
Longitudinal Survey on aMPV Circulation in French Broiler Flocks following Different Vaccination Strategies

 , , , ,
, , , ,  , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Longitudinal Studies
2.2. aMPV Detection and Sequencing
2.3. aMPV Sequence and Phylogenetic Analysis
2.4. IBV Detection and Sequencing
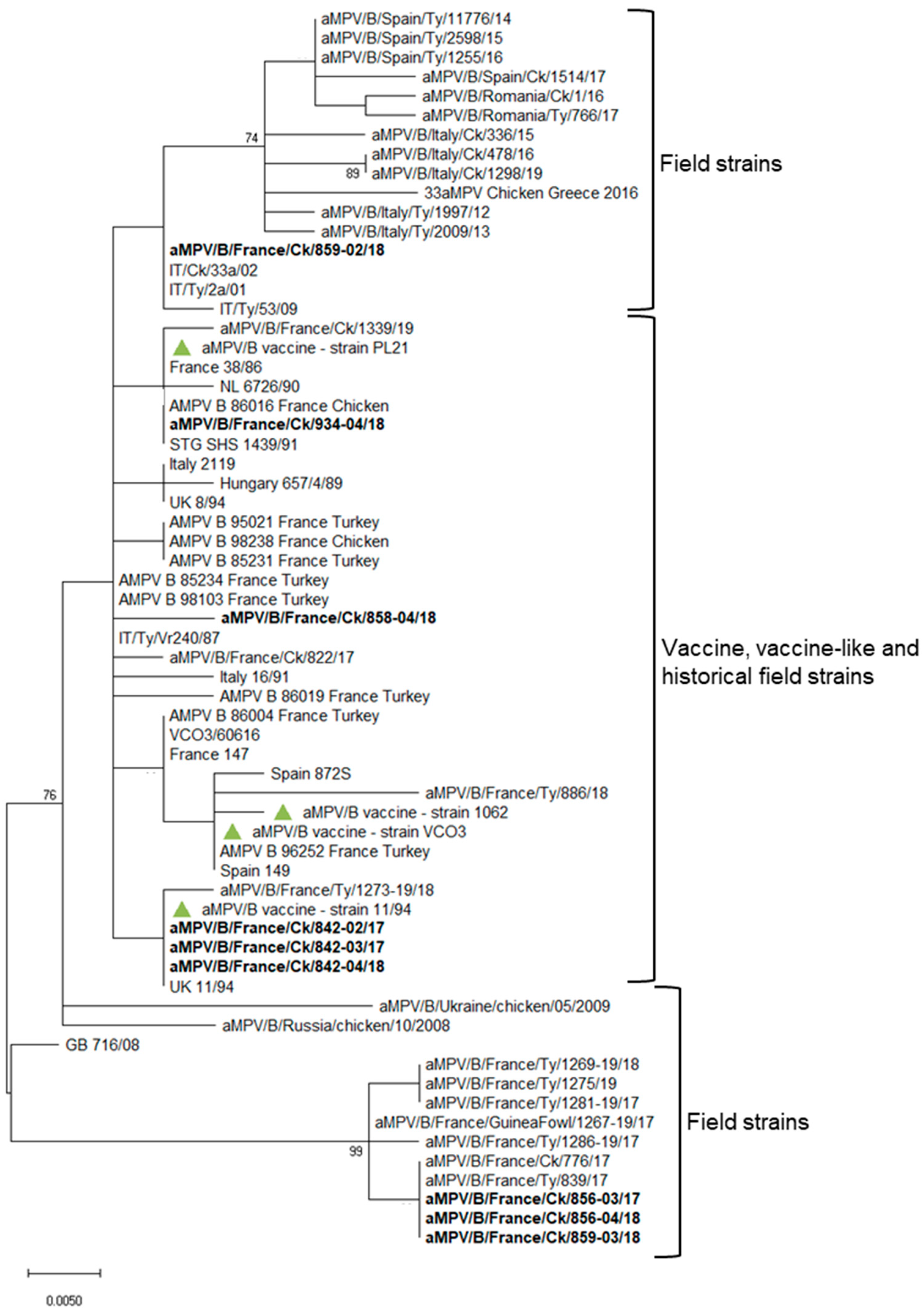
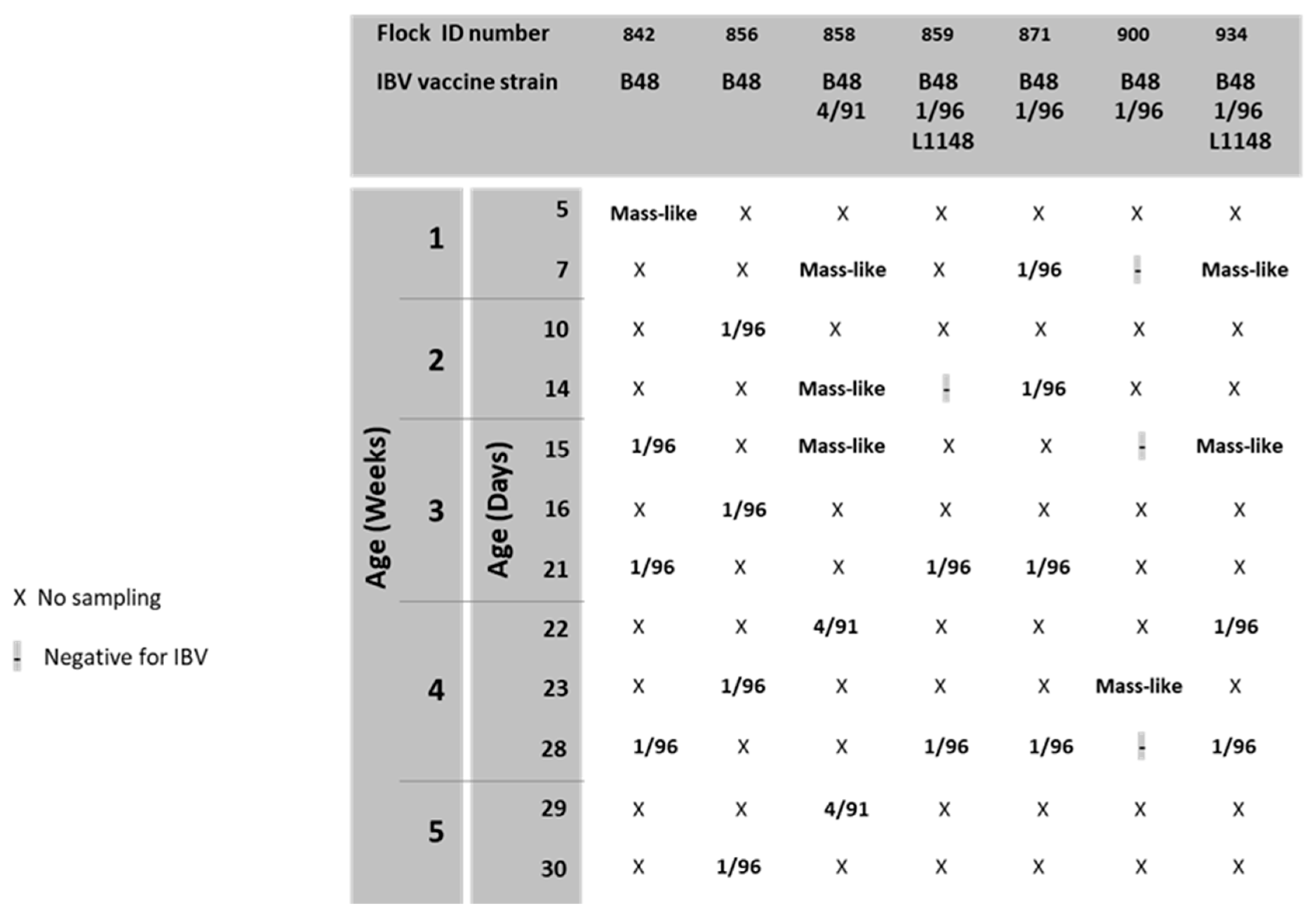
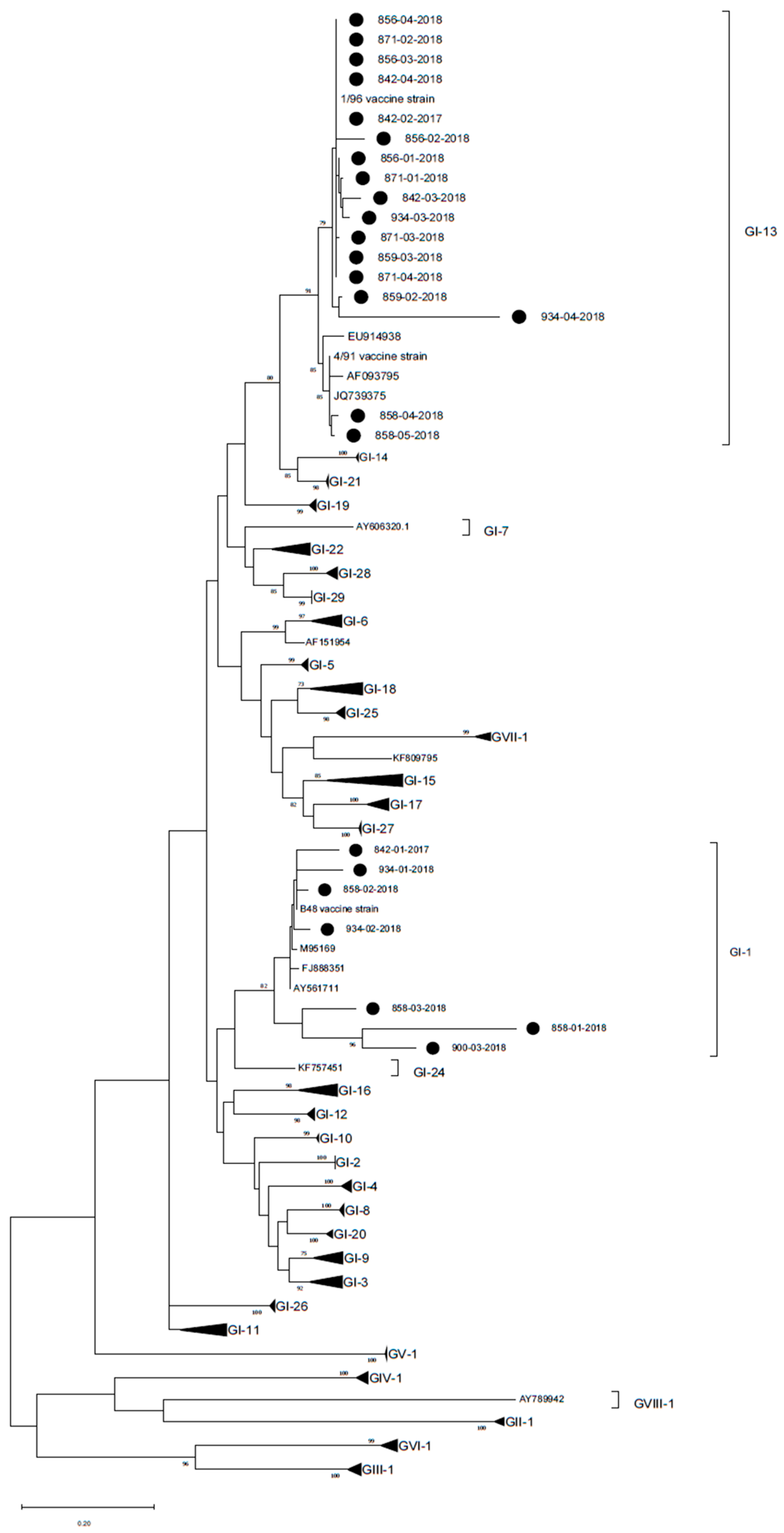
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle Disease, Other Avian Paramyxoviruses, and Avian Metapneumovirus Infections. In Diseases of Poultry; Wiley: Hoboken, NJ, USA, 2020; pp. 109–166. ISBN 9781119371199. [Google Scholar]
- Picault, J.P.; Giraud, P.; Drouin, P.; Guittet, M.; Bennejean, G.; Lamande, J.; Toquin, D.; Gueguen, C. Isolation of a TRTV-like virus from chickens with swollen-head syndrome. Vet. Rec. 1987, 121, 135. [Google Scholar] [CrossRef] [PubMed]
- Toquin, D.; Bäyon-Auboyer, M.H.; Eterradossi, N.; Jestin, V.; Morin, H. Isolation of a pneumovirus from a Muscovy duck. Vet. Rec. 1999, 145, 680. [Google Scholar] [PubMed]
- Catelli, E.; De Marco, M.A.; Delogu, M.; Terregino, C.; Guberti, V. Serological evidence of avian pneumovirus infection in reared and free-living pheasants. Vet. Rec. 2001, 149, 56–58. [Google Scholar] [CrossRef] [PubMed]
- Cecchinato, M.; Lupini, C.; Silveira, F.; Listorti, V.; Mescolini, G.; Morandini, E.; Franzo, G.; Catelli, E. Molecular characterization of avian metapneumovirus from Guinea fowls (Numida meleagridis). Pak. Vet. J. 2018, 38, 419–423. [Google Scholar] [CrossRef]
- Andral, B.; Louzis, C.; Edlinger, E.; Newman, J.A.; Toquin, D.; Bennejean, G. Respiratory disease (rhinotracheitis) in turkeys in Brittany, France, 1981–1982. II. Laboratory findings. Avian Dis. 1985, 29, 35–42. [Google Scholar] [CrossRef]
- Cecchinato, M.; Ferreira, H.L.; Munir, M.; Catelli, E. Avian Metapneumovirus. In Mononegaviruses of Veterinary Importance: Molecular Epidemiology and Control; Munir, M., Ed.; CABI: Wallingford, UK, 2016; Volume 2, pp. 127–143. [Google Scholar]
- Rima, B.; Collins, P.; Easton, A.; Fouchier, R.; Kurath, G.; Lamb, R.A.; Lee, B.; Maisner, A.; Rota, P.; Wang, L. ICTV virus taxonomy profile: Pneumoviridae. J. Gen. Virol. 2017, 98, 2912–2913. [Google Scholar] [CrossRef]
- Bäyon-Auboyer, M.H.; Jestin, V.; Toquin, D.; Cherbonnel, M.; Eterradossi, N. Comparison of F-, G- and N-based RT-PCR protocols with conventional virological procedures for the detection and typing of turkey rhinotracheitis virus. Arch. Virol. 1999, 144, 1091–1109. [Google Scholar] [CrossRef]
- Bayon-Auboyer, M.H.; Arnauld, C.; Toquin, D.; Eterradossi, N. Nucleotide sequences of the F, L and G protein genes of two non-A/non-B avian pneumoviruses (APV) reveal a novel APV subgroup. J. Gen. Virol. 2000, 81, 2723–2733. [Google Scholar] [CrossRef]
- Franzo, G.; Legnardi, M.; Mescolini, G.; Tucciarone, C.M.; Lupini, C.; Quaglia, G.; Catelli, E.; Cecchinato, M. Avian Metapneumovirus subtype B around Europe: A phylodynamic reconstruction. Vet. Res. 2020, 51, 88. [Google Scholar] [CrossRef]
- Toquin, D.; Guionie, O.; Jestin, V.; Zwingelstein, F.; Allee, C.; Eterradossi, N.; Guionie, A.O.; Jestin, A.V.; Zwingelstein, A.F.; Allee, A.C.; et al. European and American subgroup C isolates of avian metapneumovirus belong to different genetic lineages. Virus Genes 2006, 32, 97–103. [Google Scholar] [CrossRef]
- Mescolini, G.; Lupini, C.; Franzo, G.; Quaglia, G.; Legnardi, M.; Cecchinato, M.; Tucciarone, C.M.; Blanco, A.; Turblin, V.; Biarnés, M.; et al. What is new on molecular characteristics of Avian metapneumovirus strains circulating in Europe? Transbound. Emerg. Dis. 2021, 68, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Mawditt, K.; Britton, P.; Naylor, C.J. Longitudinal field studies of infectious bronchitis virus and avian pneumovirus in broilers using type-specific polymerase chain reactions. Avian Pathol. 1999, 28, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Guionie, O.; Toquin, D.; Sellal, E.; Bouley, S.; Zwingelstein, F.; Allée, C.; Bougeard, S.; Lemière, S.; Eterradossi, N. Laboratory evaluation of a quantitative real-time reverse transcription PCR assay for the detection and identification of the four subgroups of avian metapneumovirus. J. Virol. Methods 2007, 139, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Cecchinato, M.; Lupini, C.; Munoz Pogoreltseva, O.S.; Listorti, V.; Mondin, A.; Drigo, M.; Catelli, E. Development of a real-time RT-PCR assay for the simultaneous identification, quantitation and differentiation of avian metapneumovirus subtypes A and B. Avian Pathol. 2013, 42, 283–289. [Google Scholar] [CrossRef]
- Franzo, G.; Drigo, M.; Lupini, C.; Catelli, E.; Laconi, A.; Listorti, V.; Bonci, M.; Naylor, C.J.; Martini, M.; Cecchinato, M. A sensitive, reproducible, and economic real-time reverse transcription PCR detecting avian metapneumovirus subtypes A and B. Avian Dis. 2014, 58, 216–222. [Google Scholar] [CrossRef]
- Cecchinato, M.; Catelli, E.; Lupini, C.; Ricchizzi, E.; Clubbe, J.; Battilani, M.; Naylor, C.J. Avian metapneumovirus (AMPV) attachment protein involvement in probable virus evolution concurrent with mass live vaccine introduction. Vet. Microbiol. 2010, 146, 24–34. [Google Scholar] [CrossRef]
- Listorti, V.; Lupini, C.; Cecchinato, M.; Pesente, P.; Rossi, G.; Giovanardi, D.; Naylor, C.J.; Catelli, E. Rapid detection of subtype B avian metapneumoviruses using RT-PCR restriction endonuclease digestion indicates field circulation of vaccine-derived viruses in older turkeys. Avian Pathol. 2014, 43, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Tucciarone, C.M.; Andreopoulou, M.; Franzo, G.; Prentza, Z.; Chaligiannis, I.; Cecchinato, M. First Identification and Molecular Characterization of Avian metapneumovirus Subtype B from Chickens in Greece. Avian Dis. 2017, 61, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Tucciarone, C.M.; Franzo, G.; Lupini, C.; Alejo, C.T.; Listorti, V.; Mescolini, G.; Brandão, P.E.; Martini, M.; Catelli, E.; Cecchinato, M. Avian Metapneumovirus circulation in Italian broiler farms. Poult. Sci. 2018, 97, 503–509. [Google Scholar] [CrossRef]
- Andreopoulou, M.; Franzo, G.; Tucciarone, C.M.; Prentza, Z.; Koutoulis, K.C.; Cecchinato, M.; Chaligianni, I. Molecular epidemiology of infectious bronchitis virus and avian metapneumovirus in Greece. Poult. Sci. 2019, 98, 5374–5384. [Google Scholar] [CrossRef]
- Franzo, G.; Tucciarone, C.M.; Enache, M.; Bejan, V.; Ramon, G.; Koutoulis, K.C.; Cecchinato, M. First Report of Avian Metapneumovirus Subtype B Field Strain in a Romanian Broiler Flock during an Outbreak of Respiratory Disease. Avian Dis. 2017, 61, 250–254. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.J.; De Wit, M.K.; Cook, J.K.A. Infectious Bronchitis Virus Types Affecting European Countries—A Review. Avian Dis. 2021, 65, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.K.A.; Huggins, M.B.; Orbell, S.J.; Mawditt, K.; Cavanagh, D. Infectious bronchitis virus vaccine interferes with the replication of avian pneumovirus vaccine in domestic fowl. Avian Pathol. 2001, 30, 233–242. [Google Scholar] [CrossRef]
- Tucciarone, C.M.; Franzo, G.; Berto, G.; Drigo, M.; Ramon, G.; Koutoulis, K.C.; Catelli, E.; Cecchinato, M. Evaluation of 793/B-like and Mass-like vaccine strain kinetics in experimental and field conditions by real-Time RT-PCR quantification. Poult. Sci. 2018, 97, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.M.; Roe, R.T. Livestock Disease Surveys: A Field Manual for Veterinarians; Australian Government Publishing Service: Canberra, Australia, 1982. [Google Scholar]
- Felice, V.; Lupini, C.; Mescolini, G.; Silveira, F.; Guerrini, A.; Catelli, E.; Di Francesco, A. Molecular detection and characterization of Mycoplasma gallisepticum and Mycoplasma synoviae strains in backyard poultry in Italy. Poult. Sci. 2020, 99, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, K.; Catelli, E.; Lemiere, S.; Montiel, E.; Jones, R.C. Protection conferred by a live avian metapneumovirus vaccine when co-administered with live La Sota Newcastle disease vaccine in chicks. Ital. J. Anim. Sci. 2014, 13, 404–409. [Google Scholar] [CrossRef] [Green Version]
- De Marco, M.A.; Delogu, M.; Facchini, M.; Di Trani, L.; Boni, A.; Cotti, C.; Graziosi, G.; Venturini, D.; Regazzi, D.; Ravaioli, V.; et al. Serologic evidence of occupational exposure to avian influenza viruses at the wildfowl/poultry/human interface. Microorganisms 2021, 9, 2153. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Worthington, K.J.; Currie, R.J.W.; Jones, R.C. A reverse transcriptase-polymerase chain reaction survey of infectious bronchitis virus genotypes in Western Europe from 2002 to 2006. Avian Pathol. 2008, 37, 247–257. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, E.; Umar, S.; Yilmaz, A.; Turan, N.; Franzo, G.; Tucciarone, C.M.; Cecchinato, M.; Cakan, B.; Iqbal, M.; Yilmaz, H. First Molecular Characterization of Avian Metapneumovirus (aMPV) in Turkish Broiler Flocks. Avian Dis. 2018, 62, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Banet-Noach, C.; Simanov, L.; Perk, S. Characterization of Israeli avian metapneumovirus strains in turkeys and chickens. Avian Pathol. 2005, 34, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Owoade, A.A.; Ducatez, M.F.; Hübschen, J.M.; Sausy, A.; Chen, H.; Guan, Y.; Muller, C.P. Avian metapneumovirus subtype A in China and subtypes A and B in Nigeria. Avian Dis. 2008, 52, 502–506. [Google Scholar] [CrossRef]
- Chacón, J.L.; Mizuma, M.; Vejarano, M.P.; Toquín, D.; Eterradossi, N.; Patnayak, D.P.; Goyal, S.M.; Piantino Ferreira, A.J. Avian metapneumovirus subtypes circulating in Brazilian vaccinated and nonvaccinated chicken and turkey farms. Avian Dis. 2011, 55, 82–89. [Google Scholar] [CrossRef]
- Brown, P.A.; Allée, C.; Courtillon, C.; Szerman, N.; Lemaitre, E.; Toquin, D.; Mangart, J.-M.M.; Amelot, M.; Eterradossi, N. Host specificity of avian metapneumoviruses. Avian Pathol. 2019, 48, 311–318. [Google Scholar] [CrossRef]
- Wei, L.; Zhu, S.; Yan, X.; Wang, J.; Zhang, C.; Liu, S.; She, R.; Hu, F.; Quan, R.; Liu, J. Avian metapneumovirus subgroup C infection in chickens, China. Emerg. Infect. Dis. 2013, 19, 1092–1094. [Google Scholar] [CrossRef]
- Turpin, E.A.; Stallknecht, D.E.; Slemons, R.D.; Zsak, L.; Swayne, D.E. Evidence of avian metapneumovirus subtype C infection of wild birds in Georgia, South Carolina, Arkansas and Ohio, USA. Avian Pathol. 2008, 37, 343–351. [Google Scholar] [CrossRef]
- Sun, S.; Chen, F.; Cao, S.; Liu, J.; Lei, W.; Li, G.; Song, Y.; Lu, J.; Liu, C.; Qin, J.; et al. Isolation and characterization of a subtype C avian metapneumovirus circulating in Muscovy ducks in China. Vet. Res. 2014, 45, 74. [Google Scholar] [CrossRef]
- Mi, R.; Yu, Q.; Zsak, L. The pathogenicity of avian metapneumovirus subtype C wild bird isolates in domestic turkeys. Virol. J. 2013, 10, 38. [Google Scholar] [CrossRef]
- Graziosi, G.; Mescolini, G.; Silveira, F.; Lupini, C.; Tucciarone, C.M.; Franzo, G.; Cecchinato, M.; Legnardi, M.; Gobbo, F.; Terregino, C.; et al. First detection of avian metapneumovirus subtype C Eurasian lineage in a Eurasian wigeon (Mareca penelope) wintering in Northeastern Italy: An additional hint on the role of migrating birds in the viral epidemiology. Avian Pathol. 2022, 51, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Legnardi, M.; Allée, C.; Franzo, G.; Cecchinato, M.; Brown, P. Research Note: Detection of Avian metapneumovirus subgroup C specific antibodies in a mallard flock in Italy. Poult. Sci. 2021, 100, 101186. [Google Scholar] [CrossRef] [PubMed]
- Tucciarone, C.M.; Franzo, G.; Legnardi, M.; Pasotto, D.; Lupini, C.; Catelli, E.; Quaglia, G.; Graziosi, G.; Dal Molin, E.; Gobbo, F.; et al. Molecular Survey on A, B, C and New Avian Metapneumovirus (aMPV) Subtypes in Wild Birds of Northern-Central Italy. Vet. Sci. 2022, 9, 373. [Google Scholar] [CrossRef]
- Graziosi, G.; Lupini, C.; Catelli, E. Disentangling the role of wild birds in avian metapneumovirus (aMPV) epidemiology: A systematic review and meta-analysis. Transbound. Emerg. Dis. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Jesse, S.T.; RibóMolina, P.; Jo, W.K.; Rautenschlein, S.; Vuong, O.; Fouchier, R.A.M.; Ludlow, M.; Osterhaus, A.D.M.E. Molecular characterization of avian metapneumovirus subtype C detected in wild mallards (Anas platyrhynchos) in The Netherlands. Transbound. Emerg. Dis. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Catelli, E.; Cecchinato, M.; Savage, C.E.; Jones, R.C.; Naylor, C.J. Demonstration of loss of attenuation and extended field persistence of a live avian metapneumovirus vaccine. Vaccine 2006, 24, 6476–6482. [Google Scholar] [CrossRef] [PubMed]
- Cecchinato, M.; Catelli, E.; Lupini, C.; Ricchizzi, E.; Prosperi, S.; Naylor, C.J. Reversion to virulence of a subtype B avian metapneumovirus vaccine: Is it time for regulators to require availability of vaccine progenitors? Vaccine 2014, 32, 4660–4664. [Google Scholar] [CrossRef] [PubMed]
- Legnardi, M.; Franzo, G.; Koutoulis, K.C.; Wiśniewski, M.; Catelli, E.; Tucciarone, C.M.; Cecchinato, M. Vaccine or field strains: The jigsaw pattern of infectious bronchitis virus molecular epidemiology in Poland. Poult. Sci. 2019, 98, 6388–6392. [Google Scholar] [CrossRef]
- Legnardi, M.; Cecchinato, M.; Homonnay, Z.; Dauphin, G.; Koutoulis, K.C.; Tucciarone, C.M.; Franzo, G. Viral subpopulation variability in different batches of Infectious bronchitis virus (IBV) vaccines based on GI-23 lineage: Implications for the field. Virus Res. 2022, 319, 198877. [Google Scholar] [CrossRef]
- Legnardi, M.; Tucciarone, C.M.; Franzo, G.; Cecchinato, M. Infectious Bronchitis Virus Evolution, Diagnosis and Control. Vet. Sci. 2020, 7, 79. [Google Scholar] [CrossRef]
- Tucciarone, C.M.; Franzo, G.; Bianco, A.; Berto, G.; Ramon, G.; Paulet, P.; Koutoulis, K.C.; Cecchinato, M. Infectious bronchitis virus gel vaccination: Evaluation of Mass-like (B-48) and 793/B-like (1/96) vaccine kinetics after combined administration at 1 day of age. Poult. Sci. 2018, 97, 3501–3509. [Google Scholar] [CrossRef] [PubMed]
- Tucciarone, C.M.; Franzo, G.; Legnardi, M.; Fortin, A.; Valastro, V.; Lazzaro, E.; Terregino, C.; Cecchinato, M. Effect of assay choice, viral concentration and operator interpretation on infectious bronchitis virus detection and characterization. Avian Pathol. 2021, 50, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Naylor, C.J.; Drigo, M.; Croville, G.; Ducatez, M.F.; Catelli, E.; Laconi, A.; Cecchinato, M. Subpopulations in aMPV vaccines are unlikely to be the only cause of reversion to virulence. Vaccine 2015, 33, 2438–2441. [Google Scholar] [CrossRef] [PubMed]
- Naylor, C.J.; Ling, R.; Edworthy, N.; Savage, C.E.; Easton, A.J. Avian metapneumovirus SH gene end and G protein mutations influence the level of protection of live-vaccine candidates. J. Gen. Virol. 2007, 88, 1767–1775. [Google Scholar] [CrossRef]
- Cook, J.K.A.; Ellis, M.M.; Dolby, C.A.; Huggins, M.B.; Holmes, H.C.; Finney, P.M. A Live Attenuated Turkey Rhinotracheitis Virus Vaccine. 1. Stability of the Attenuated Strain. Avian Pathol. 1989, 18, 511–522. [Google Scholar] [CrossRef]
- Giraud, P.; Bennejean, G.; Guittet, M.; Toquin, D. Turkey rhinotracheitis in France: Preliminary investigations on a ciliostatic virus. Vet. Rec. 1986, 119, 606–607. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flock ID N° | aMPV Vaccine Strain—Age | IBV Vaccine Strain—Age |
|---|---|---|
| 842 | 11/94—1 day | B48—day of hatch |
| 856 | PL21—1 day | B48—day of hatch |
| 858 | PL21—day of hatch | B48—day of hatch 4/91—14 days |
| 859 | None—NA | B48 + 1/96—day of hatch L1148—13 days |
| 871 | None—NA | B48 + 1/96—1 day |
| 900 | PL21—7 days | B48 + 1/96—day of hatch |
| 934 | PL21—1 day | B48 + 1/96—day of hatch L1148—14 days |
| Strain Name | Year of Isolation |
|---|---|
| AMPV B 85231 France Turkey | 1985 |
| AMPV B 85234 France Turkey | 1985 |
| AMPV B 86004 France Turkey | 1986 |
| AMPV B 86016 France Chicken | 1986 |
| AMPV B 86019 France Turkey | 1986 |
| AMPV B 95021 France Turkey | 1995 |
| AMPV B 96252 France Turkey | 1996 |
| AMPV B 98103 France Turkey | 1998 |
| AMPV B 98238 France Chicken | 1998 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupini, C.; Tucciarone, C.M.; Mescolini, G.; Quaglia, G.; Graziosi, G.; Turblin, V.; Brown, P.; Cecchinato, M.; Legnardi, M.; Delquigny, T.; et al. Longitudinal Survey on aMPV Circulation in French Broiler Flocks following Different Vaccination Strategies. Animals 2023, 13, 57. https://doi.org/10.3390/ani13010057
Lupini C, Tucciarone CM, Mescolini G, Quaglia G, Graziosi G, Turblin V, Brown P, Cecchinato M, Legnardi M, Delquigny T, et al. Longitudinal Survey on aMPV Circulation in French Broiler Flocks following Different Vaccination Strategies. Animals. 2023; 13(1):57. https://doi.org/10.3390/ani13010057
Chicago/Turabian StyleLupini, Caterina, Claudia Maria Tucciarone, Giulia Mescolini, Giulia Quaglia, Giulia Graziosi, Vincent Turblin, Paul Brown, Mattia Cecchinato, Matteo Legnardi, Thomas Delquigny, and et al. 2023. "Longitudinal Survey on aMPV Circulation in French Broiler Flocks following Different Vaccination Strategies" Animals 13, no. 1: 57. https://doi.org/10.3390/ani13010057
APA StyleLupini, C., Tucciarone, C. M., Mescolini, G., Quaglia, G., Graziosi, G., Turblin, V., Brown, P., Cecchinato, M., Legnardi, M., Delquigny, T., Lemiere, S., Perreul, G., & Catelli, E. (2023). Longitudinal Survey on aMPV Circulation in French Broiler Flocks following Different Vaccination Strategies. Animals, 13(1), 57. https://doi.org/10.3390/ani13010057