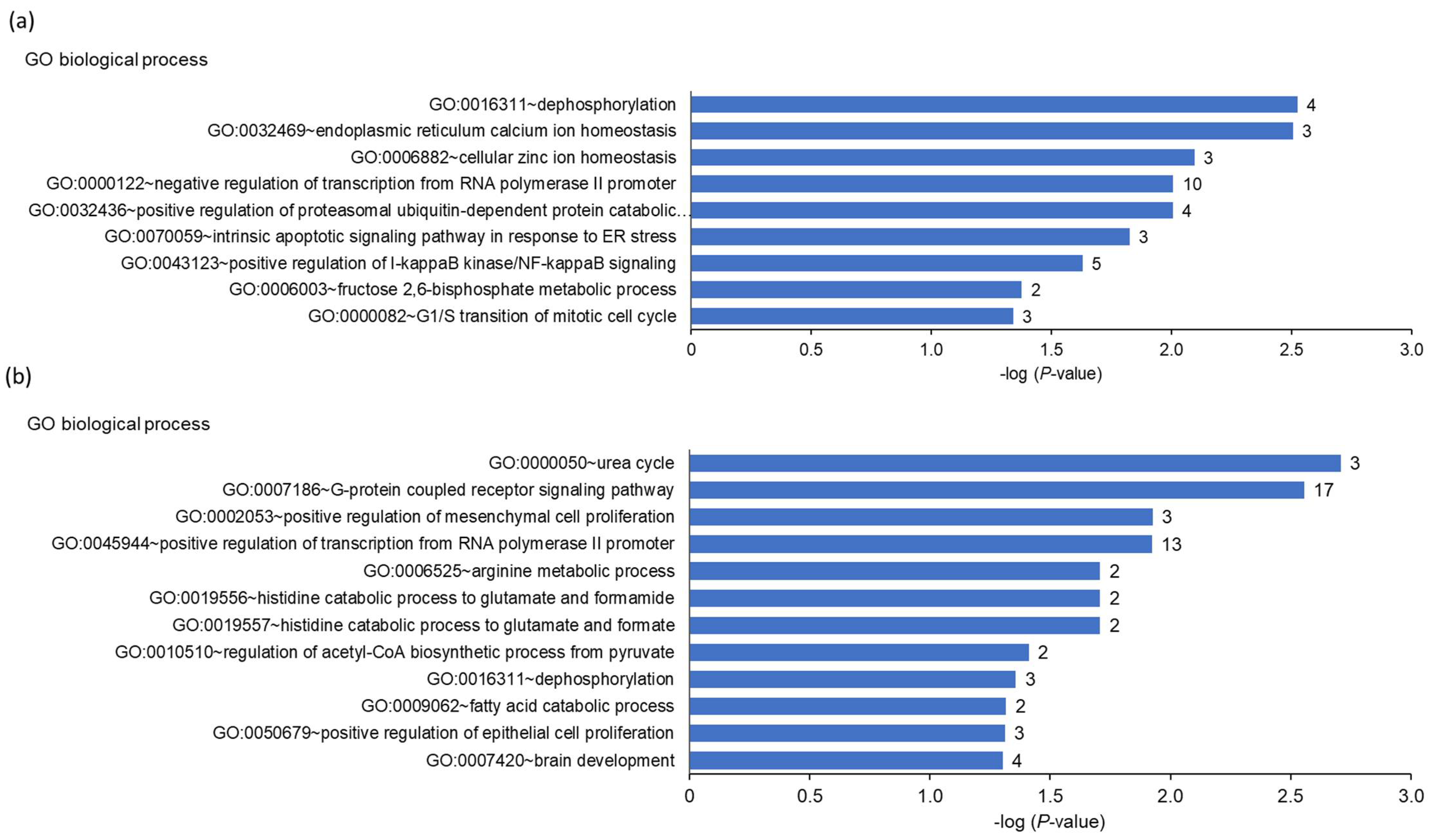
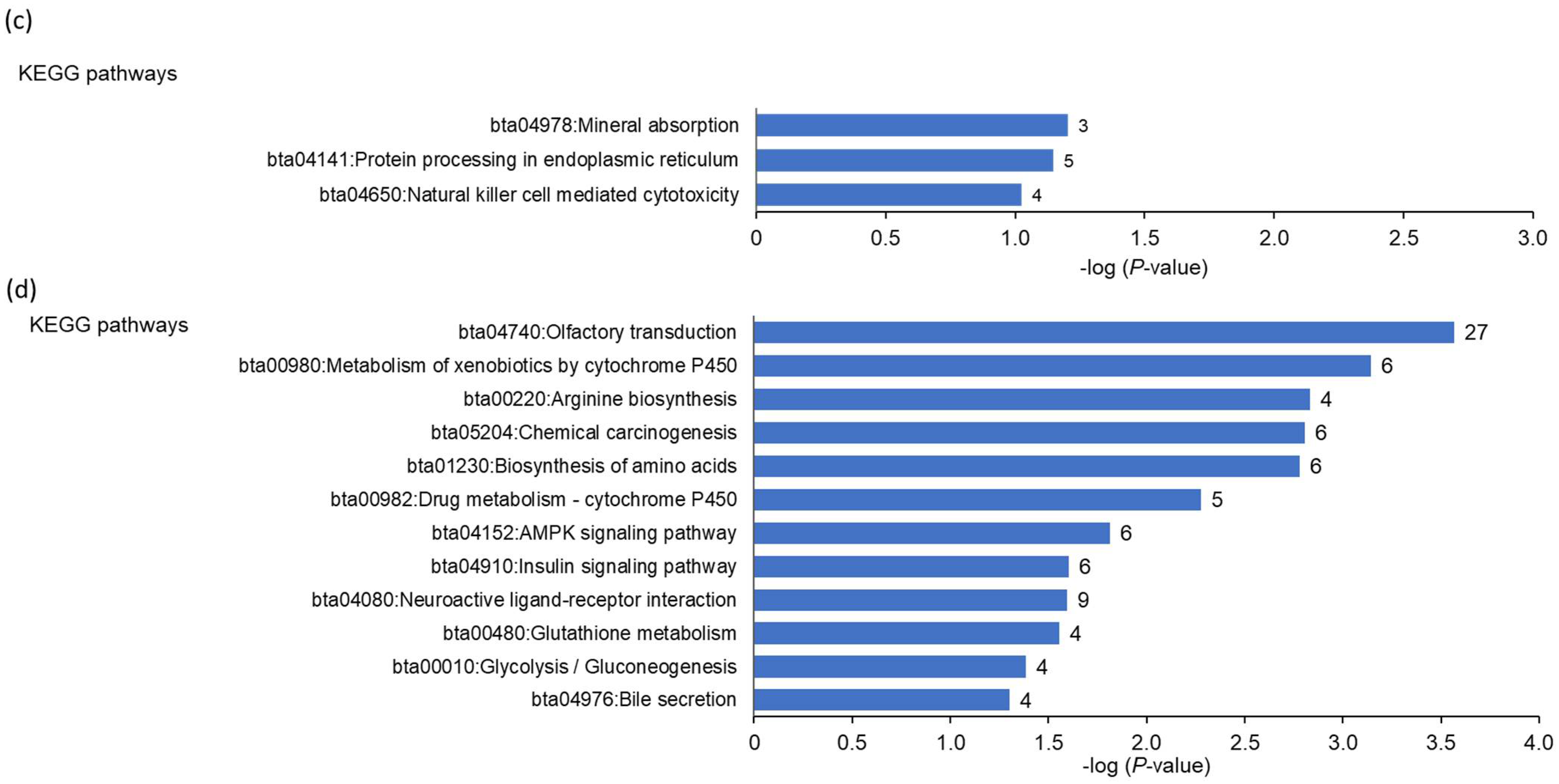
Previous studies have shown that dairy cows are subject to ER stress during early lactation, a phenomenon that could be involved in the development of fatty liver and metabolic diseases during this period [
47,
52,
53,
54]. With respect to this finding, an interesting observation from the liver transcriptomics of the present study is that the upregulated transcripts in the cows with high hepatic
FGF21 expression were involved particularly in the ER stress-induced unfolded protein response (UPR). This was evident from the gene set enrichment analysis, which demonstrated that the intrinsic apoptotic signaling pathway in response to ER stress, response to nutrient levels, response to extracellular stimulus, response to starvation, programmed cell death, and protein processing in the ER were amongst the most enriched biological process terms and KEGG pathways within the genes upregulated in the cows with high hepatic
FGF21 expression. In line with this, the upregulated transcripts in cows with high hepatic
FGF21 expression included several typical ER stress-target genes, such as
ATF4, HERPUD1, HSPA5, DDIT3, WFS1, DNAJB11, CHAC1, and
TRIB3. ER stress is well-known to be activated as a consequence of different stress conditions, such as nutrient deprivation, inflammation, and oxidative stress, which cause imbalances in ER quality control pathways leading to the accumulation of unfolded or misfolded proteins in the ER [
55]. In order to combat ER stress, mammalian cells are equipped with a defense system, called UPR, which enables the cell to re-establish ER quality control and decrease the accumulation of unfolded or misfolded proteins in the ER [
56]. Apart from protein kinase RNA-like ER kinase (PERK)-dependent transient attenuation of new protein synthesis and stimulation of inositol-requiring protein 1a (IRE1)-dependent mRNA degradation, another important UPR mechanism is the targeting of unfolded or misfolded proteins towards the ER-associated degradation pathway, where they are transferred to the cytosol and degraded by the ubiquitin proteasome system (UPS) [
57]—the most important pathway of intracellular protein degradation. In agreement with this UPR mechanism, the gene set enrichment analysis also revealed that regulation of protein catabolic process, positive regulation of proteasomal ubiquitin-dependent protein catabolic process, regulation of protein metabolic process, and positive regulation of proteasomal protein catabolic process were overrepresented terms amongst the genes upregulated in the cows with high hepatic
FGF21 expression. Remarkably, a recent study convincingly demonstrated that FGF21 plays an important role in muscle atrophy during fasting, reporting that the muscle mass of wild-type mice was significantly reduced in response to fasting, whereas muscle-specific FGF21 knockout mice were protected against muscle loss and weakness during fasting [
58]. In addition, the authors showed that in vivo FGF21 overexpression in skeletal muscle induced muscle atrophy, thus supporting a role for FGF21 in skeletal muscle proteolysis. Despite that the authors of this study did not study a possible involvement of ER stress, their results indicated that
FGF21 expression was associated with increased protein catabolism.
If ER stress is overwhelming and ER homeostasis cannot be restored, the UPR can also activate signaling pathways that initiate programmed cell death. Convincing evidence has been gained from cell culture studies and studies with laboratory animals that ER stress-induced UPR leads to an upregulation of hepatic
FGF21 expression through the activation of ER stress-transducers PERK and IRE1 [
18,
19]. Based on these findings, it is likely that ER stress was responsible for the strong induction of
FGF21 expression in the cows with high hepatic
FGF21 expression. Previous observations with dairy cows, in which polyphenol-enriched feeding rations decreased not only the hepatic expression of UPR-associated genes but also the expression of FGF21 [
20,
21,
22], are supportive of such a relationship between hepatic ER stress and FGF21 production.
Activation of nuclear factor E2-related factor 2 (Nrf2) in the liver of dairy cows during early lactation was shown in a previous study [
46]. Nrf2 is a transcription factor that regulates the expression of a broad range of antioxidant and cytoprotective genes [
59]. Hepatic transcript profiling in the present study revealed the upregulation of several target genes of Nrf2, including
GPX3, MT1A, MT2A, and
MT1E, in cows with high hepatic
FGF21 expression. Because activation of Nrf2 signaling is a known downstream event of ER stress, which aims to counteract oxidative stress and inflammatory conditions which are frequently associated with ER stress [
60,
61], induction of Nrf2-dependent genes is likely a consequence of the induction of ER stress-related genes and a further indicator of the occurrence of ER stress in the cows with high hepatic
FGF21 expression. It has been well established that ER stress is linked to the induction of pro-inflammatory conditions [
62]. Therefore, we also considered genes associated with inflammation in the liver and concentrations of acute phase proteins in plasma. We observed that there was an up-regulation of two genes encoding serum amyloid A (
SAA2, SAA4) in the liver and an increased concentration of SAA in plasma of the cows with high
FGF21 expression in the liver compared to cows with low
FGF21 expression in the liver. SAA belongs to the group of acute phase proteins produced in the liver and released into the blood in the course of inflammation [
63]. Therefore, increased production of SAA in the liver of the group of cows with high hepatic
FGF21 expression indicates that these cows could have suffered pro-inflammatory conditions in the liver. However, the expression of several other genes encoding proteins involved in inflammation (such as
TNF) and acute phase reaction (
CP,
HP) in the liver did not differ between the two groups of cows, indicating that the increased expression of
FGF21 was not related to the development of a pro-inflammatory condition. Moreover, plasma concentrations of oxylipids, including 9-HODE, 13-HODE, 12-HETE, 15-HETE, LTB4, and PGF2α, also did not differ between the two groups of cows. Oxylipids are a class of lipid mediators produced by enzymatic and non-enzymatic oxidation of PUFA, dominantly linoleic acid (LA) and arachidonic acid (AA). LA-derived 9-HODE and 13-HODE, but also AA-derived HETEs and eicosanoids, promote inflammatory responses by acting as chemoattractants for circulating immune cells [
64]. Thus, the present data suggest that the cows with high hepatic
FGF21 expression did not exhibit a systemic inflammatory process. In addition, the cows with high and low hepatic
FGF21 expression did not differ with regard to the plasma concentrations of antioxidants, such as tocopherols, β-carotene, and GSH, and indicators of antioxidant status, such as TEAC, TBARS, and protein carbonyls. This indicated that the cows with high hepatic
FGF21 expression also did not experience systemic oxidative stress, which is frequently associated with inflammation. Albeit speculative, it is possible that the induction of Nrf2-dependent genes observed in the cows with high hepatic
FGF21 expression prevented the development of systemic oxidative stress, because activation of Nrf2 signaling activated the expression of genes encoding cytoprotective enzymes, including antioxidant enzymes, thereby providing protection against ROS generated during the inflammatory process. This assumption is supported by recent evidence that FGF21 inhibits oxidative stress by stimulating Nrf2-dependent induction of cytoprotective and antioxidative genes [
65,
66,
67]. This would suggest that FGF21 plays an important role in the adaptation to cellular stress conditions in dairy cows during early lactation—a phase during which high-yield dairy cows are confronted with several different stress stimuli [
5,
6,
7,
8]. Indeed, our assumption of FGF21 as a stress-modulating factor in early-lactation dairy cows is supported by the results from a mice study that suggested that
FGF21 induction was a potential strategy to protect against the toxicity resulting from experimentally induced sepsis [
68]. While the upregulated genes in the cows with high
FGF21 expression were found to be involved particularly in ER stress-induced UPR, the downregulated genes in the cows with high hepatic
FGF21 expression were identified to play a role in the cellular amino acid catabolic process, alpha-amino acid metabolic process, glutamine family amino acid metabolic process, arginine biosynthesis, and urea cycle. This indicated that hepatic amino acid catabolism was attenuated in the cows with high compared to low hepatic
FGF21 expression. Although speculative, this might be interpreted as a mechanism to provide amino acids for specific organ functions, such as acute phase protein synthesis, which are activated as a consequence of ER stress, as seen in the cows with high hepatic
FGF21 expression. Overall, our data indicate that induction of hepatic
FGF21 in cows after parturition (1 week postpartum) might be, at least in part, caused by cellular stress such as ER stress. In turn, FGF21 might induce hepatic signaling pathways, such as Nrf2, which helps the body to cope with stress conditions and protect the liver against damage induced by the inflammation process and increased generation of reactive oxygen species commonly observed in dairy cows after parturition [
8,
69].

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}