1. Introduction
Over the last decades, the aquaculture sector has been interested in the reduction of unsustainable fish meal (FM) and fish oil (FO) in aquafeed formulations to promote alternative and more sustainable ingredients [
1,
2,
3]. Subsequently, a number of new ingredients have been searched and tested over the last years, including vegetable ingredients [
4], processed animal proteins (PAPs) [
5,
6] and dried microbial biomass from microalgae [
7,
8,
9,
10,
11].
More recently, insect meal has gained attention in aquaculture nutrition [
2,
12,
13,
14,
15]. In this context, the black soldier fly (
Hermetia illucens) has been widely considered [
16,
17,
18,
19,
20,
21] due to (i) a proper protein content (approximately 32%) [
22] and an essential amino acid (EAA) profile close to FM [
23,
24,
25]; (ii) a high feed-conversion efficiency, substrate consumption and waste reduction index of the larvae [
26,
27]; (iii) the ability to grow on different organic by-products [
28,
29]; and the low environmental impact of rearing culture (i.e., land use, water consumption and CO
2 production) [
30,
31]. These properties allow the application of the circular economy concept to the aquaculture sector, using for example land-produced organic by-product for insect rearing. Furthermore, it is well established that black soldier fly larvae/prepupae naturally contain different molecules (i.e., chitin, antimicrobial peptides, and lauric acid) that can play an immunostimulant and/or anti-inflammatory role in fish [
15,
32,
33,
34].
However, regarding the lipid content, the fatty acids (FA) profile of black soldier fly prepupae meal (HM) is characterized by a low level of long-chain polyunsaturated fatty acids (PUFA) and a high quantity of saturated ones (SFA) [
33,
35,
36,
37]. Nevertheless, the FA profile of the larvae can be partially modulated by their diet [
38,
39], potentially overcoming the drawback related to their unbalanced SFA/PUFA ratio [
40,
41,
42,
43]. Hence, the inclusion of the dried biomass from a marine protist rich in long-chain PUFA (
Schizochytrium sp., 10%
w/
w) in the growth substrate of black soldier fly larvae has been shown to successfully improve the PUFAs content of the insects’ final biomass [
39].
On this regard, the dried biomass of spirulina (
Arthrospira platensis) can represent an equally valid supplement for the insect growth substrate because of its nutritional values [
44,
45]. In fact, spirulina, the most commonly cultured cyanobacteria produced at commercial scale [
46], shows a high content of PUFAs for fish requirement [
47,
48]. Furthermore, spirulina contains several compounds such as vitamins, minerals, tocopherols, and carotenoids which can have a positive antioxidant effect on fish [
49,
50,
51].
The low inclusion (1 to 15%) of spirulina in fish diets has already been tested in different studies [
52,
53,
54] showing that it is able to improve fish growth, survival rate, feed intake, and fillet color and firmness. However, higher dietary inclusion levels led to controversial results regarding fish-growth performances, mostly related to a reduced feed acceptance [
55,
56,
57,
58]. In addition, the 9% dietary inclusion of spirulina in diets totally deprived of FM was not able to fully counteract the adverse effects of certain plant protein-rich ingredients on the gut health status of rainbow trout (
Oncorhynchus mykiss) [
59].
These results, in addition to its high production costs [
60], suggest that spirulina, instead of an aquafeed ingredient or supplement in aquafeed formulation, could be used to enrich the growth substrate of black soldier fly larvae, giving the possibility to exploit its beneficial properties avoiding a direct inclusion in fish diets. This aspect is also crucial to avoid the regularly used defatting processes applied to insect meal, which is one of the main reasons of the high final costs of this new ingredient.
For these reasons, this study aimed to enrich the insects’ rearing substrate (in terms of PUFAs and antioxidants molecules contents) with a 15%
w/
w inclusion of spirulina and then use the obtained full-fat prepupae meal to replace 0, 3 or 20% of FM and FO in experimental diets (named HM0, HM3, and HM20, respectively) intended for rainbow trout (which represents 53% of the total freshwater European aquaculture production) [
61]. The two selected inclusion levels were chosen according to the range used over the last year in studies on rainbow trout [
62]; this was because 20% represents an ecological inclusion level (a global 20% FM substitution in aquafeeds has a proper ecological impact), while the 3% substitution level should be considered as a feed supplement inclusion with possible ameliorative effects on fish health status. A multidisciplinary approach was then applied to evaluate fish zootechnical performances, gut and liver health status, and quality-marketable characteristics.
2. Materials and Methods
2.1. Ethics
Fish manipulation was performed following the procedures established by the European Communities Council Directive (86/609/EEC and 2010/63/EU) for animal health and approved by the Camerino’s University Animal Ethics Committee (Approval Number 6/2021). Optimal rearing conditions were ensured, and animal suffering was minimized using the MS222 anesthetic (Merck KGaA, Darmstadt, Germany).
2.2. Insects’ Rearing and Production of Fish Diets
Full-fat black soldier fly prepupae were obtained after rearing phase performed at the D3A Department of the Marche Polytechnic University (Ancona, Italy). Briefly, black soldier fly larvae were reared for 21 days in a climatic chamber (27 ± 1 °C; 65 ± 5% relative humidity) in continuous darkness following the methods adopted by Spranghers et al. [
63]. Larval rearing was performed on a growth substrate based on 85% of coffee silverskin and 15% (
w/
w) of spirulina dry biomass, adding distilled water to obtain ~70% of final moisture [
23]. Rearing density was 0.3/cm
2 [
64] and the feeding rate per larva was 100 mg/day [
65]. The growth substrate was replaced once a week until the prepupal stage was reached, identified by the color change of the integument from white to black [
66]. At the end of the rearing phase, prepupae were freeze dried and ground to obtain an insect meal (HM) for the subsequent fish diets formulation. Three complete diets were prepared to cover all the nutrient requirements for rainbow trout [
67]: (i) a control diet (HM0) based on conventional marine (FM and FO) and vegetable ingredients, and (ii) two test diets obtained from the HM0 formulation by replacing 3 (HM3) or 20 (HM20) % of FM and FO with HM (1.8 and 12% levels of dietary inclusion, respectively). All ingredients were ground (0.5 mm) with a Retsch Centrifugal Grinding Mill ZM 1000 (Retsch GmbH, Haan, Germany) and well-mixed with FO to form a homogeneous blend (Kenwood kMix KMX53 stand Mixer). The resulting meshes were added with water (~350 g/kg) and the doughs, thus obtained, were cold extruded into 3 mm pellets using a meat mincer provided with a knife at the die. The wet pellets were then dried in a ventilated oven at 37 °C for 48 h. The diets obtained were stored in sealed bags under vacuum and kept at 4 °C until use. The HM and the experimental diets were analyzed in duplicate for dry matter (DM), total nitrogen (CP) by using the Kjeldahl method, ether extract (EE), and ash content following the AOAC [
68]. The gross energy content (GE) was determined using an adiabatic calorimetric bomb (IKA C7000, Werke GmbH & Co., Staufen, Germany). For the N determination, a nitrogen-to-protein factor of 4.67 was used, as suggested by Janssen et al. [
69], only for the insect ingredient used for diet formulation. In addition, the chitin content of HM and experimental diets was determined according to the method described by Hahn et al. [
70]. The dietary ingredients and proximate composition of both HM and experimental diets are reported in
Table 1.
2.3. Total Lipids, Fatty Acids, Carotenoids and Tocopherol Determination in Fish Diet
Total lipids were isolated as described by Folch et al. [
71]. Minced freeze-dried diets (2 g) were dissolved in 2:1 chloroform to methanol (
v/
v, 40 mL), with added tridecanoic and nonadecanoic acid methyl esters as internal standards (500 μL of a 10 mg/mL solution in
n-hexane), agitated for 5 min, and centrifuged (3000 rpm, 10 min, 4 °C). The organic phase was washed with distilled water (5 mL), filtered through Whatman filter paper (Grade 4, 90 mm, Merck KGaA, Darmstadt, Germany) over anhydrous sodium sulphate (3 g) and evaporated with a rotary evaporator (30 °C) to collect the fat.
Fatty acids methyl esters (FAME) were obtained from total lipids through transmethylation by a BF
3-MeOH reagent [
72]. Briefly, 20 mg of fat were added to
n-hexane (0.5 mL) and a BF
3-MeOH solution (0.5 mL) and vortexed. After 15 min at 100 °C, the reaction was interrupted with distilled water (0.5 mL), and the mixture was centrifuged (4500 rpm, 3 min). Capillary gas chromatography was used to analyze the organic phase as reported by Balzano et al. [
73].
Carotenoids were extracted from freeze-dried samples of diets and analyzed by liquid chromatography as reported by Nartea et al. [
74]. Quantification of carotenoid was performed by external calibration and, in all cases, a correlation coefficient of 0.999 was obtained. For zeaxanthin, the instrumental limit of detection (LOD) and quantification (LOQ) were 5 and 16 ng/mL, respectively, while for β-carotene it was 5 and 18 ng/mL, respectively.
For tocopherol determination, 100 mg of freeze-dried samples were added to 5 mL of hexane, vortexed for 5 minutes, and centrifuged (3500 rpm, 2 min). The organic phase was collected. The extraction was repeated a second time and the organic fractions were pooled together, filtered (0.45 μm, Sartorius Regenerated Cellulose Membrane), dried, resuspended in 0.5 mL hexane, and injected in a Waters Ultra Pressure Liquid Chromatographic Acquity system (UPLC Acquity H-Class, Waters Corporation, Milford, CT, USA) equipped with a Fluorimetric Detector (FLD) and an Ascentis Express Hilic column (15 cm × 2.1 mm, 2.7 mm) [
74,
75]. An isocratic elution of
n-hexane, isopropanol, and acetic acid (95.5, 0.4, 0.1%, respectively) at 0.3 mL/min was performed at a temperature of 30 °C (column heater and sample loading). Tocopherols were detected by comparison of retention time with pure standards and quantified with external calibration. The calibration curves of α-, γ- and δ-tocopherol ranged from 3 to 100 mg/mL with correlation coefficients higher than 0.986. The instrumental LOD and LOQ were 4 and 14 ng/mL for α-tocopherol, 2 and 7 ng/mL for γ-tocopherol, and 2 and 7 ng/mL for δ-tocopherol. Vitamin E content was calculated considering only α-tocopherol as proposed by EFSA NDA Panel.
2.4. Fish Rearing Conditions and Sampling
Six hundred rainbow trout juveniles (15.3 ± 2.2 g) were provided by Itticoltura Valpotenza snc (Fiuminata, MC, Italy) and were maintained for a two-week acclimatization period at “Unità di Ricerca e Didattica di San Benedetto del Tronto (AP, Italy), URDIS-University of Camerino” in 4 m3 tanks with a mechanical and a biological filtration system (Scubla aquaculture snc, Remanzacco, UD, Italy). The temperature was maintained at 16.2 ± 0.2 °C by chillers (Scubla aquaculture snc, Remanzacco, UD, Italy) and pH values were kept constant at 7.88 ± 0.30.
At the beginning of the trial, fish were lightly anesthetized (150 mg/L of MS222; Merck KGaA, Darmstadt, Germany) and individually measured. Five hundred forty fish were distributed into nine fiberglass square tanks (4 m3 each; three tanks per dietary group, 60 fish per tank) that were maintained at the same chemical-physical conditions and were equipped with the same filtration system described for the acclimatization phase. For 6 weeks, fish were fed the experimental diets provided in 1 daily meal in the morning until apparent satiety. All the feed provided was completely ingested by the fish within 30 min after feeding.
After the 6 weeks of trial, fish were sacrificed with a lethal dose of MS222 (1 g/L) and individually measured. Samples of liver, pyloric caeca, distal intestine, and whole fish (residual to the previous samplings) were collected and properly stored for further analyses.
2.5. Growth Performance, Condition Factor, Biometric and Marketable Characteristics
Final survival rate was calculated by removing the number of dead fish from the initial number of specimens. Biometric measurements (final body weight -FBW- and standard length), growth performance [relative growth rate (RGR), specific growth rate (SGR), Fulton’s condition factor (K)], and feed utilization (FCR) were calculated per fish (
n = 60), as follows:
Marketable characteristic [fillet yield (FY), hepatosomatic index (HSI), carcass weight (CW) and viscerosomatic index (VSI)] were calculated per fish (
n = 15), as follows:
2.6. Histological Analysis
Distal intestine and liver samples from 5 fish per tank (
n = 15) were fixed for 24 h at 4 °C in Bouin’s solution and then processed according to Cardinaletti et al. [
13] and Randazzo et al. [
76]. Briefly, samples were washed with ethanol (70%), dehydrated in graded ethanol solutions, washed with xylene, and finally embedded in paraffin (Bio-Optica, Milano, Italy). A Leica RM2125 RTS microtome (Nussloch, Germany) was used to cut sections of 5 µm thickness that were stained with Mayer hematoxylin and eosin Y (Merck KGaA). Histological evaluations were conducted by three operators through independent blind evaluations using a Zeiss Axio Imager.A2 (Zeiss, Oberkochen, Germany) microscope equipped with a combined color digital camera (Axiocam 105, Zeiss). Image analyses were performed using the ZEN 2.3 software (Zeiss).
Distal intestine. For each sample, 6 transversal sections were collected at 200 μm intervals and analyzed to measure mucosal fold height, submucosa width, goblet cells and supranuclear vacuoles according to Randazzo et al. [
77].
Regarding the evaluation of the goblet cell’s relative abundance, cells were counted on 300 μm
2 of absorptive surface on each section. For supranuclear vacuole analysis, an arbitrary index score was assigned as follows: + = scattered, ++ = abundant) [
78].
Liver. Three sections from each sample collected at 200 μm intervals were analyzed to evaluate hepatocyte morphology and the structure of hepatic parenchyma.
2.7. Vibrational Spectroscopic Analyses
Liver sample preparation and spectroscopic measurements using Fourier Transform InfraRed Imaging (FTIRI) analysis were carried out according to the literature [
79,
80,
81,
82]. After sampling, liver samples from three fish per tank (
n = 9) were immediately stored at −80 °C. For each sample, three 10 μm sections were collected at 200 μm intervals using a cryotome. Then, sections were deposited on CaF
2 optical windows (13 mm diameter, 1 mm thick) and then air-dried for 30 min before the acquisition. A Bruker INVENIO-R interferometer coupled with a Hyperion 3000 Vis-IR microscope and equipped with a Focal Plane Array (FPA) detector operating at liquid nitrogen temperature (Bruker Optics, Ettlingen, Germany) was used for FTIRI measurements. For each liver section, a 15X condenser/objective was used to detect specific areas in which the IR maps (164 × 164 μm size; 4096 pixel/spectra with a spatial resolution 2.56 × 2.56 μm) were acquired in the middle infrared range (4000–800 cm
−1) and in transmission mode, performing 256 scans with a spectral resolution of 4 cm
−1. A background spectrum was acquired on a clean portion of the CaF
2 optical window before each acquisition. Raw IR maps were submitted to spectral pre-processing to correct the water vapor and carbon dioxide contributions and to remove thickness variation artifacts on the full frequency range (respectively through Atmospheric Compensation and Vector Normalization routines; OPUS 7.5 software package).
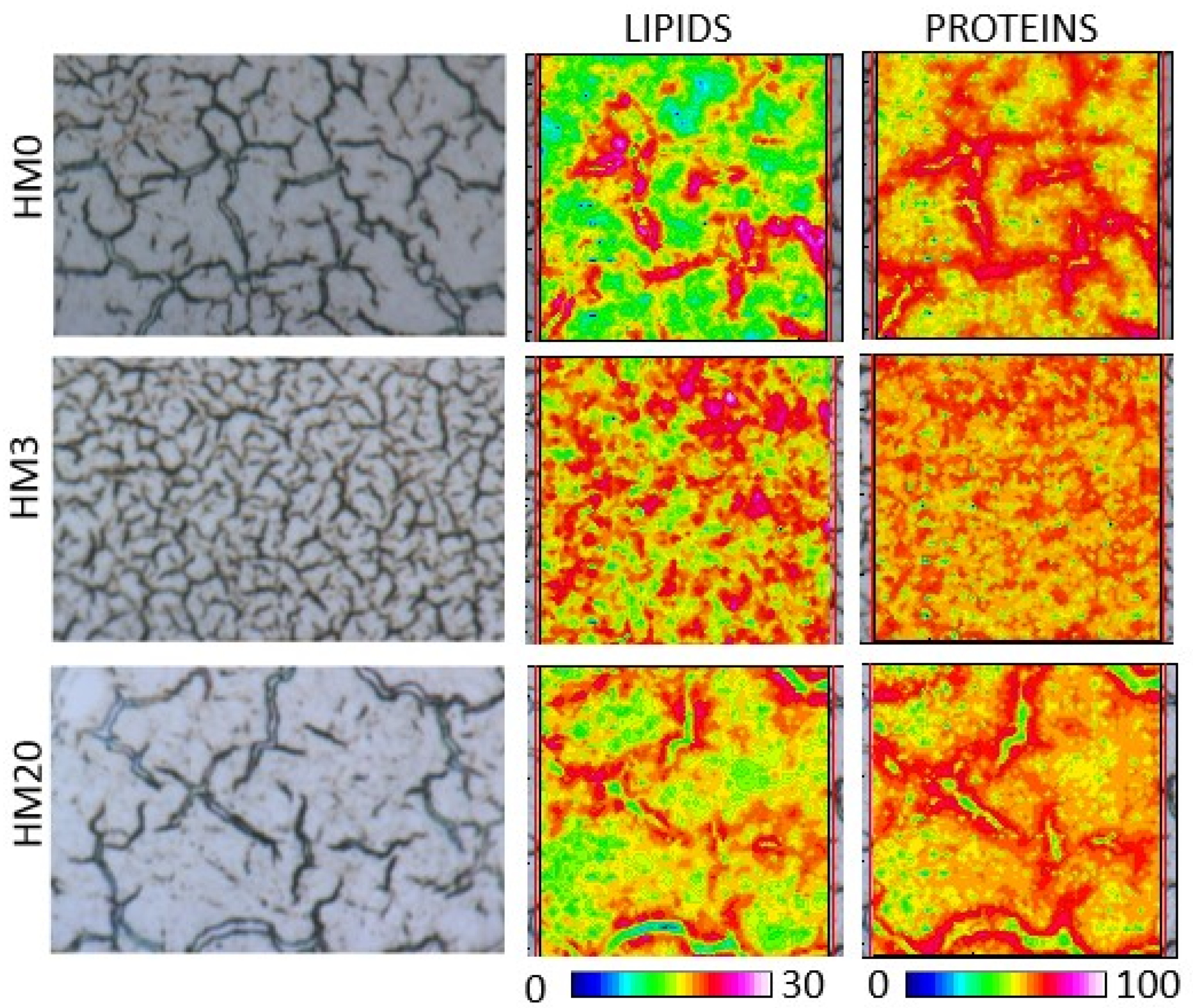
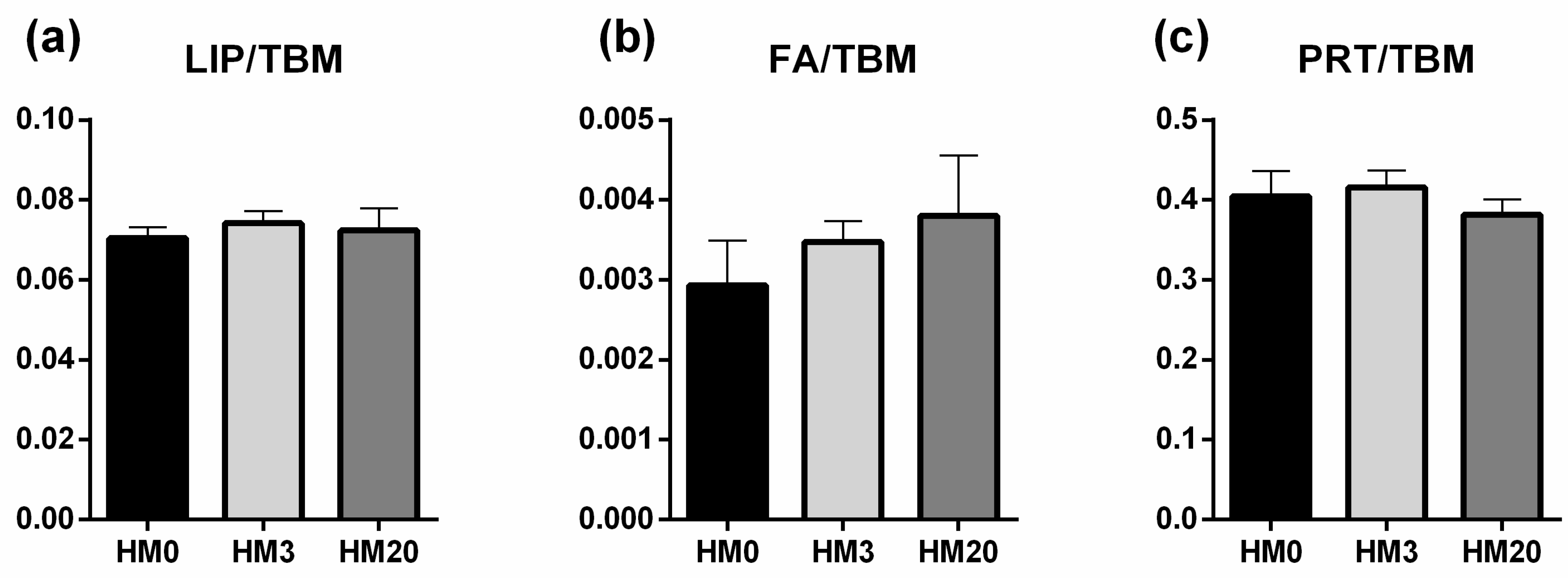
To obtain the false color images, the pre-processed IR maps were integrated in the following spectral ranges: 3000–2800 cm−1 (representative of lipids, LIPIDS), and 1710–1480 cm−1 (representative of proteins, PROTEINS). Then, the band area ratios LIP/TBM (area of the 3050–2800 cm−1 region divided for the sum of the areas of the 3050–2800 cm−1 and 1760–950 cm−1 regions, named TBM), FA/TBM (area of the 1780–1700 cm−1 region, representative of fatty acids, divided for TBM), and PRT/TBM (area of the 1700–1480 cm−1 region divided for TBM) were calculated and statistically analyzed.
2.8. RNA Extraction and cDNA Synthesis
Total RNA was extracted from the liver, distal intestine and pyloric caeca from 5 fish from each tank (
n = 15) using RNAzol RT reagent (Merck KGaA), eluted in 20 µL of RNase-free water (Qiagen), and then stored at −80 °C until use [
83]. A NanoPhotometer P-Class (Implen, München, Germany) was used to determine final the RNA concentration of each sample. RNA integrity was verified by GelRed
TM staining of 28S and 18S ribosomal RNA bands on 1% agarose gel. A cDNA synthesis was performed using the High-Capacity cDNA Reverse Transcription Kit (Bio-Rad, Milano, Italy) from 1 μg of RNA.
2.9. Real-Time PCR
PCR analyses were performed using an iQ5 iCycler thermal cycler (Bio-Rad, Milano, Italy) following the method reported in Maradonna et al. [
84]. For all the reactions, the thermal profile started with 3 min at 95 °C, and then was characterized by 45 cycles of 20 s at 95 °C, 20 s at 60 °C, and 20 s at 72 °C. At the end of each cycle, the fluorescent signal was detected. The melting curve analysis confirmed, for each reaction, the presence of only one PCR amplification product, except for negative controls that did not reveal amplification products and primer-dimer formation. Amplification products were sequenced, and homology was verified.
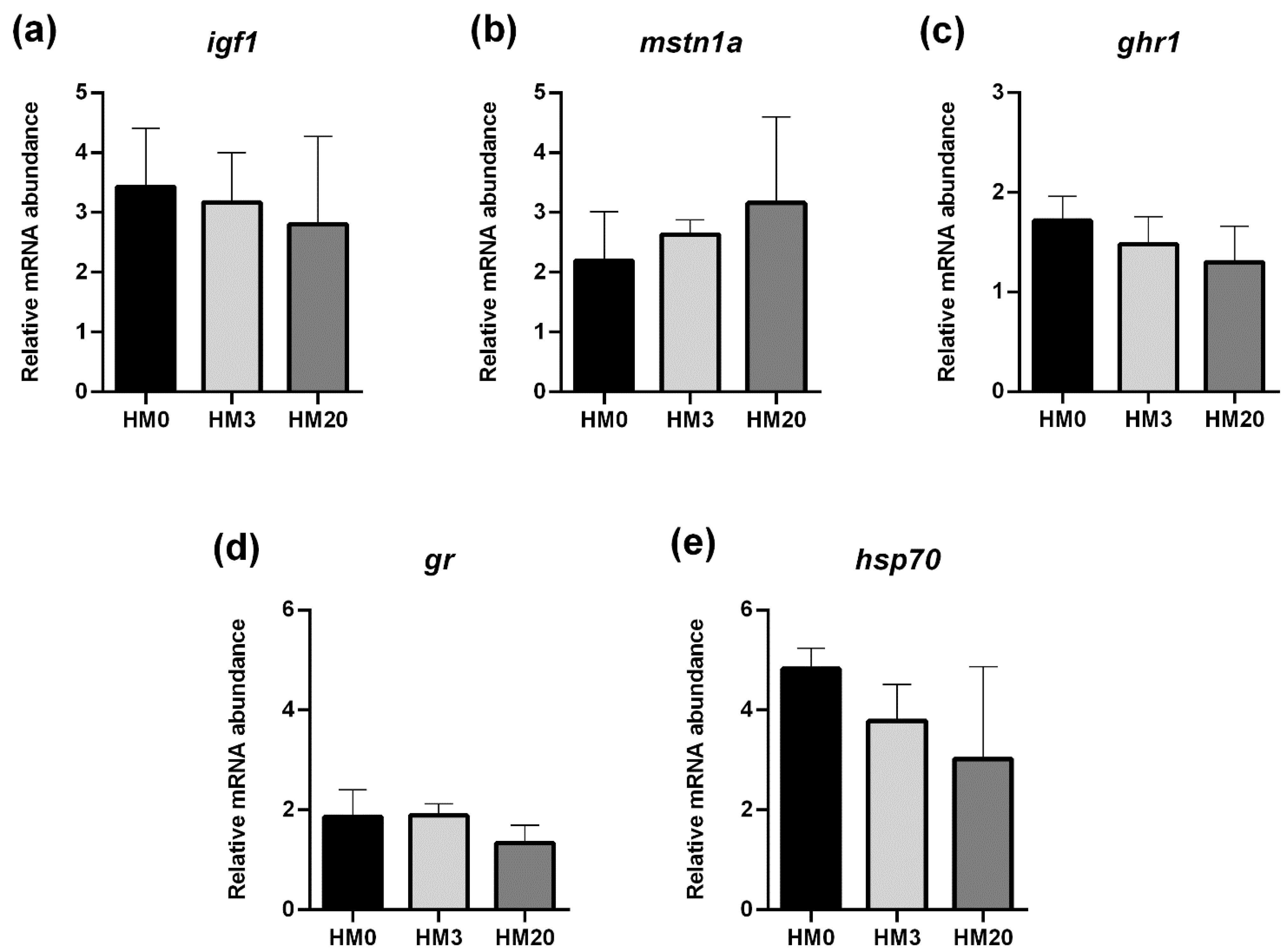
Quantification of relative mRNA abundance was performed in (i) liver samples for genes involved in stress response [glucocorticoid receptor (
gr), heat shock protein 70 (
hsp70)] and fish growth [insulin-like growth factor-1 (
igf1), growth hormone receptor-1 (
ghr1), myostatin1a (
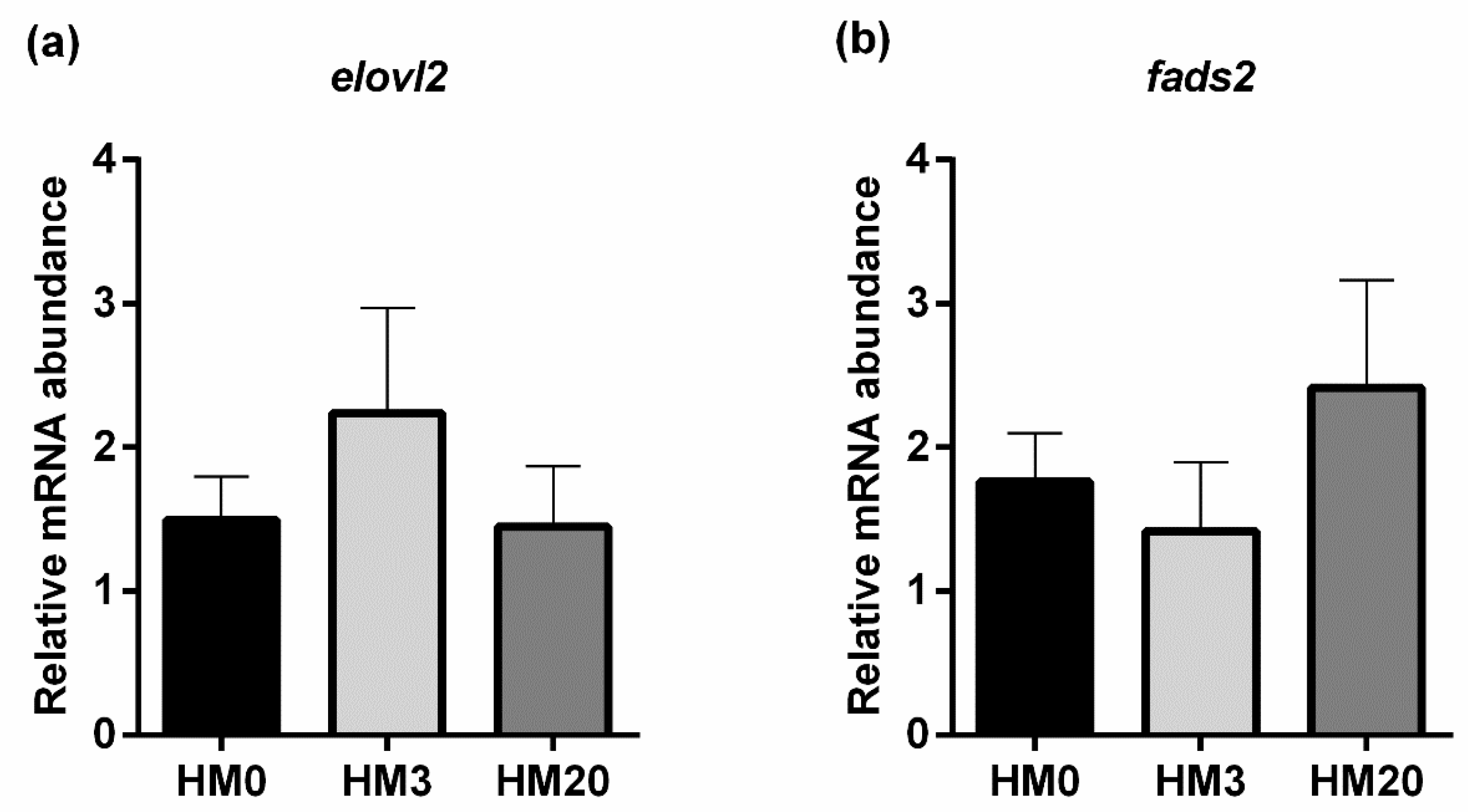
mstn1a)] was performed on liver samples; (ii) pyloric caeca samples for genes involved in PUFA biosynthesis [fatty acid elongase 2 (
elovl2), fatty acid desaturases 2 (
fads2)] (according to Bruni et al. [
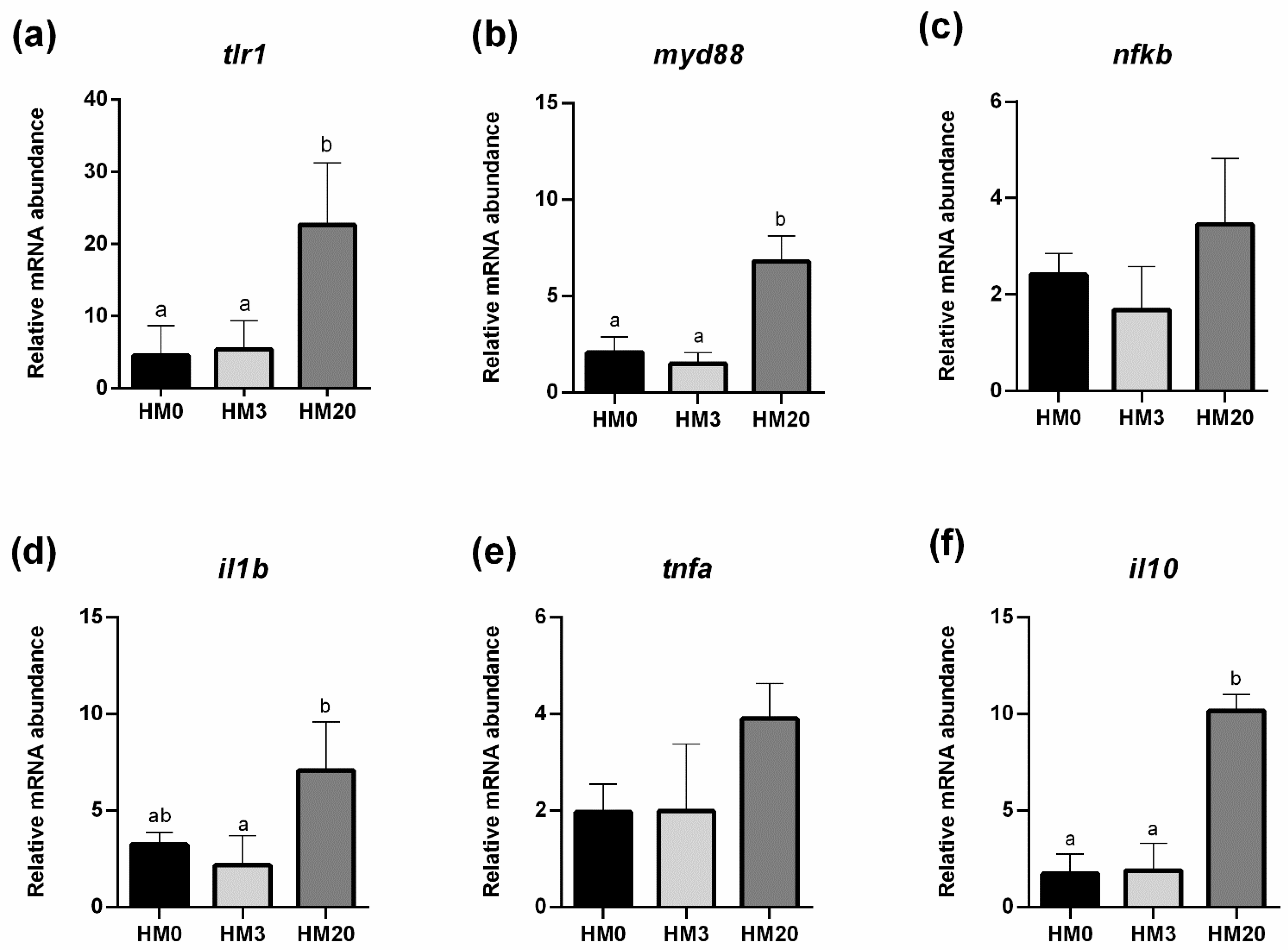
85]); (iii) in distal intestine samples for genes involved in the inflammatory response [toll-like receptor 1 (
tlr1), myeloid differentiation primary response 88 (
myd88), nuclear factor kappa-light-chain-enhancer of activated B cells (
nfkb), interleukin-1β (
il1b), tumor necrosis factor alpha (
tnfa), and interleukin-10 (
il10)]. Both 60S ribosomal and β-actin were used as housekeeping genes. Data obtained were analyzed using the iQ5 optical system software version 2.0 (including GeneEx Macro iQ5 Conversion and GeneEx Macro iQ5 files; Bio-Rad). Primer sequences are reported in
Table 2.
2.10. Physico-Chemical Characterization of Fish Fillets
The color of the fish skin and fillets (
n = 15 per experimental group) were measured on triplicate positions (cranial, medial, and caudal) of the dorsal and of the epaxial region, respectively, and expressed as lightness (
L*), redness index (
a*) and yellowness index (
b*) according to CIE [
87] with the CHROMA METER CR-200 (Konica Minolta, Chiyoda, Japan). Muscle pH was measured also on triplicate positions both in the right and left fillets. A pH-meter SevenGo SG2™ equipped with an Inlab puncture electrode (Mettler-Toledo, Schwerzenbach, Switzerland) was utilized. Then, the fillets were skinned and homogenized to ascertain the Water Holding Capacity (WHC) [
88]. In brief, 2 g from the homogenized fillets were taken and inserted in plexiglass cylinders provided with a filter net; then, they were centrifuged at 1500 rpm (210 g) for 5 min. As a result, WHC was measured as the difference between the initial gross weight of cylinders and their gross weight after centrifugation divided by the fillet water content [
68] and expressed as a percentage.
The total lipids content was obtained following the Folch’s method [
71], gravimetrically quantified to express the result as g of lipid/100 g of fillet, and then suspended into 5 mL of chloroform. A quantity corresponding to 2.5 mg of total lipids were overnight trans-esterified to methyl esters (FAME) in 1% (
v/
v) sulfuric acid in methanol to determine the FA profile composition in the lipid extract [
89]. After the methyl-esterification, the samples were injected into a Varian GC gas chromatograph (Varian Inc., Palo Alto, CA, USA) provided with a flame ionization detector and a Supelco Omegawax™ 320 capillary column (30 m × 0.32 mm × 0.25 μm) (Supelco, Bellefonte, PA, USA). The oven temperature started at 120 °C for 0.5 min and increased up to 170 °C at the rate of 10 °C/min, staying at this temperature for 6 min. Then, it rose to 220 °C at the rate of 3 °C/min and kept stable for 12 min. The duration of the run was 40 min for each sample. The injector and the detector temperatures were set at 220 °C and 300 °C, respectively. Helium was the carrier gas (constant flow of 1.5 mL/min), and the samples were injected in split mode (1:20) with an injection volume of 10 μL. Chromatograms were recorded with the Galaxie Chromatography Data System 1.9.302.952 (Varian Inc., Palo Alto, CA, USA). The FA were identified by comparing the peak retention time of the samples with the retention times of the fatty acid methyl ester standards (C4–C24, 18919-AMP, Sigma-Aldrich
®, St. Louis, MO, USA). Then, they were individually quantified through calibration curves using tricosanoic acid (C23:0) (Sigma-Aldrich, St. Louis, MO, USA) as the internal standard. Data were expressed as a percentage of the total FAME.
2.11. Statistical Analyses
Data from growth parameters and somatic index, histological analysis, FTIR and relative quantification of gene expression were analyzed by one-way ANOVA followed by Tukey’s multiple comparison post hoc test (Prism 8; GraphPad software). Significance was set at p < 0.05. The assumption of normality was checked using the Shapiro–Wilk test. Furthermore, a simple linear regression coefficient was used to evaluate the correlations among fatty acids quantified in the present study.
Data from marketable indexes and physico-chemical analysis of fillets were analyzed by a one-way ANOVA followed by a post hoc Tukey test with the SAS statistical software [
90]. A
p-value of 0.05 was set as the minimum level of significance.
4. Discussion
The physiological responses of rainbow trout to different feed ingredients, including insect meal, have been deeply investigated over the last years [
2,
13,
91,
92,
93]. However, no specific studies have been performed on rainbow trout fed diets containing insect meal obtained from black soldier fly prepupae cultured on a growth substrate enriched with spirulina. In the present study, an increasing content of total PUFA, n-3 PUFA, DHA and EPA from HM0 to HM20 diets was detected. The enrichment procedure of the insects’ biomass thus resulted in a progressive increase in important long-chain PUFA for fish dietary requirements in HM3 and HM20 diets, respectively, highlighting the suitability of this method. In fish, one of the major sites for the synthesis of long-chain PUFA, particularly DHA, is represented by pyloric caeca [
85,
94] that are also involved in the absorption of various lipid components (i.e., free lysophospholipids and monoacylglycerols) [
95]. Two key classes of enzymes are involved in the synthesis of long-chain PUFA in fish: the FA elongase (Elovl) that catalyzes the extension of the FA chain of two carbons and the FA desaturase (Fads) that add a double bond to PUFA substrates [
96]. Usually, elongase and desaturase genes are upregulated in fish when diets with low amounts of long-chain PUFA are provided in order to sustain the biochemical conversion of their precursors [
97], as found by Bruni et al. [
85]. In the present study, the expression of the genes related to fish-lipid metabolism analyzed in pyloric caeca did not show significant differences among the experimental groups, further confirming that all the diets were well balanced in terms of the PUFA profile.
In addition, the FA analyses on fish fillet revealed a comparable amount of n-3 PUFA among the experimental groups and a decreasing SFA content from the HM0 to HM20 groups. In this regard, the enrichment procedure used in the present study, together with the low dietary inclusions of HM, have possibly solved the well-known SFA-related drawback of insect-based diets; this is because, usually, the use of full-fat HM can determine a SFA increase in fish fillet with a parallel decrease of PUFA [
98,
99]. In fact, the addition of spirulina to the insect growth substrate increased the HM PUFA content, which was secondly transferred to the diets during the production activity. This led to comparable fillet LA, ALA, EPA and DHA content among the experimental groups.
As a confirmation of the proper formulation of the diets and their good acceptance by the fish, both the HM dietary inclusion levels did not affect growth performances (as also supported by the gene expression of growth-related markers) and marketable characteristics. These outcomes were expected since most of the literature does not indicate a growth reduction in rainbow trout fed diets including full-fat HM ranging from 10.5 to 20% [
13,
100].
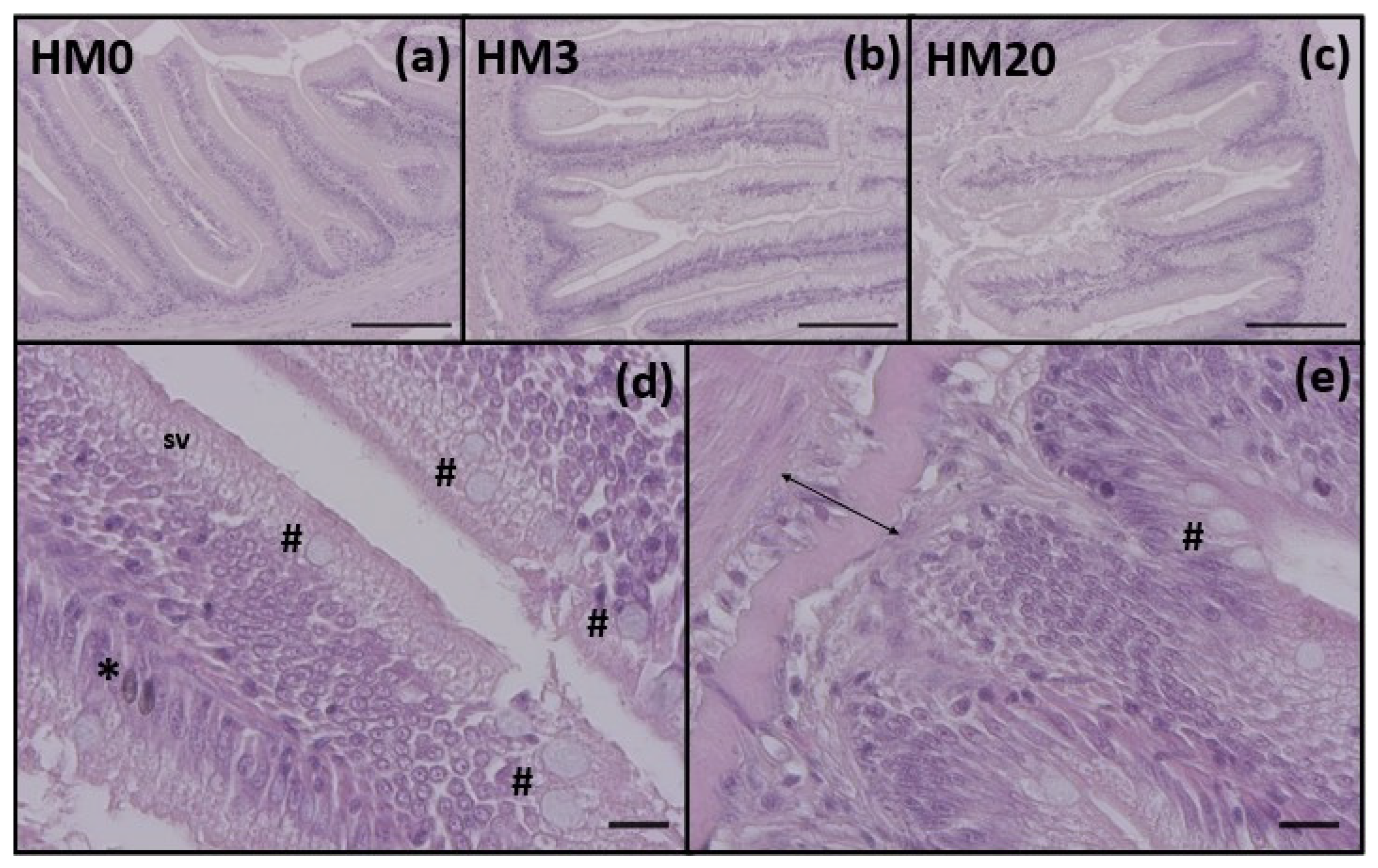
When new ingredients are tested in aquaculture, it is essential to evaluate the gut health and its functional status, especially in terms of the micro-anatomical structure and expression of immune response-related markers. In this regard, histological analyses did not reveal major signs of inflammation and histopathological alterations, despite a significant reduction of the mucosal fold height observed in fish fed an HM20 diet compared to fish fed an HM0 one. Accordingly, a reduction in the absorptive epithelial surface (decrease of intestinal fold height) was observed in rainbow trout fed diets containing 25 or 50% full-fat HM with respect to the control FM-based diet by Cardinaletti et al. [
13]. However, it should be pointed out that, despite the significantly lower value, the mucosal folds height showed by fish fed the HM20 diet was comparable to those observed in healthy rainbow trout analyzed in other studies [
2,
101].
Despite the good results obtained from the histological analyses, the expression of most of the immune response markers analyzed in the present study showed an upregulation in the HM20 group compared to the HM3 and HM0 ones.
This result may be possibly explained by analyzing the role of chitin. Lee at al. [
102] stated that the action of chitin on the mammalian immune system is related to the polymer size. Larger chitin fragments (>70 μm) are inert; conversely, intermediate fragments (70–40 μm) can be recognized by TLR-2 possibly stimulating the TLR-2/MyD-88 signaling pathway with consequent cytokine production [
103], while smaller fragments (<40 μm) can stimulate anti-inflammatory cytokine-like interleukin 10 production [
102]. Additionally, Henry et al. [
104], by analyzing the effects of diets including insect meal on
Dicentrarchus labrax, suggested that the same polyhedric effect of different fragments of chitin is present in fish. TLR-2 works in synergy with TLR-1 [
105], generating a complex that can be involved in the recognition of chitin fragments [
106]. Although chitin fragment size was not measured in the present study, the natural digestion/degradation of chitin through the enzymatic activity could have generated fragments of a certain size responsible for the upregulation of certain immune-related markers observed in the fish fed the HM20 diet.
However, despite the upregulation of inflammatory markers, gut health was preserved in all the experimental groups, and this positive
scenario can be attributed to specific molecules detected in HM diets. In this regard, the HM20 diet presented the highest amount of anti-inflammatory and antioxidant molecules, such as lauric acid (C12:0), tocopherols and carotenoids, with respect to the HM0 and HM3 ones. Specifically, several studies have shown that lauric acid, typically abundant in HM fat fraction, is a powerful anti-inflammatory molecule that can positively influence gut health [
2,
32]. However, the results of the present study highlighted a low amount of lauric acid in the HM20 diet (2.5%) if compared to other similar studies [
107]; thus, the absence of macroscopic intestinal inflammation signs could be mainly related to other molecules such as tocopherols and carotenoids (β-carotene and zeaxanthin) which have been demonstrated to be successfully transferred from spirulina to insect and, secondly, to the feed in the present study.
It is widely known that animal organisms endogenously produce reactive oxygen species (ROS) during inflammatory processes [
108], and the failure of the antioxidant systems to counteract their activities can result in prolonged inflammation, eventually developing the signs of a pathological condition [
109]. Other authors have highlighted that carotenoids operate synergistically with tocopherols, acting as potent radical scavengers, thus preventing oxidative stress and inflammation [
110,
111,
112]. In support to our hypothesis, these previous findings strongly suggest that the absence of inflammation observed in HM20 could be related to the action of these dietary antioxidant molecules.
However, it should be noted that, in addition to their role in cellular metabolism, carotenoids can also be deposited directly within fish chromatophore cells, thus coloring the skin and other tissues [
110]. Carotenoids, together with tocopherols, have been demonstrated to be efficient antioxidants to preserve fish fillet quality, counteracting oxidation during cooking [
75]. In the present study, most of the dietary antioxidant molecules were used by the HM20 fish to avoid the possible occurrence of inflammation in response to alternative ingredients, rather than to be deposited into the flesh as demonstrated by fillet color analysis. In fact, the highest scores for the redness (a*) and the yellowness (b*) indexes were observed in the HM3 group and not in the HM20 group—which, however, was fed the diet richest in these molecules. These data confirm the previous hypothesis that antioxidant molecules possibly exploit their antioxidant activity in preserving gut health and not acting as pigments in fish muscle when the gut is subjected to inflammatory events.
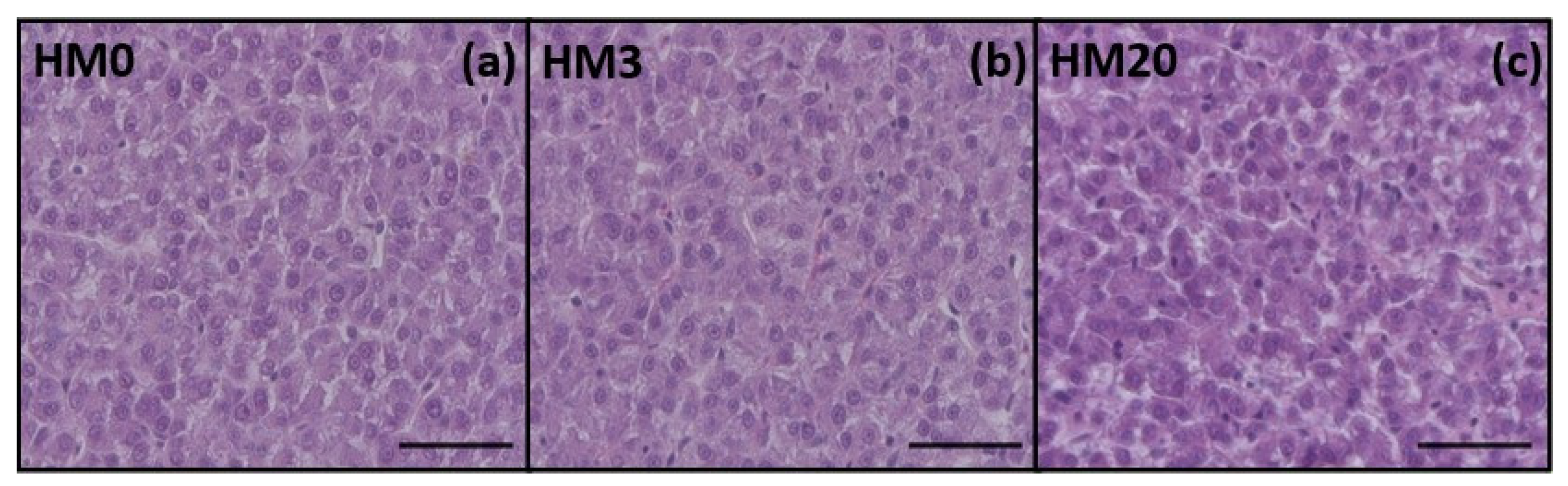
Finally, the liver is another organ that should be considered when new aquafeed ingredients are tested [
113]. Several studies have demonstrated that the use of full-fat HM, at high dietary inclusion levels, can cause a severe steatosis in fish—mainly attributed to a high n-6/n-3 dietary
ratio [
114,
115] and a consequent upregulation of genes involved in stress response [
34]. Fish stress response is extremely important in aquaculture [
116], and in recent years malnutrition has been listed as a possible stressor [
117,
118,
119,
120]. In the present study, no differences in the structure of liver parenchyma were observed among the experimental groups, and FT-IR analyses showed an absence of diversity in terms of total lipids, fatty acids and proteins. These results could be explained by the proper dietary formulation as well as by the n-6/n-3 dietary ratio, which was <1 in all the experimental diets. Consequently,
gr and
hsp70 gene expression, analyzed in the same tissue, did not show differences among the experimental groups—confirming, in accordance with previous studies [
13,
33], the general state of health of the fish.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}