
Heat Stress: A Serious Disruptor of the Reproductive Physiology of Dairy Cows

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
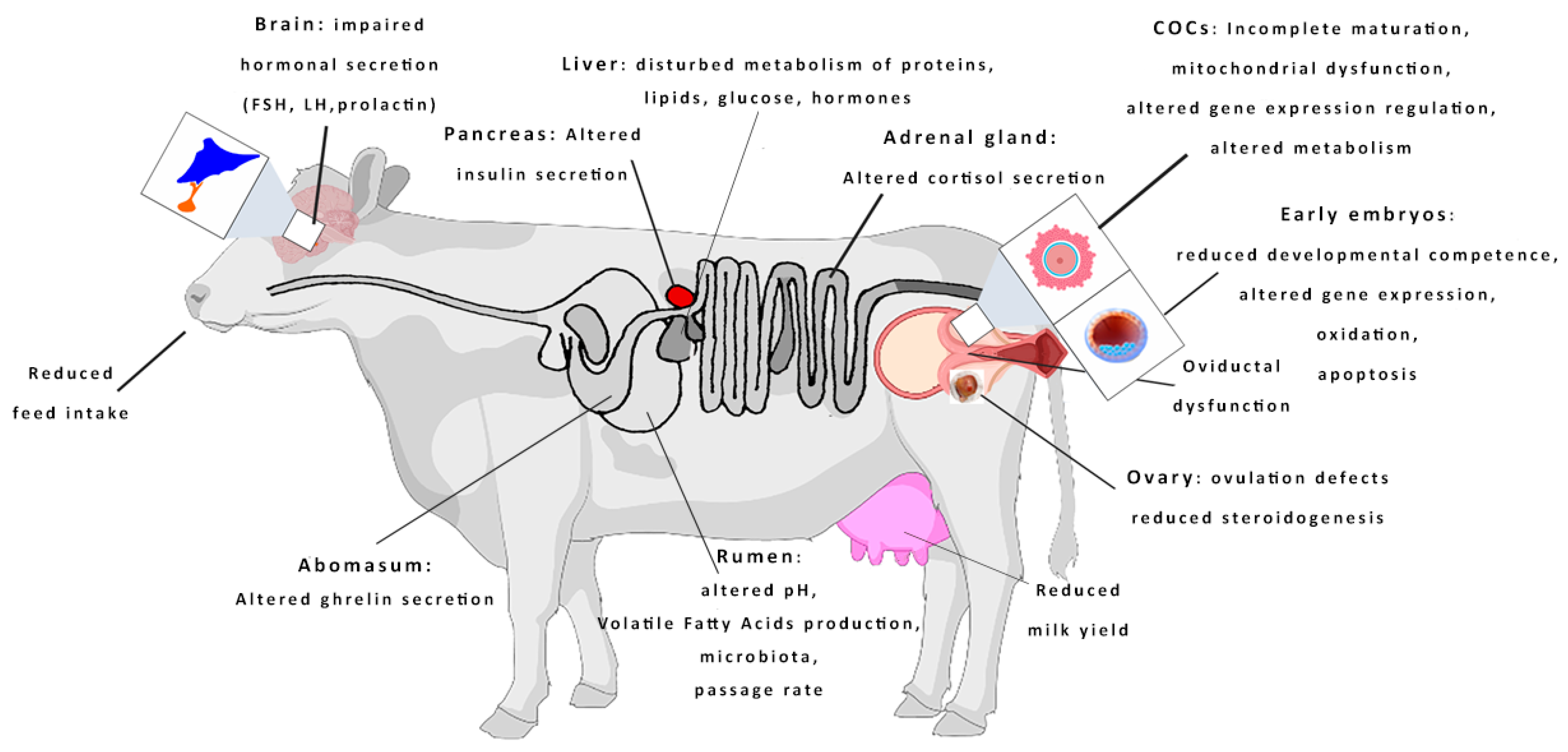
1. Introduction
2. Heat Shock Response: The First Line of Molecular Defense Mechanisms against Heat Stress
3. Endocrine Response to Heat Stress
3.1. Reproductive Endocrinology under Heat Stress
3.2. LH
3.3. FSH
3.4. Estradiol
3.5. Progesterone
3.6. Cortisol
3.7. Prolactin
3.8. Follicular Development
4. Cumulus Complexes, Oocyte Complexes, and the Early Embryo
4.1. The Oocyte and the Surrounding Cumulus Cells (COCs)
4.2. Early Embryos
5. Oviduct—Gravid Uterus
6. Conclusions and Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, P.R.; Raftery, A.E. Country-based rate of emissions reductions should increase by 80% beyond nationally determined contributions to meet the 2 °C target. Commun. Earth Environ. 2021, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar]
- Crowley, T.J.; Lowery, T.S. How warm was the medieval warm period? AMBIO A J. Hum. Environ. 2000, 29, 51–54. [Google Scholar] [CrossRef]
- Jacobsen, K.L. The well-being of dairy cows in hot and humid climates. Part 1. Housing and effects of heat stress. Comp. Contin. Educ. Pract. Vet. 1996, 18, S59–S63. [Google Scholar]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Roth, Z. Impact of heat stress on cow reproduction and fertility. Anim. Front. 2019, 9, 32–38. [Google Scholar] [CrossRef]
- Collier, R.J.; Baumgard, L.H.; Zimbelman, R.B.; Xiao, Y. Heat stress: Physiology of acclimation and adaptation. Anim. Front. Rev. Mag. Anim. Agric. 2019, 9, 12–19. [Google Scholar] [CrossRef]
- Kendall, P.E.; Nielsen, P.P.; Webster, J.R.; Verkerk, G.A.; Littlejohn, R.P.; Matthews, L.R. The effects of providing shade to lactating dairy cows in a temperate climate. Livest. Sci. 2006, 103, 148–157. [Google Scholar] [CrossRef]
- Schütz, K.E.; Cox, N.R.; Matthews, L.R. How important is shade to dairy cattle? Choice between shade or lying following different levels of lying deprivation. Appl. Anim. Behav. Sci. 2008, 114, 307–318. [Google Scholar] [CrossRef]
- McDowell, R.E. Water exchange of cattle under heat stress. Biometeorology 1967, 2, 414. [Google Scholar]
- Kadzere, C.T.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Armstrong, D. Heat stress interaction with shade and cooling. J. Dairy Sci. 1994, 77, 2044–2050. [Google Scholar] [CrossRef]
- Pegorer, M.F.; Vasconcelos, J.L.M.; Trinca, L.A.; Hansen, P.J.; Barros, C.M. Influence of sire and sire breed (Gyr versus Holstein) on establishment of pregnancy and embryonic loss in lactating Holstein cows during summer heat stress. Theriogenology 2007, 67, 692–697. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef]
- Wheelock, J.B.; Rhoads, R.P.; Vanbaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Krishnan, G.; Bagath, M.; Pragna, P.; Vidya, M.K.; Aleena, J.; Archana, P.R.; Sejian, V.; Bhatta, R. Mitigation of the Heat Stress Impact in Livestock Reproduction; Carreira, R.P., Ed.; IntechOpen: Rijeka, Croatia, 2017; p. Ch. 4. ISBN 978-953-51-3478-7. [Google Scholar]
- Mishra, S.R. Behavioural, physiological, neuro-endocrine and molecular responses of cattle against heat stress: An updated review. Trop. Anim. Health Prod. 2021, 53, 400. [Google Scholar] [CrossRef]
- Stamperna, K.; Dovolou, E.; Giannoulis, T.; Kalemkeridou, M.; Nanas, I.; Dadouli, K.; Moutou, K.; Mamuris, Z.; Amiridis, G.S. Developmental competence of heat stressed oocytes from Holstein and Limousine cows matured in vitro. Reprod. Domest. Anim. 2021, 56, 1302–1314. [Google Scholar] [CrossRef]
- Stamperna, K.; Giannoulis, T.; Cañon-Beltrán, K.; Dovolou, E.; Kalemkeridou, M.; Nanas, I.; Rizos, D.; Moutou, K.A.; Mamuris, Z.; Amiridis, G.S. Oviductal epithelial cells transcriptome and extracellular vesicles characterization during thermoneutral and heat stress conditions in dairy cows. Theriogenology 2022, 187, 152–163. [Google Scholar] [CrossRef]
- Nanas, I.; Chouzouris, T.; Dovolou, E.; Dadouli, K.; Stamperna, K.; Kateri, I.; Barbagianni, M.; Amiridis, G.S. Early embryo losses, progesterone and pregnancy associated glycoproteins levels during summer heat stress in dairy cows. J. Therm. Biol. 2021, 98, 102951. [Google Scholar] [CrossRef]
- Page, T.J.; Sikder, D.; Yang, L.; Pluta, L.; Wolfinger, R.D.; Kodadek, T.; Thomas, R.S. Genome-wide analysis of human HSF1 signaling reveals a transcriptional program linked to cellular adaptation and survival. Mol. Biosyst. 2006, 2, 627–639. [Google Scholar] [CrossRef]
- Beere, H.M.; Green, D.R. Stress management—Heat shock protein-70 and the regulation of apoptosis. Trends Cell Biol. 2001, 11, 6–10. [Google Scholar] [CrossRef]
- Borges, T.J.; Wieten, L.; Van Herwijnen, M.J.C.; Broere, F.; Van derZee, R.; Bonorino, C.; Van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. I Nterleukin-10 and the I Nterleukin-10 R Eceptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Wegele, H.; Müller, L.; Buchner, J. Hsp70 and Hsp90—A relay team for protein folding. In Reviews of Physiology, Biochemistry and Pharmacology. Reviews of Physiology, Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 2004; Volume 151. [Google Scholar] [CrossRef]
- Ju, J. Cellular responses of oocytes and embryos under thermal stress: Hints to molecular signaling. Anim. Reprod 2018, 2, 79–90. [Google Scholar]
- Ashburner, M. Patterns of puffing activity in the salivary gland chromosomes of Drosophila—V. Responses to environmental treatments. Chromosoma 1970, 31, 356–376. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-S.; Nejad, J.G.; Roh, S.-G.; Lee, H.-G. Heat-Shock Proteins Gene Expression in Peripheral Blood Mononuclear Cells as an Indicator of Heat Stress in Beef Calves. Animals 2020, 10, 895. [Google Scholar] [CrossRef]
- Khan, A.; Dou, J.; Wang, Y.; Jiang, X.; Khan, M.Z.; Luo, H.; Usman, T.; Zhu, H. Evaluation of heat stress effects on cellular and transcriptional adaptation of bovine granulosa cells. J. Anim. Sci. Biotechnol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Kumar, A.; Ashraf, S.; Goud, T.S.; Grewal, A.; Singh, S.V.; Yadav, B.R.; Upadhyay, R.C. Expression profiling of major heat shock protein genes during different seasons in cattle (Bos indicus) and buffalo (Bubalus bubalis) under tropical climatic condition. J. Therm. Biol. 2015, 51, 55–64. [Google Scholar] [CrossRef]
- Nanas, I.; Chouzouris, T.M.; Dadouli, K.; Dovolou, E.; Stamperna, K.; Barbagianni, M.; Valasi, I.; Tsiaras, A.; Amiridis, G.S. A study on stress response and fertility parameters in phenotypically thermotolerant and thermosensitive dairy cows during summer heat stress. Reprod. Domest. Anim. 2020, 55, 1774–1783. [Google Scholar] [CrossRef]
- Archana, P.R. Role of Heat Shock Proteins in Livestock Adaptation to Heat Stress. J. Dairy Vet. Anim. Res. 2017, 5, 00127. [Google Scholar] [CrossRef]
- Cheruiyot, E.K.; Haile-Mariam, M.; Cocks, B.G.; MacLeod, I.M.; Xiang, R.; Pryce, J.E. New loci and neuronal pathways for resilience to heat stress in cattle. Sci. Rep. 2021, 11, 16619. [Google Scholar] [CrossRef]
- Edwards, J.L.; Ealy, A.D.; Monterroso, V.H.; Hansen, P.J. Ontogeny of temperature-regulated heat shock protein 70 synthesis in preimplantation bovine embryos. Mol. Reprod. Dev. 1997, 48, 25–33. [Google Scholar] [CrossRef]
- Basiricò, L.; Morera, P.; Primi, V.; Lacetera, N.; Nardone, A.; Bernabucci, U. Cellular thermotolerance is associated with heat shock protein 70.1 genetic polymorphisms in Holstein lactating cows. Cell Stress Chaperones 2011, 16, 441–448. [Google Scholar] [CrossRef]
- Sailo, L.; Gupta, I.; Archana, V.; Ramendra, D.; Chaudhari, M.V. Association of single nucleotide polymorphism of Hsp90ab1 gene with thermotolerance and milk yield in Sahiwal cows. Afr. J. Biochem. Res. 2015, 9, 99–103. [Google Scholar] [CrossRef]
- Diaz, F.A.; Gutierrez-Castillo, E.J.; Foster, B.A.; Hardin, P.T.; Bondioli, K.R.; Jiang, Z. Evaluation of Seasonal Heat Stress on Transcriptomic Profiles and Global DNA Methylation of Bovine Oocytes. Front. Genet. 2021, 12, 699920. [Google Scholar] [CrossRef]
- Kalemkeridou, M.; Nanas, I.; Moutou, K.; Amiridis, G.S.; Tsipourlianos, A.; Dovolou, E.; Mamuris, Z.; Giannoulis, T. Genetic diversity and thermotolerance in Holstein cows: Pathway analysis and marker development using whole-genome sequencing. Reprod. Domest. Anim. 2022, 58, 146–157. [Google Scholar] [CrossRef]
- Luo, H.; Hu, L.; Brito, L.F.; Dou, J.; Sammad, A.; Chang, Y.; Ma, L.; Guo, G.; Liu, L.; Zhai, L.; et al. Weighted single-step GWAS and RNA sequencing reveals key candidate genes associated with physiological indicators of heat stress in Holstein cattle. J. Anim. Sci. Biotechnol. 2022, 13, 108. [Google Scholar] [CrossRef]
- Otto, P.I.; Guimarães, S.E.F.; Verardo, L.L.; Azevedo, A.L.S.; Vandenplas, J.; Sevillano, C.A.; Marques, D.B.D.; Pires, M.D.F.A.; de Freitas, C.; Verneque, R.S.; et al. Genome-wide association studies for heat stress response in Bos taurus × Bos indicus crossbred cattle. J. Dairy Sci. 2019, 102, 8148–8158. [Google Scholar] [CrossRef]
- Zeng, L.; Chen, N.; Ning, Q.; Yao, Y.; Chen, H.; Dang, R.; Zhang, H.; Lei, C. PRLH and SOD1 gene variations associated with heat tolerance in Chinese cattle. Anim. Genet. 2018, 49, 447–451. [Google Scholar] [CrossRef]
- Liu, Y.; Li, D.; Li, H.; Zhou, X.; Wang, G. A novel SNP of the ATP1A1 gene is associated with heat tolerance traits in dairy cows. Mol. Biol. Rep. 2011, 38, 83–88. [Google Scholar] [CrossRef]
- Li, Q.; Han, J.; Du, F.; Ju, Z.; Huang, J.; Wang, J.; Li, R.; Wang, C.; Zhong, J. Novel SNPs in HSP70A1A gene and the association of polymorphisms with thermo tolerance traits and tissue specific expression in Chinese Holstein cattle. Mol. Biol. Rep. 2011, 38, 2657–2663. [Google Scholar] [CrossRef]
- Zeng, L.; Qu, K.; Zhang, J.; Huang, B.; Lei, C. Genes related to heat tolerance in cattle—A review. Anim. Biotechnol. 2022, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Peñagaricano, F.; Driver, A.; Chen, H.; Khatib, H. Differential expression of heat shock protein genes and their splice variants in bovine preimplantation embryos. J. Dairy Sci. 2011, 94, 4174–4182. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Possible Roles for Heat Schock Protein 70 and Glutathione in Protection of the Mammalian Preimplantation Embryo from Heat Shock. Ann. Rev. Biomed. 1999, 1, 5–29. [Google Scholar]
- Velazquez, M.M.L.; Alfaro, N.S.; Dupuy, C.R.F.; Salvetti, N.R.; Rey, F.; Ortega, H.H. Heat shock protein patterns in the bovine ovary and relation with cystic ovarian disease. Anim. Reprod. Sci. 2010, 118, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Payton, R.R.; Rispoli, L.A.; Saxton, A.M.; Edwards, J.L. Impact of heat stress exposure during meiotic maturation on oocyte, surrounding cumulus cell, and embryo RNA populations. J. Reprod. Dev. 2011, 57, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Camargo, L.S.A.; Viana, J.H.M.; Ramos, A.A.; Serapião, R.V.; de Sa, W.F.; Ferreira, A.M.; Guimarães, M.F.M.; do Vale Filho, V.R. Developmental competence and expression of the Hsp 70.1 gene in oocytes obtained from Bos indicus and Bos taurus dairy cows in a tropical environment. Theriogenology 2007, 68, 626–632. [Google Scholar] [CrossRef]
- Souza-Cácares, M.B.; Fialho, A.L.L.; Silva, W.A.L.; Cardoso, C.J.T.; Pöhland, R.; Martins, M.I.M.; Melo-Sterza, F.A. Oocyte quality and heat shock proteins in oocytes from bovine breeds adapted to the tropics under different conditions of environmental thermal stress. Theriogenology 2019, 130, 103–110. [Google Scholar] [CrossRef]
- Roth, Z.; Hansen, P.J. Disruption of nuclear maturation and rearrangement of cytoskeletal elements in bovine oocytes exposed to heat shock during maturation. Reproduction 2005, 129, 235–244. [Google Scholar] [CrossRef]
- Edwards, J.L.; Saxton, A.M.; Lawrence, J.L.; Payton, R.R.; Dunlap, J.R. Exposure to a physiologically relevant elevated temperature hastens in vitro maturation in bovine oocytes. J. Dairy Sci. 2005, 88, 4326–4333. [Google Scholar] [CrossRef]
- Gendelman, M.; Roth, Z. In vivo vs. in vitro models for studying the effects of elevated temperature on the GV-stage oocyte, subsequent developmental competence and gene expression. Anim. Reprod. Sci. 2012, 134, 125–134. [Google Scholar] [CrossRef]
- Stamperna, K.; Giannoulis, T.; Nanas, I.; Kalemkeridou, M.; Dadouli, K.; Moutou, K.; Amiridis, G.S.; Dovolou, E. Short term temperature elevation during IVM affects embryo yield and alters gene expression pattern in oocytes, cumulus cells and blastocysts in cattle. Theriogenology 2020, 156, 36–45. [Google Scholar] [CrossRef]
- De Sousa, P.A.; Caveney, A.; Westhusin, M.E.; Watson, A.J. Temporal patterns of embryonic gene expression and their dependence on oogenetic factors. Theriogenology 1998, 49, 115–128. [Google Scholar] [CrossRef]
- Memili, E.; First, N.L. Zygotic and embryonic gene expression in cow: A review of timing and mechanisms of early gene expression as compared with other species. Zygote 2000, 8, 87–96. [Google Scholar] [CrossRef]
- Eberhardt, B.G.; Satrapa, R.A.; Capinzaiki, C.R.L.; Trinca, L.A.; Barros, C.M. Influence of the breed of bull (Bos taurus indicus vs. Bos taurus taurus) and the breed of cow (Bos taurus indicus, Bos taurus taurus and crossbred) on the resistance of bovine embryos to heat. Anim. Reprod. Sci. 2009, 114, 54–61. [Google Scholar] [CrossRef]
- Sakatani, M.; Alvarez, N.V.; Takahashi, M.; Hansen, P.J. Consequences of physiological heat shock beginning at the zygote stage on embryonic development and expression of stress response genes in cattle. J. Dairy Sci. 2012, 95, 3080–3091. [Google Scholar] [CrossRef]
- Huber, E.; Notaro, U.S.; Recce, S.; Rodríguez, F.M.; Ortega, H.H. Fetal programming in dairy cows: Effect of heat stress on progeny fertility and associations with the hypothalamic-pituitary-adrenal axis functions. Anim. Reprod. Sci. 2020, 216, 106348. [Google Scholar] [CrossRef]
- Pavlok, A.; Kopecný, V.; Lucas-Hahn, A.; Niemann, H. Transcriptional activity and nuclear ultrastructure of 8-cell bovine embryos developed by in vitro maturation and fertilization of oocytes from different growth categories of antral follicles. Mol. Reprod. Dev. 1993, 35, 233–243. [Google Scholar] [CrossRef]
- Frei, R.E.; Schultz, G.A.; Church, R.B. Qualitative and quantitative changes in protein synthesis occur at the 8-16-cell stage of embryogenesis in the cow. J. Reprod. Fertil. 1989, 86, 637–641. [Google Scholar] [CrossRef]
- Lepock, J.R. How do cells respond to their thermal environment? Int. J. Hyperth. Off. J. Eur. Soc. Hyperthermic Oncol. North Am. Hyperth. Gr. 2005, 21, 681–687. [Google Scholar] [CrossRef]
- Sakatani, M.; Bonilla, L.; Dobbs, K.B.; Block, J.; Ozawa, M.; Shanker, S.; Yao, J.Q.; Hansen, P.J. Changes in the transcriptome of morula-stage bovine embryos caused by heat shock: Relationship to developmental acquisition of thermotolerance. Reprod. Biol. Endocrinol. 2013, 11, 3. [Google Scholar] [CrossRef]
- Edwards, J.L.; Hansen, P.J. Elevated temperature increases heat shock protein 70 synthesis in bovine two-cell embryos and compromises function of maturing oocytes. Biol. Reprod. 1996, 55, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Lelièvre, J.-M.; Peynot, N.; Ruffini, S.; Laffont, L.; Le Bourhis, D.; Girard, P.-M.; Duranthon, V. Regulation of heat-inducible HSPA1A gene expression during maternal-to-embryo transition and in response to heat in in vitro-produced bovine embryos. Reprod. Fertil. Dev. 2017, 29, 1868–1881. [Google Scholar] [CrossRef] [PubMed]
- Gwazdauskas, F.C.; Thatcher, W.W.; Wilcox, C.J. Physiological, environmental, and hormonal factors at insemination which may affect conception. J. Dairy Sci. 1973, 56, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Badinga, L.; Collier, R.J.; Thatcher, W.W.; Wilcox, C.J. Effects of climatic and management factors on conception rate of dairy cattle in subtropical environment. J. Dairy Sci. 1985, 68, 78–85. [Google Scholar] [CrossRef] [PubMed]
- de la Sota, R.L.; Burke, J.M.; Risco, C.A.; Moreira, F.; DeLorenzo, M.A.; Thatcher, W.W. Evaluation of timed insemination during summer heat stress in lactating dairy cattle. Theriogenology 1998, 49, 761–770. [Google Scholar] [CrossRef]
- Wise, M.E.; Armstrong, D.V.; Huber, J.T.; Hunter, R.; Wiersma, F. Hormonal alterations in the lactating dairy cow in response to thermal stress. J. Dairy Sci. 1988, 71, 2480–2485. [Google Scholar] [CrossRef]
- De Rensis, F.; Scaramuzzi, R.J. Heat stress and seasonal effects on reproduction in the dairy cow—A review. Theriogenology 2003, 60, 1139–1151. [Google Scholar] [CrossRef]
- Wolfenson, D.; Lew, B.J.; Thatcher, W.W.; Graber, Y.; Meidan, R. Seasonal and acute heat stress effects on steroid production by dominant follicles in cows. Anim. Reprod. Sci. 1997, 47, 9–19. [Google Scholar] [CrossRef]
- Roth, Z. Heat stress, the follicle, and its enclosed oocyte: Mechanisms and potential strategies to improve fertility in dairy cows. Reprod. Domest. Anim. 2008, 43 (Suppl. S2), 238–244. [Google Scholar] [CrossRef]
- Vanselow, J.; Vernunft, A.; Koczan, D.; Spitschak, M.; Kuhla, B. Exposure of Lactating Dairy Cows to Acute Pre-Ovulatory Heat Stress Affects Granulosa Cell-Specific Gene Expression Profiles in Dominant Follicles. PLoS ONE 2016, 11, e0160600. [Google Scholar] [CrossRef]
- Greve, T.; Groendahl, C.; Schmidt, M.; Hunter, R.H.F.; Avery, B. Bovine preovulatory follicular temperature: Implications for in vitro production of embryos. Arch. Fuer Tierz. 1996, 7–14. [Google Scholar]
- López-Gatius, F.; Hunter, R. Clinical relevance of pre-ovulatory follicular temperature in heat-stressed lactating dairy cows. Reprod. Domest. Anim. 2017, 52, 366–370. [Google Scholar] [CrossRef]
- López-Gatius, F.; Hunter, R.H.F. Pre-ovulatory follicular temperature in bi-ovular cows. J. Reprod. Dev. 2019, 65, 191–194. [Google Scholar] [CrossRef]
- Ominski, K.H.; Kennedy, A.D.; Wittenberg, K.M.; Nia, S.A.M. Physiological and production responses to feeding schedule in lactating dairy cows exposed to short-term, moderate heat stress. J. Dairy Sci. 2002, 85, 730–737. [Google Scholar] [CrossRef]
- Clarke, I.J. Interface between metabolic balance and reproduction in ruminants: Focus on the hypothalamus and pituitary. Horm. Behav. 2014, 66, 15–40. [Google Scholar] [CrossRef]
- Roland, A.V.; Moenter, S.M. Regulation of gonadotropin-releasing hormone neurons by glucose. Trends Endocrinol. Metab. 2011, 22, 443–449. [Google Scholar] [CrossRef]
- Ioannis, N.; Dimitrios, P.; Katerina, D.; Thomas-Markos, C.; Maria, S.; Panagiotis, G.; Georgios, S.A. Age, gestational and heat stress effects on ghrelin secretion in dairy cattle. Theriogenology 2021, 176, 82–93. [Google Scholar] [CrossRef]
- Chouzouris, T.M.; Dovolou, E.; Dafopoulos, K.; Georgoulias, P.; Vasileiou, N.G.; Fthenakis, G.C.; Anifandis, G.; Amiridis, G.S. Ghrelin suppresses the GnRH-induced preovulatory gonadotropin surge in dairy heifers. Theriogenology 2016, 86, 1615–1621. [Google Scholar] [CrossRef]
- Seminara, S.B.; Messager, S.; Chatzidaki, E.E.; Thresher, R.R.; Acierno, J.S.J.; Shagoury, J.K.; Bo-Abbas, Y.; Kuohung, W.; Schwinof, K.M.; Hendrick, A.G.; et al. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003, 349, 1614–1627. [Google Scholar] [CrossRef]
- Kadokawa, H.; Suzuki, S.; Hashizume, T. Kisspeptin-10 stimulates the secretion of growth hormone and prolactin directly from cultured bovine anterior pituitary cells. Anim. Reprod. Sci. 2008, 105, 404–408. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Iwasa, T.; Kinouchi, R.; Yoshida, S.; Murakami, M.; Gereltsetseg, G.; Yamamoto, S.; Kuwahara, A.; Yasui, T.; Irahara, M. Fasting reduces the kiss1 mRNA levels in the caudal hypothalamus of gonadally intact adult female rats. Endocr. J. 2011, 58, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Kinsey-Jones, J.S.; Li, X.F.; Knox, A.M.I.; Wilkinson, E.S.; Zhu, X.L.; Chaudhary, A.A.; Milligan, S.R.; Lightman, S.L.; O’Byrne, K.T. Down-regulation of hypothalamic Kisspeptin and its receptor, Kiss1r, mRNA expression is associated with stress-induced suppression of luteinising hormone secretion in the female rat. J. Neuroendocr. 2009, 21, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-D.; Kim, Y.-J.; Jang, M.; Bae, S.-G.; Yun, S.-H.; Lee, M.-R.; Seo, Y.-R.; Cho, J.-K.; Kim, S.-J.; Lee, W.-J. Heat Stress during Summer Attenuates Expression of the Hypothalamic Kisspeptin, an Upstream Regulator of the Hypothalamic-Pituitary-Gonadal Axis, in Domestic Sows. Animals 2022, 12, 2967. [Google Scholar] [CrossRef] [PubMed]
- Gwazdauskas, F.C.; Thatcher, W.; Kiddy, C.; Paape, M.; Wilcox, C. Hormonal patterns during heat stress following PGF(2)alpha-tham salt induced luteal regression in heifers. Theriogenology 1981, 16, 271–285. [Google Scholar] [CrossRef]
- Howell, J.L.; Fuquay, J.W.; Smith, A.E. Corpus luteum growth and function in lactating Holstein cows during spring and summer. J. Dairy Sci. 1994, 77, 735–739. [Google Scholar] [CrossRef]
- Ronchi, B.; Stradaioli, G.; Supplizi, A.V.; Bernabucci, U.; Lacetera, N.; Accorsi, P.A.; Nardone, A.; Seren, E. Influence of heat stress or feed restriction on plasma progesterone, oestradiol-17β, LH, FSH, prolactin and cortisol in Holstein heifers. Livest. Prod. Sci. 2001, 68, 231–241. [Google Scholar] [CrossRef]
- Roman-Ponce, H.; Thatcher, W.W.; Wilcox, C.J. Hormonal interelationships and physiological responses of lactating dairy cows to a shade management system in a subtropical environment. Theriogenology 1981, 16, 139–154. [Google Scholar] [CrossRef]
- Gilad, E.; Meidan, R.; Berman, A.; Graber, Y.; Wolfenson, D. Effect of heat stress on tonic and GnRH-induced gonadotrophin secretion in relation to concentration of oestradiol in plasma of cyclic cows. Reproduction 1993, 99, 315–321. [Google Scholar] [CrossRef]
- Madan, M.L.; Johnson, H.D. Environmental Heat Effects on Bovine Luteinizing Hormone. J. Dairy Sci. 1973, 56, 1420–1423. [Google Scholar] [CrossRef]
- Tauck, S.A.; Olsen, J.R.; Wilkinson, J.R.C.; Wedlake, R.J.; Davis, K.C.; Berardinelli, J.G. Characteristics of temporal patterns of cortisol and luteinizing hormone in primiparous, postpartum, anovular, suckled, beef cows exposed acutely to bulls. Reprod. Biol. Endocrinol. 2010, 8, 89. [Google Scholar] [CrossRef]
- Wolfenson, D.; Thatcher, W.W.; Badinga, L.; Savio, J.D.; Meidan, R.; Lew, B.J.; Braw-Tal, R.; Berman, A. Effect of heat stress on follicular development during the estrous cycle in lactating dairy cattle. Biol. Reprod. 1995, 52, 1106–1113. [Google Scholar] [CrossRef]
- Wilson, S.J.; Marion, R.S.; Spain, J.N.; Spiers, D.E.; Keisler, D.H.; Lucy, M.C. Effects of controlled heat stress on ovarian function of dairy cattle. 1. Lactating cows. J. Dairy Sci. 1998, 81, 2124–2131. [Google Scholar] [CrossRef]
- Roth, Z. Physiology and Endocrinology Symposium: Cellular and molecular mechanisms of heat stress related to bovine ovarian function. J. Anim. Sci. 2015, 93, 2034–2044. [Google Scholar] [CrossRef]
- Wolfenson, D.; Roth, Z.; Meidan, R. Impaired reproduction in heat-stressed cattle: Basic and applied aspects. Anim. Rep. Sci. 2000, 60–61, 535–547. [Google Scholar] [CrossRef]
- Roth, Z.; Meidan, R.; Braw-Tal, R.; Wolfenson, D. Immediate and delayed effects of heat stress on follicular development and its association with plasma FSH and inhibin concentration in cows. J. Reprod. Fertil. 2000, 120, 83–90. [Google Scholar] [CrossRef]
- Kaneko, H.; Nakanishi, Y.; Taya, K.; Kishi, H.; Watanabe, G.; Sasamoto, S.; Hasegawa, Y. Evidence that inhibin is an important factor in the regulation of FSH secretion during the mid-luteal phase in cows. J. Endocrinol. 1993, 136, 35–41. [Google Scholar] [CrossRef]
- Kaneko, H.; Taya, K.; Watanabe, G.; Noguchi, J.; Kikuchi, K.; Shimada, A.; Hasegawa, Y. Inhibin is involved in the suppression of FSH secretion in the growth phase of the dominant follicle during the early luteal phase in cows. Domest. Anim. Endocrinol. 1997, 14, 263–271. [Google Scholar] [CrossRef]
- Richards, J.S.; Midgley, A.R.J. Protein hormone action: A key to understanding ovarian follicular and luteal cell development. Biol. Reprod. 1976, 14, 82–94. [Google Scholar] [CrossRef]
- Li, J.; Gao, H.; Tian, Z.; Wu, Y.; Wang, Y.; Fang, Y.; Lin, L.; Han, Y.; Wu, S.; Haq, I.; et al. Effects of chronic heat stress on granulosa cell apoptosis and follicular atresia in mouse ovary. J. Anim. Sci. Biotechnol. 2016, 7, 57. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Shen, M.; Wu, W.-J.; Li, B.-J.; Weng, Q.-N.; Li, M.; Liu, H.-L. Expression of PUMA in Follicular Granulosa Cells Regulated by FoxO1 Activation during Oxidative Stress. Reprod. Sci. 2015, 22, 696–705. [Google Scholar] [CrossRef]
- Fu, Y.; He, C.-J.; Ji, P.-Y.; Zhuo, Z.-Y.; Tian, X.-Z.; Wang, F.; Tan, D.-X.; Liu, G.-S. Effects of melatonin on the proliferation and apoptosis of sheep granulosa cells under thermal stress. Int. J. Mol. Sci. 2014, 15, 21090–21104. [Google Scholar] [CrossRef] [PubMed]
- Alemu, T.W.; Pandey, H.O.; Wondim, D.S.; Gebremedhn, S.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Oxidative and endoplasmic reticulum stress defense mechanisms of bovine granulosa cells exposed to heat stress. Theriogenology 2018, 110, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Averill-Bates, D.A. Thermotolerance induced at a mild temperature of 40 °C alleviates heat shock-induced ER stress and apoptosis in HeLa cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2015, 1853, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Park, S.-J.; Kim, T.-S.; Park, H.-J.; Park, J.; Kim, B.K.; Kim, G.-R.; Kim, J.-M.; Huang, S.M.; Chae, J.-I.; et al. Testicular hyperthermia induces Unfolded Protein Response signaling activation in spermatocyte. Biochem. Biophys. Res. Commun. 2013, 434, 861–866. [Google Scholar] [CrossRef]
- Shimizu, T.; Ohshima, I.; Ozawa, M.; Takahashi, S.; Tajima, A.; Shiota, M.; Miyazaki, H.; Kanai, Y. Heat stress diminishes gonadotropin receptor expression and enhances susceptibility to apoptosis of rat granulosa cells. Reproduction 2005, 129, 463–472. [Google Scholar] [CrossRef]
- Rispoli, L.A.; Payton, R.; Gondro, C.; Saxton, A.; Nagle, K.; Jenkins, B.W.; Schrick, F.; Edwards, J.L. Heat stress effects on the cumulus cells surrounding the bovine oocyte during maturation: Altered matrix metallopeptidase 9 and progesterone production. Reproduction 2013, 146, 193–207. [Google Scholar] [CrossRef]
- McKenzie, K.A.; Dias, J.A.; Cohen, B.D. Investigation of human follicle stimulating hormone residency in membrane microdomains. FASEB J. 2009, 23, 880.7. [Google Scholar] [CrossRef]
- Trout, J.P.; McDowell, L.R.; Hansen, P.J. Characteristics of the estrous cycle and antioxidant status of lactating Holstein cows exposed to heat stress. J Dairy Sci. 1998, 81, 1244–1250. [Google Scholar] [CrossRef]
- Bridges, P.J.; Brusie, M.A.; Fortune, J.E. Elevated temperature (heat stress) in vitro reduces androstenedione and estradiol and increases progesterone secretion by follicular cells from bovine dominant follicles. Domest. Anim. Endocrinol. 2005, 29, 508–522. [Google Scholar] [CrossRef]
- Badinga, L.; Thatcher, W.; Diaz, T.; Drost, M.; Wolfenson, D. Effect of environmental heat stress on follicular development and steroidogenesis in lactating Holstein cows. Theriogenology 1993, 39, 797–810. [Google Scholar] [CrossRef]
- Nanas, I.; Barbagianni, M.; Dadouli, K.; Dovolou, E.; Amiridis, G.S. Ultrasonographic findings of the corpus luteum and the gravid uterus during heat stress in dairy cattle. Reprod. Domest. Anim. 2021, 56, 1329–1341. [Google Scholar] [CrossRef]
- Scully, S.; Evans, A.C.O.; Carter, F.; Duffy, P.; Lonergan, P.; Crowe, M.A. Ultrasound monitoring of blood flow and echotexture of the corpus luteum and uterus during early pregnancy of beef heifers. Theriogenology 2015, 83, 449–458. [Google Scholar] [CrossRef]
- Siqueira, L.G.; Arashiro, E.K.; Ghetti, A.M.; Souza, E.D.; Feres, L.F.; Pfeifer, L.F.; Fonseca, J.F.; Viana, J.H. Vascular and morphological features of the corpus luteum 12 to 20 days after timed artificial insemination in dairy cattle. J. Dairy Sci. 2019, 102, 5612–5622. [Google Scholar] [CrossRef]
- Wolfenson, D.; Sonego, H.; Bloch, A.; Shaham-Albalancy, A.; Kaim, M.; Folman, Y.; Meidan, R. Seasonal differences in progesterone production by luteinized bovine thecal and granulosa cells. Domest. Anim. Endocrinol. 2002, 22, 81–90. [Google Scholar] [CrossRef]
- Mann, G.E.; Lamming, G.E.; Robinson, R.S.; Wathes, D.C. The regulation of interferon-tau production and uterine hormone receptors during early pregnancy. J. Reprod. Fertil. Suppl. 1999, 54, 317–328. [Google Scholar]
- Ahmad, N.; Schrick, F.N.; Butcher, R.L.; Inskeep, E.K. Effect of persistent follicles on early embryonic losses in beef cows. Biol. Reprod. 1995, 52, 1129–1135. [Google Scholar] [CrossRef]
- Christison, G.I.; Johnson, H.D. Cortisol turnover in heat-stressed cows. J. Anim. Sci. 1972, 35, 1005–1010. [Google Scholar] [CrossRef]
- Yadav, B.; Singh, G.; Wankar, A. Adaptive Capability as Indicated by Redox Status and Endocrine Responses in Crossbred Cattle Exposed to Thermal Stress. J. Anim. Res. 2015, 5, 67–73. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Peng, D.; Li, G.; Chen, J.; Gu, X. Exposure to heat-stress environment affects the physiology, circulation levels of cytokines, and microbiome in dairy cows. Sci. Rep. 2018, 8, 14606. [Google Scholar] [CrossRef]
- Pereira, A.M.F.; Titto, E.L.; Infante, P.; Titto, C.G.; Geraldo, A.M.; Alves, A.; Leme, T.M.; Baccari, F.J.; Almeida, J.A. Evaporative heat loss in Bos taurus: Do different cattle breeds cope with heat stress in the same way? J. Therm. Biol. 2014, 45, 87–95. [Google Scholar] [CrossRef]
- Hansen, P.J. Physiological and cellular adaptations of zebu cattle to thermal stress. Anim. Reprod. Sci. 2004, 82–83, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Stamperna, K.; Giannoulis, T.; Dovolou, E.; Kalemkeridou, M.; Nanas, I.; Dadouli, K.; Moutou, K.; Mamuris, Z.; Amiridis, G.S. Heat Shock Protein 70 Improves In Vitro Embryo Yield and Quality from Heat Stressed Bovine Oocytes. Animals 2021, 11, 1794. [Google Scholar] [CrossRef] [PubMed]
- Tilbrook, A.J.; Canny, B.J.; Serapiglia, M.D.; Ambrose, T.J.; Clarke, I.J. Suppression of the secretion of luteinizing hormone due to isolation/restraint stress in gonadectomised rams and ewes is influenced by sex steroids. J. Endocrinol. 1999, 160, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.S.; Dobson, H.; Ward, W.R. Relationship between an increase in plasma cortisol during transport-induced stress and failure of oestradiol to induce a luteinising hormone surge in dairy cows. Res. Vet. Sci. 1990, 49, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, D.F.; Krasa, H.B.; Padmanabhan, V.; Viguié, C.; Karsch, F.J. Endocrine alterations that underlie endotoxin-induced disruption of the follicular phase in ewes. Biol. Reprod. 2000, 62, 45–53. [Google Scholar] [CrossRef]
- Peter, A.T.; Bosu, W.T.; DeDecker, R.J. Suppression of preovulatory luteinizing hormone surges in heifers after intrauterine infusions of Escherichia coli endotoxin. Am. J. Vet. Res. 1989, 50, 368–373. [Google Scholar]
- Breen, K.M.; Billings, H.J.; Wagenmaker, E.R.; Wessinger, E.W.; Karsch, F.J. Endocrine basis for disruptive effects of cortisol on preovulatory events. Endocrinology 2005, 146, 2107–2115. [Google Scholar] [CrossRef]
- Schreiber, J.R.; Nakamura, K.; Erickson, G.F. Rat ovary glucocorticoid receptor: Identification and characterization. Steroids 1982, 39, 569–584. [Google Scholar] [CrossRef]
- Hsueh, A.J.; Erickson, G.F. Glucocorticoid inhibition of FSH-induced estrogen production in cultured rat granulosa cells. Steroids 1978, 32, 639–648. [Google Scholar] [CrossRef]
- Riddle, O.; Bates, R.W.; Dykshorn, S.W. The Preparation, Identification and Assay of Prolactin—A hormone of the anterior pituitary. Am. J. Physiol. Content 1933, 105, 191–216. [Google Scholar] [CrossRef]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef]
- Scharf, B.; Leonard, M.J.; Weaber, R.L.; Mader, T.L.; Hahn, G.L.; Spiers, D.E. Determinants of bovine thermal response to heat and solar radiation exposures in a field environment. Int. J. Biometeorol. 2011, 55, 469–480. [Google Scholar] [CrossRef]
- Tucker, H.A.; Wettemann, R.P. Effects of ambient temperature and relative humidity on serum prolactin and growth hormone in heifers. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1976, 151, 623–626. [Google Scholar] [CrossRef]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat stress abatement during the dry period influences metabolic gene expression and improves immune status in the transition period of dairy cows. J. Dairy Sci. 2011, 94, 86–96. [Google Scholar] [CrossRef]
- Igono, M.O.; Johnson, H.D.; Steevens, B.J.; Hainen, W.A.; Shanklin, M.D. Effect of season on milk temperature, milk growth hormone, prolactin, and somatic cell counts of lactating cattle. Int. J. Biometeorol. 1988, 32, 194–200. [Google Scholar] [CrossRef]
- Collier, R.J.; Doelger, S.G.; Head, H.H.; Thatcher, W.W.; Wilcox, C.J. Effects of heat stress during pregnancy on maternal hormone concentrations, calf birth weight and postpartum milk yield of Holstein cows. J. Anim. Sci. 1982, 54, 309–319. [Google Scholar] [CrossRef]
- Do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, J.; Bubolz, J.; Dahl, G.E. Heat stress abatement during the dry period influences prolactin signaling in lymphocytes. Domest. Anim. Endocrinol. 2010, 38, 38–45. [Google Scholar] [CrossRef]
- Ouellet, V.; Negrao, J.; Skibiel, A.L.; Lantigua, V.A.; Fabris, T.F.; Marrero, M.G.; Dado-Senn, B.; Laporta, J.; Dahl, G.E. Endocrine Signals Altered by Heat Stress Impact Dairy Cow Mammary Cellular Processes at Different Stages of the Dry Period. Animals 2021, 11, 563. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P., Jr. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef]
- Tortonese, D.J.; Brooks, J.; Ingleton, P.M.; McNeilly, A.S. Detection of prolactin receptor gene expression in the sheep pituitary gland and visualization of the specific translation of the signal in gonadotrophs. Endocrinology 1998, 139, 5215–5223. [Google Scholar] [CrossRef]
- Gregory, S.J.; Townsend, J.; McNeilly, A.S.; Tortonese, D.J. Effects of prolactin on the luteinizing hormone response to gonadotropin-releasing hormone in primary pituitary cell cultures during the ovine annual reproductive cycle. Biol. Reprod. 2004, 70, 1299–1305. [Google Scholar] [CrossRef] [PubMed]
- Mcneilly, A.S.; Friesen, H.G. Prolactin during Pregnancy and Lactation in the Rabbit. Endocrinology 1978, 102, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, A.S.; Lincoln, D.W. Influence of pinealectomy on the suppression of gonadotrophin secretion induced by hyperprolactinaemia in the adult male rat. J. Endocrinol. 1983, 97, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Boyar, R.M.; Kapen, S.; Finkelstein, J.W.; Perlow, M.; Sassin, J.F.; Fukushima, D.K.; Weitzman, E.D.; Hellman, L. Hypothalamic-pituitary function in diverse hyperprolactinemic states. J. Clin. Investig. 1974, 53, 1588–1598. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, A.S.; Sharpe, R.M.; Davidson, D.W.; Fraser, H.M. Inhibition of gonadotrophin secretion by induced hyperprolactinaemia in the male rat. J. Endocrinol. 1978, 79, 59–68. [Google Scholar] [CrossRef]
- Smith, M.S. Effect of pulsatile gonadotropin-releasing hormone on the release of luteinizing hormone and follicle-stimulating hormone in vitro by anterior pituitaries from lactating and cycling rats. Endocrinology 1982, 110, 882–891. [Google Scholar] [CrossRef]
- Cheung, C.Y. Prolactin suppresses luteinizing hormone secretion and pituitary responsiveness to luteinizing hormone-releasing hormone by a direct action at the anterior pituitary. Endocrinology 1983, 113, 632–638. [Google Scholar] [CrossRef]
- Davis, S.L.; Hill, K.M.; Ohlson, D.L.; Jacobs, J.A. Influence of Chronic Thyrotropin-Releasing Hormone Injections on Secretion of Prolactin, Thyrotropin and Growth Hormone and on Growth Rate in Wether Lambs. J. Anim. Sci. 1976, 42, 1244–1250. [Google Scholar] [CrossRef]
- Curlewis, J.D. Seasonal prolactin secretion and its role in seasonal reproduction: A review. Reprod. Fertil. Dev. 1992, 4, 1–23. [Google Scholar] [CrossRef]
- Ginther, O.J.; Kot, K.; Kulick, L.J.; Wiltbank, M.C. Emergence and deviation of follicles during the development of follicular waves in cattle. Theriogenology 1997, 48, 75–87. [Google Scholar]
- Fortune, J.E. Ovarian follicular growth and development in mammals. Biol. Reprod. 1994, 50, 225–232. [Google Scholar] [CrossRef]
- De Rensis, F.; Peters, A.R. The control of follicular dynamics by PGF2α, gnrh, hCG and oestrus synchronization in cattle. Reprod. Domest. Anim. 1999, 34, 49–59. [Google Scholar] [CrossRef]
- Lussier, J.G.; Matton, P.; Dufour, J.J. Growth rates of follicles in the ovary of the cow. Reproduction 1987, 81, 301–307. [Google Scholar] [CrossRef]
- Roth, Z.; Meidan, R.; Wolfenson, D. Delayed effect of heat stress on steroid production in medium-sized and preovulatory bovine follicles. Reprod. -Camb. 2001, 121, 745–751. [Google Scholar] [CrossRef]
- Roth, Z.; Arav, A.; Bor, A.; Zeron, Y.; Braw-Tal, R.; Wolfenson, D. Improvement of quality of oocytes collected in the autumn by enhanced removal of impaired follicles from previously heat-stressed cows. Reproduction 2001, 122, 737–744. [Google Scholar] [CrossRef]
- Xu, M.; Fazleabas, A.T.; Shikanov, A.; Jackson, E.; Barrett, S.L.; Hirshfeld-Cytron, J.; Kiesewetter, S.E.; Shea, L.D.; Woodruff, T.K. In vitro oocyte maturation and preantral follicle culture from the luteal-phase baboon ovary produce mature oocytes. Biol. Reprod. 2011, 84, 689–697. [Google Scholar] [CrossRef]
- Monniaux, D.; Genêt, C.; Maillard, V.; Jarrier, P.; Adriaensen, H.; Hennequet-Antier, C.; Lainé, A.-L.; Laclie, C.; Papillier, P.; Plisson-Petit, F.; et al. Prenatal programming by testosterone of follicular theca cell functions in ovary. Cell. Mol. Life Sci. 2020, 77, 1177–1196. [Google Scholar] [CrossRef]
- de Aguiar, L.H.; Hyde, K.A.; Pedroza, G.H.; Denicol, A.C. Heat stress impairs in vitro development of preantral follicles of cattle. Anim. Reprod. Sci. 2020, 213, 106277. [Google Scholar] [CrossRef]
- Paes, V.M.; Vieira, L.A.; Correia, H.H.V.; Sa, N.A.R.; Moura, A.A.A.; Sales, A.D.; Rodrigues, A.P.R.; Magalhães-Padilha, D.M.; Santos, F.W.; Apgar, G.A.; et al. Effect of heat stress on the survival and development of in vitro cultured bovine preantral follicles and on in vitro maturation of cumulus-oocyte complex. Theriogenology 2016, 86, 994–1003. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef]
- Hatami, S.; Zavareh, S.; Salehnia, M.; Lashkarbolouki, T.; Karimi, I. Comparison of oxidative status of mouse pre-antral follicles derived from vitrified whole ovarian tissue and vitrified pre-antral follicles in the presence of alpha lipoic acid. J. Obstet. Gynaecol. Res. 2014, 40, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- Cardone, D.A.; Cáceres, A.R.R.; Sanhueza, M.A.; Bruna, F.A.; Laconi, M.R. Effects of short-term in vitro heat stress on bovine preantral follicles. Livest. Sci. 2022, 264, 105076. [Google Scholar] [CrossRef]
- Guzeloglu, A.; Ambrose, J.D.; Kassa, T.; Diaz, T.; Thatcher, M.J.; Thatcher, W.W. Long-term follicular dynamics and biochemical characteristics of dominant follicles in dairy cows subjected to acute heat stress. Anim. Reprod. Sci. 2001, 66, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Boland, M.P. Frequency of twin births among Holstein-Friesian cows in a warm dry climate. Theriogenology 1991, 36, 1–10. [Google Scholar] [CrossRef]
- De Rensis, F.; Saleri, R.; Garcia-Ispierto, I.; Scaramuzzi, R.; López-Gatius, F. Effects of Heat Stress on Follicular Physiology in Dairy Cows. Animals 2021, 11, 3406. [Google Scholar] [CrossRef]
- Boni, R. Heat stress, a serious threat to reproductive function in animals and humans. Mol. Reprod. Dev. 2019, 86, 1307–1323. [Google Scholar] [CrossRef]
- Sammad, A.; Umer, S.; Shi, R.; Zhu, H.; Zhao, X.; Wang, Y. Dairy cow reproduction under the influence of heat stress. J. Anim. Physiol. Anim. Nutr. 2020, 104, 978–986. [Google Scholar] [CrossRef]
- Sirard, M.A.; Florman, H.M.; Leibfried-Rutledge, M.L.; Barnes, F.L.; Sims, M.L.; First, N.L. Timing of Nuclear Progression and Protein Synthesis Necessary for Meiotic Maturation of Bovne Oocytes. Biol. Reprod. 1989, 40, 1257–1263. [Google Scholar] [CrossRef]
- Lonergan, P.; Fair, T.; Khatir, H.; Cesaroni, G.; Mermillod, P. Effect of protein synthesis inhibition before or during in vitro maturation on subsequent development of bovine oocytes. Theriogenology 1998, 50, 417–431. [Google Scholar] [CrossRef]
- Vigneron, C.; Perreau, C.; Dupont, J.; Uzbekova, S.; Prigent, C.; Mermillod, P. Several signaling pathways are involved in the control of cattle oocyte maturation. Mol. Reprod. Dev. Inc. Gamete Res. 2004, 69, 466–474. [Google Scholar] [CrossRef]
- Salhab, M.; Dhorne-Pollet, S.; Auclair, S.; Guyader-Joly, C.; Brisard, D.; Dalbies-Tran, R.; Dupont, J.; Ponsart, C.; Mermillod, P.; Uzbekova, S. In vitro maturation of oocytes alters gene expression and signaling pathways in bovine cumulus cells. Mol. Reprod. Dev. 2013, 80, 166–182. [Google Scholar] [CrossRef]
- Payton, R.R.; Romar, R.; Coy, P.; Saxton, A.M.; Lawrence, J.L.; Edwards, J.L. Susceptibility of Bovine Germinal Vesicle-Stage Oocytes from Antral Follicles to Direct Effects of Heat Stress in Vitro. Biol. Reprod. 2004, 71, 1303–1308. [Google Scholar] [CrossRef]
- Soto, P.; Smith, L.C. BH4 peptide derived from Bcl-xL and Bax-inhibitor peptide suppresses apoptotic mitochondrial changes in heat stressed bovine oocytes. Mol. Reprod. Dev. 2009, 76, 637–646. [Google Scholar] [CrossRef]
- Sun, Q.-Y.; Schatten, H. Regulation of dynamic events by microfilaments during oocyte maturation and fertilization. Reproduction 2006, 131, 193–205. [Google Scholar] [CrossRef]
- Payton, R.R.; Rispoli, L.A.; Nagle, K.A.; Gondro, C.; Saxton, A.M.; Voy, B.H.; Edwards, J.L. Mitochondrial-related consequences of heat stress exposure during bovine oocyte maturation persist in early embryo development. J. Reprod. Dev. 2018, 64, 243–251. [Google Scholar] [CrossRef]
- Jordan, E.R. Effects of Heat Stress on Reproduction. J. Dairy Sci. 2003, 86, E104–E114. [Google Scholar] [CrossRef]
- Hansen, P.J. To be or not to be—Determinants of embryonic survival following heat shock. Theriogenology 2007, 68, S40–S48. [Google Scholar] [CrossRef]
- Stamperna, K.; Giannoulis, T.; Dovolou, E.; Kalemkeridou, M.; Nanas, I.; Dadouli, K.; Moutou, K.; Mamuris, Z.; Amiridis, G.S. The effects of heat shock protein 70 addition in the culture medium on the development and quality of in vitro produced heat shocked bovine embryos. Animals 2021, 11, 3347. [Google Scholar] [CrossRef]
- Silva, C.F.; Sartorelli, E.S.; Castilho, A.C.S.; Satrapa, R.A.; Puelker, R.Z.; Razza, E.M.; Ticianelli, J.S.; Eduardo, H.P.; Loureiro, B.; Barros, C.M. Effects of heat stress on development, quality and survival of Bos indicus and Bos taurus embryos produced in vitro. Theriogenology 2013, 79, 351–357. [Google Scholar] [CrossRef]
- Molavian, H.; Tonekaboni, A.M.; Kohandel, M.; Sivaloganathan, S. The synergetic coupling among the cellular antioxidants glutathione peroxidase/peroxiredoxin and other antioxidants and its effect on the concentration of H2O2. Sci. Rep. 2015, 5, srep13620. [Google Scholar] [CrossRef]
- Lonergan, P.; Rizos, D.; Gutierrez-Adan, A.; Moreira, P.M.; Pintado, B.; De La Fuente, J.; Boland, M.P. Temporal divergence in the pattern of messenger RNA expression in bovine embryos cultured from the zygote to blastocyst stage in vitro or in vivo. Biol. Reprod. 2003, 69, 1424–1431. [Google Scholar] [CrossRef] [PubMed]
- Alm, H.; Torner, H.; Löhrke, B.; Viergutz, T.; Ghoneim, I.M.; Kanitz, W. Bovine blastocyst development rate in vitro is influenced by selection of oocytes by brillant cresyl blue staining before IVM as indicator for glucose-6-phosphate dehydrogenase activity. Theriogenology 2005, 63, 2194–2205. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, L.; Chen, X.; Lu, Y.; Wang, D. Effects of heat stress on body temperature, milk production, and reproduction in dairy cows: A novel idea for monitoring and evaluation of heat stress—A review. Asian-Australas. J. Anim. Sci. 2019, 32, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.L.; Hansen, P.J. Differential responses of bovine oocytes and preimplantation embryos to heat shock. Mol. Reprod. Dev. 1997, 46, 138–145. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, K.P.; Singh, M.K.; Saini, N.; Palta, P.; Manik, R.S.; Singla, S.K.; Upadhyay, R.C.; Chauhan, M.S. Effect of Physiologically Relevant Heat Shock on Development, Apoptosis and Expression of Some Genes in Buffalo (Bubalus bubalis) Embryos Produced In Vitro. Reprod. Domest. Anim. 2013, 48, 858–865. [Google Scholar] [CrossRef]
- Cartmill, J.A.; El-Zarkouny, S.Z.; Hensley, B.A.; Rozell, T.G.; Smith, J.F.; Stevenson, J.S. An alternative AI breeding protocol for dairy cows exposed to elevated ambient temperatures before or after calving or both. J. Dairy Sci. 2001, 84, 799–806. [Google Scholar] [CrossRef]
- Leese, H.J. The formation and function of oviduct fluid. Reproduction 1988, 82, 843–856. [Google Scholar] [CrossRef]
- Lamy, J.; Liere, P.; Pianos, A.; Aprahamian, F.; Mermillod, P.; Saint-Dizier, M. Steroid hormones in bovine oviductal fluid during the estrous cycle. Theriogenology 2016, 86, 1409–1420. [Google Scholar] [CrossRef]
- Lamy, J.; Gatien, J.; Dubuisson, F.; Nadal-Desbarats, L.; Salvetti, P.; Mermillod, P.; Saint-Dizier, M. Metabolomic profiling of bovine oviductal fluid across the oestrous cycle using proton nuclear magnetic resonance spectroscopy. Reprod. Fertil. Dev. 2018, 30, 1021–1028. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Wakamiya, K.; Kohka, M.; Yamamoto, Y.; Okuda, K. Summer heat stress affects prostaglandin synthesis in the bovine oviduct. Reproduction 2013, 146, 103–110. [Google Scholar] [CrossRef]
- Kölle, S.; Hughes, B.; Steele, H. Early embryo-maternal communication in the oviduct: A review. Mol. Reprod. Dev. 2020, 87, 650–662. [Google Scholar] [CrossRef]
- Bauersachs, S.; Rehfeld, S.; Ulbrich, S.E.; Mallok, S.; Prelle, K.; Wenigerkind, H.; Einspanier, R.; Blum, H.; Wolf, E. Monitoring gene expression changes in bovine oviduct epithelial cells during the oestrous cycle. J. Mol. Endocrinol. 2004, 32, 449–466. [Google Scholar] [CrossRef]
- Rąpała, Ł.; Starzyński, R.R.; Trzeciak, P.Z.; Dąbrowski, S.; Gajewska, M.; Jurka, P.; Smolarczyk, R.; Duszewska, A.M. Influence of elevated temperature on bovine oviduct epithelial cells (BOECs). PLoS ONE 2018, 13, e0198843. [Google Scholar] [CrossRef]
- Ni, Y.; Wu, F.; Chen, Q.; Cai, J.; Hu, J.; Shen, J.; Zhang, J. Long noncoding RNA and mRNA profiling of hypothalamic-pituitary-mammary gland axis in lactating sows under heat stress. Genomics 2020, 112, 3668–3676. [Google Scholar] [CrossRef]
- Li, Q.; Qiao, J.; Zhang, Z.; Shang, X.; Chu, Z.; Fu, Y.; Chu, M. Identification and analysis of differentially expressed long non-coding RNAs of Chinese Holstein cattle responses to heat stress. Anim. Biotechnol. 2020, 31, 9–16. [Google Scholar] [CrossRef]
- Place, R.F.; Noonan, E.J. Non-coding RNAs turn up the heat: An emerging layer of novel regulators in the mammalian heat shock response. Cell Stress Chaperones 2014, 19, 159–172. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Zhang, R.; Li, D.; Gao, M.-Q. LRRC75A antisense lncRNA1 knockout attenuates inflammatory responses of bovine mammary epithelial cells. Int. J. Biol. Sci. 2020, 16, 251. [Google Scholar] [CrossRef]
- Lavery, K.; Gabler, C.; Day, J.; Killian, G. Expression of haptoglobin mRNA in the liver and oviduct during the oestrous cycle of cows (Bos taurus). Anim. Reprod. Sci. 2004, 84, 13–26. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Almiñana, C.; Tsikis, G.; Labas, V.; Uzbekov, R.; da Silveira, J.C.; Bauersachs, S.; Mermillod, P. Deciphering the oviductal extracellular vesicles content across the estrous cycle: Implications for the gametes-oviduct interactions and the environment of the potential embryo. BMC Genom. 2018, 19, 622. [Google Scholar] [CrossRef] [PubMed]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Ramírez, M.Á.; Yáñez-Mó, M.; Rizos, D. Effect of bovine oviductal extracellular vesicles on embryo development and quality in vitro. Reproduction 2017, 153, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yue, T.; Ahmad, M.J.; Hu, X.; Zhang, X.; Deng, T.; Hu, Y.; He, C.; Zhou, Y.; Yang, L. Transcriptome Analysis Reveals Potential Regulatory Genes Related to Heat Tolerance in Holstein Dairy Cattle. Genes 2020, 11, 68. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dovolou, E.; Giannoulis, T.; Nanas, I.; Amiridis, G.S. Heat Stress: A Serious Disruptor of the Reproductive Physiology of Dairy Cows. Animals 2023, 13, 1846. https://doi.org/10.3390/ani13111846
Dovolou E, Giannoulis T, Nanas I, Amiridis GS. Heat Stress: A Serious Disruptor of the Reproductive Physiology of Dairy Cows. Animals. 2023; 13(11):1846. https://doi.org/10.3390/ani13111846
Chicago/Turabian StyleDovolou, Eleni, Themistoklis Giannoulis, Ioannis Nanas, and Georgios S. Amiridis. 2023. "Heat Stress: A Serious Disruptor of the Reproductive Physiology of Dairy Cows" Animals 13, no. 11: 1846. https://doi.org/10.3390/ani13111846