
Developmental Characteristics of Skeletal Muscle during the Embryonic Stage in Chinese Yellow Quail (Coturnix japonica)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Morphometric Analysis
2.3. RNA Extraction, Reverse Transcription, and Quantitative Real-Time PCR
2.4. Statistical Analysis
3. Results
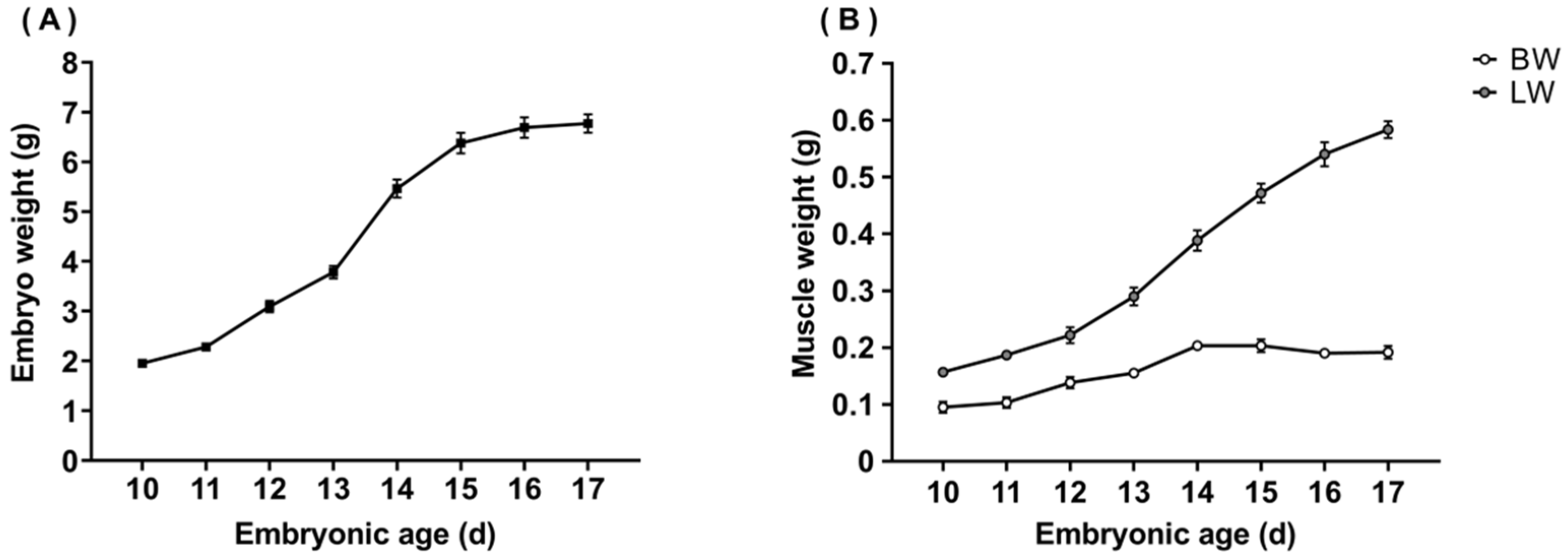
3.1. Weight of Embryos, Breast, and Leg Muscles
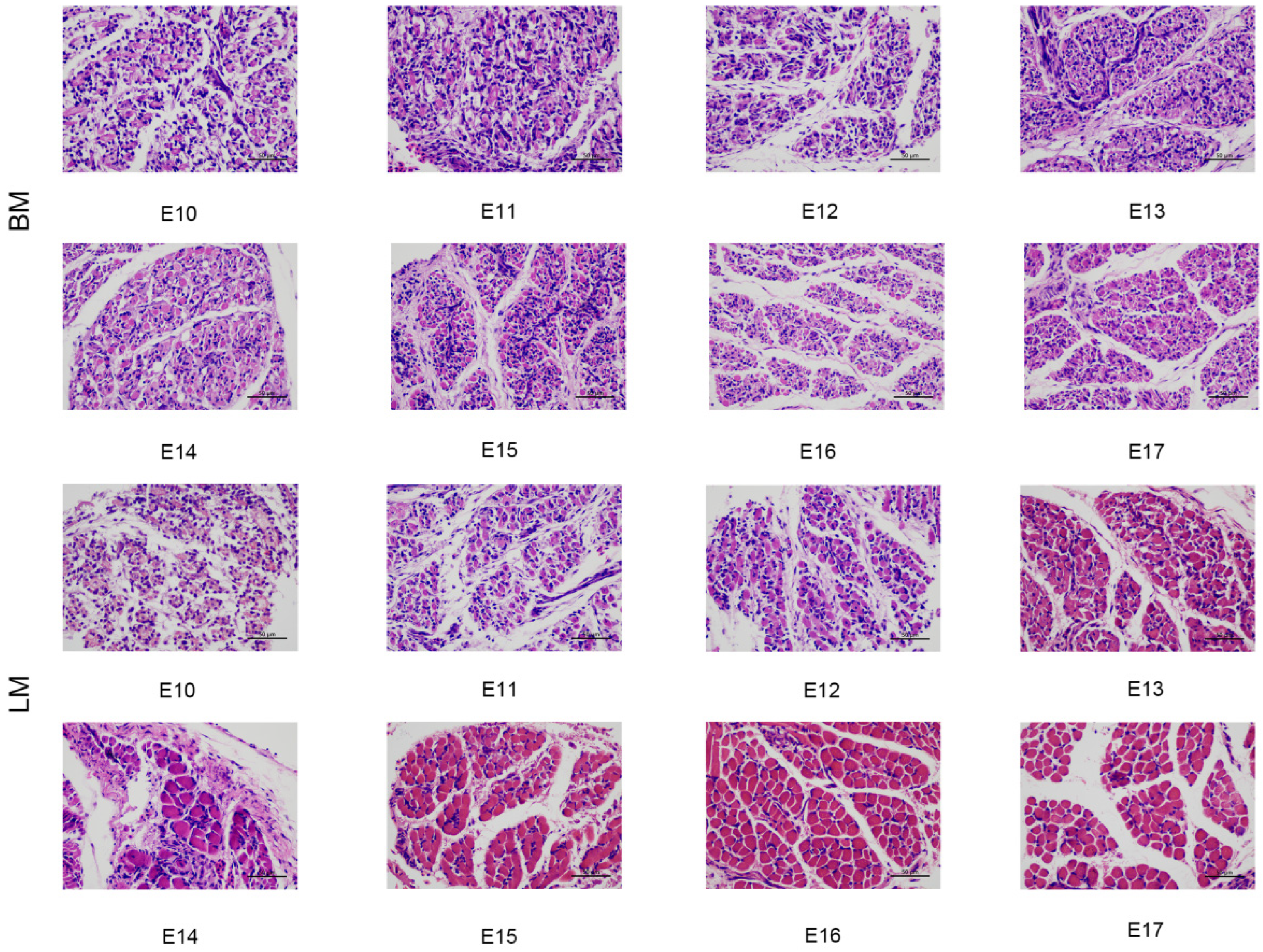
3.2. Histological Morphology of Breast and Leg Muscles
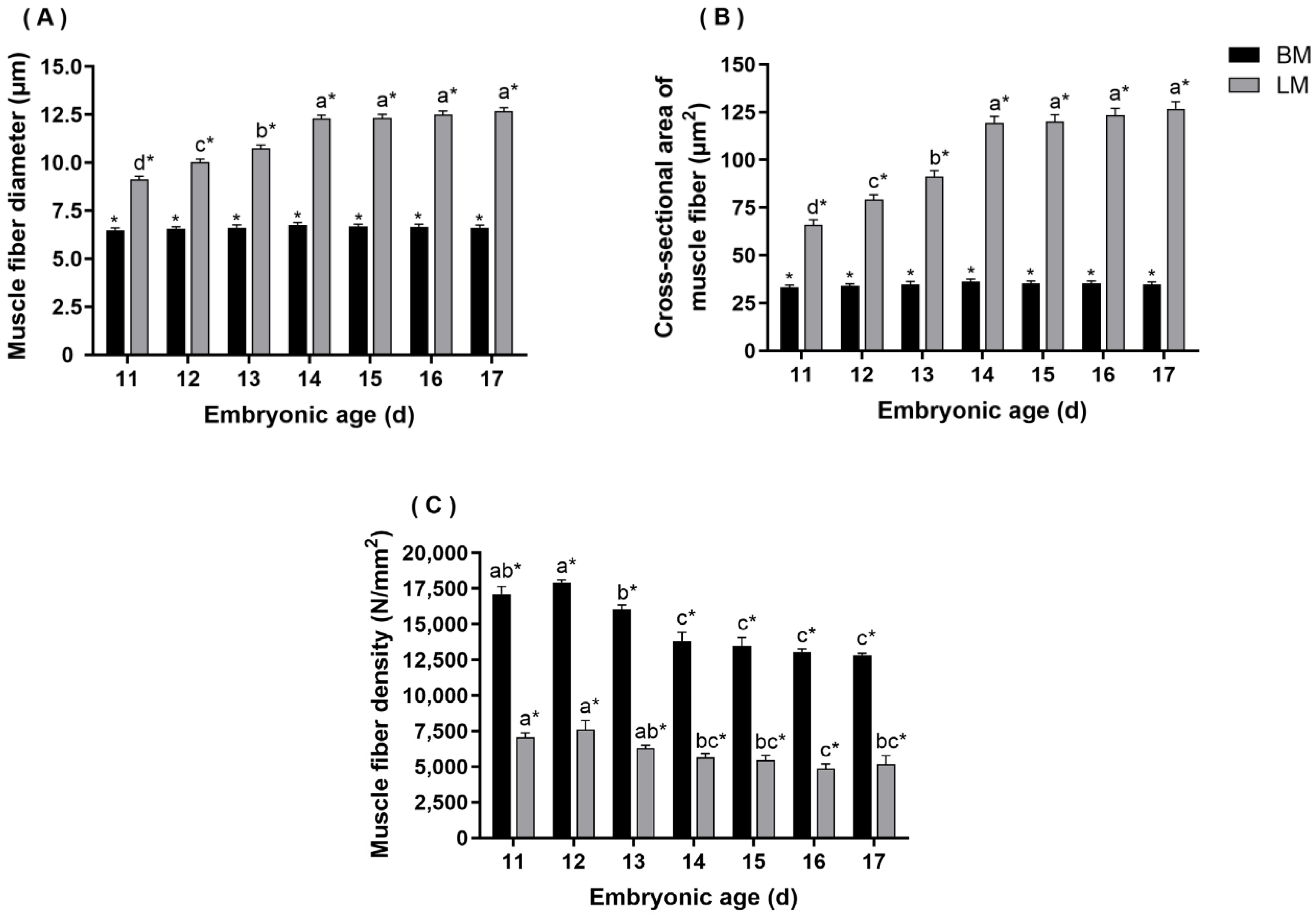
3.3. Morphometric Analysis
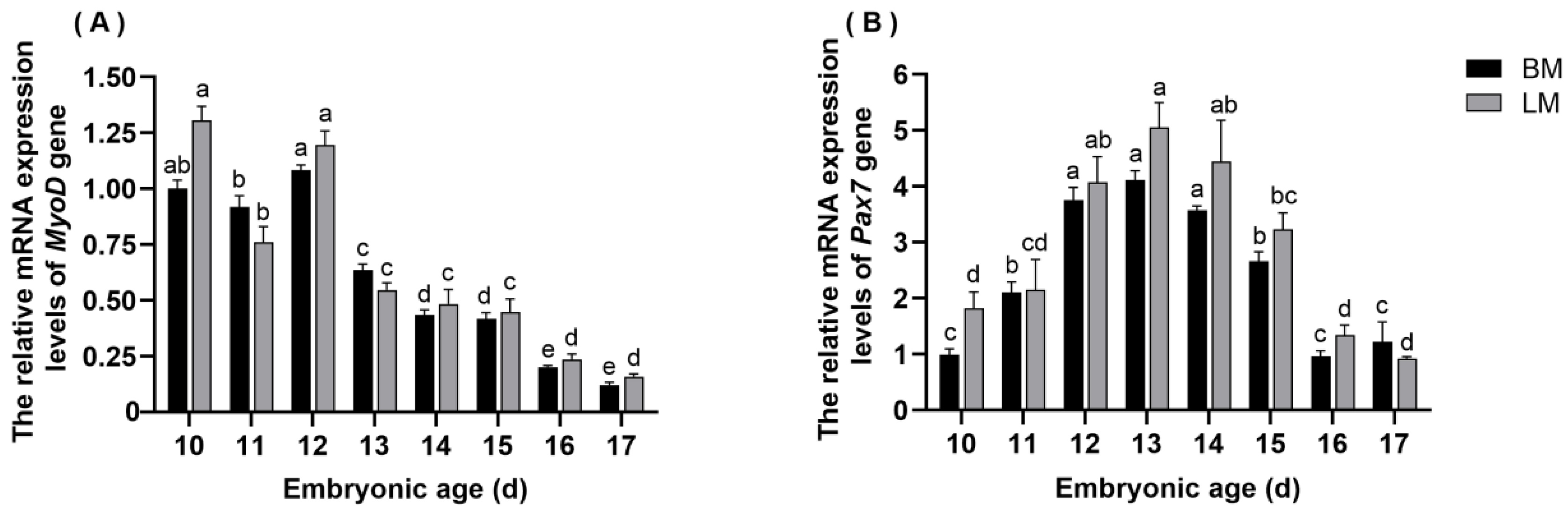
3.4. The Expression Pattern of MyoD in Breast and Leg Muscle Tissues
3.5. The Expression Pattern of Pax7 in Breast and Leg Muscle Tissues
3.6. Correlation Analysis in Different Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building Muscle: Molecular Regulation of Myogenesis. Cold Spring Harbor Perspect. Biol. 2012, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.D.; Ma, Q.G.; Tang, M.Y.; Ji, C. Development of Breast Muscle and Meat Quality in Arbor Acres Broilers, Jingxing 100 Crossbred Chickens and Beijing Fatty Chickens. Meat Sci. 2007, 77, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Shin, S. Application of Quail Model for Studying the Poultry Functional Genomics. Korean J. Poult. Sci. 2017, 44, 103–111. [Google Scholar] [CrossRef]
- Lukanov, H. Domestic Quail (Coturnix japonica domestica), Is There Such Farm Animal? Worlds Poult. Sci. J. 2019, 75, 547–557. [Google Scholar] [CrossRef]
- Sabow, A.B. Carcass Characteristics, Physicochemical Attributes, and Fatty Acid and Amino Acid Compositions of Meat Obtained from Different Japanese Quail Strains. Trop. Anim. Health Prod. 2020, 52, 131–140. [Google Scholar] [CrossRef]
- Mnisi, C.M.; Marareni, M.; Manyeula, F.; Madibana, M.J. A Way Forward for the South African Quail Sector As a Potential Contributor to Food and Nutrition Security Following the Aftermath of COVID-19: A Review. Agric. Food Secur. 2021, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Purohit, A.S.; Reed, C.; Mohan, A. Development and Evaluation of Quail Breakfast Sausage. LWT-Food Sci. Technol. 2016, 69, 447–453. [Google Scholar] [CrossRef]
- Santhi, D.; Kalaikannan, A. Japanese Quail (Coturnix coturnix japonica) Meat: Characteristics and Value Addition. Worlds Poult. Sci. J. 2017, 73, 337–343. [Google Scholar] [CrossRef]
- Braun, T.; Buschhausen-Denker, G.; Bober, E.; Tannich, E.; Arnold, H.H. A Novel Human Muscle Factor Related to but Distinct from MyoD1 Induces Myogenic Conversion in 10T1/2 Fibroblasts. EMBO J. 1989, 8, 701–709. [Google Scholar] [CrossRef]
- Sabourin, L.A.; Rudnicki, M.A. The Molecular Regulation of Myogenesis. Clin. Genet. 2000, 57, 16–25. [Google Scholar] [CrossRef]
- Wright, W.E.; Sassoon, D.A.; Lin, V.K. Myogenin, a Factor Regulating Myogenesis, has a Domain Homologous to MyoD. Cell 1989, 56, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, S.J.; Konieczny, S.F. Identification of MRF4: A New Member of the Muscle Regulatory Factor Gene Family. Genes Dev. 1989, 3, 2050–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Hernandez, M.; Garcia-Gonzalez, E.G.; Brun, C.E.; Rudnicki, M.A. The Myogenic Regulatory Factors, Determinants of Muscle Development, Cell Identity and Regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef]
- Ishibashi, J.; Perry, R.L.; Asakura, A.; Rudnicki, M.A. MyoD Induces Myogenic Differentiation Through Cooperation of Its NH2- and COOH-terminal Regions. J. Cell Biol. 2005, 171, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Fong, A.P.; Tapscott, S.J. Skeletal Muscle Programming and Re-programming. Curr. Opin. Genet. Dev. 2013, 23, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudnicki, M.A.; Schnegelsberg, P.N.; Stead, R.H.; Braun, T.; Arnold, H.H.; Jaenisch, R. MyoD or Myf-5 is Required for the Formation of Skeletal Muscle. Cell 1993, 75, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Kuang, S.; Charge, S.B.; Seale, P.; Huh, M.; Rudnicki, M.A. Distinct Roles for Pax7 and Pax3 in Adult Regenerative Myogenesis. J. Cell Biol. 2006, 172, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, M.; Relaix, F. PAX3 and PAX7 As Upstream Regulators of Myogenesis. Semin. Cell Dev. Biol. 2015, 44, 115–125. [Google Scholar] [CrossRef]
- Lin, J.; Wang, C.; Yang, C.; Fu, S.; Redies, C. Pax3 and Pax7 Interact Reciprocally and Regulate the Expression of Cadherin-7 Through Inducing Neuron Differentiation in the Developing Chicken Spinal Cord. J. Comp. Neurol. 2016, 524, 940–962. [Google Scholar] [CrossRef]
- Mansouri, A.; Stoykova, A.; Torres, M.; Gruss, P. Dysgenesis of Cephalic Neural Crest Derivatives in Pax7-/- Mutant Mice. Development 1996, 122, 831–838. [Google Scholar] [CrossRef]
- Relaix, F.; Rocancourt, D.; Mansouri, A.; Buckingham, M. A Pax3/Pax7-Dependent Population of Skeletal Muscle Progenitor Cells. Nature 2005, 435, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.M.; Suh, Y.; Shin, S.; Lee, K. Skeletal Muscle Characterization of Japanese Quail Line Selectively Bred for Lower Body Weight as an Avian Model of Delayed Muscle Growth with Hypoplasia. PLoS ONE 2014, 9, e95932. [Google Scholar] [CrossRef]
- Kim, D.-H.; Choi, Y.M.; Suh, Y.; Shin, S.; Lee, J.; Hwang, S.; Lee, S.S.; Lee, K. Research Note: Increased Myostatin Expression and Decreased Expression of Myogenic Regulatory Factors in Embryonic Ages in a Quail Line with Muscle Hypoplasia. Poult. Sci. 2021, 100, 100978. [Google Scholar] [CrossRef]
- Wlazlo, L.; Drabik, K.; Al-Shammari, K.I.A.; Batkowska, J.; Nowakowicz-Debek, B.; Gryzinska, M. Use of reactive oxygen species (ozone, hydrogen peroxide) for disinfection of hatching eggs. Poult. Sci 2020, 99, 2478–2484. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Harrison, D.L.; Dayton, A.D. Measuring Skeletal Muscle Fiber Width or Diameter: Variance within and among Methods. J. Food Sci. 1981, 46, 294–295. [Google Scholar] [CrossRef]
- Liu, H.H.; Mao, H.G.; Dong, X.Y.; Cao, H.Y.; Liu, K.; Yin, Z.Z. Expression of MSTN gene and its correlation with pectoralis muscle fiber traits in the domestic pigeons (Columba livia). Poult. Sci 2019, 98, 5265–5271. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E.; Shea, R.E.; Choi, I.-H. Inverse relationship between functional maturity and exponential growth rate of avian skeletal muscle: A constraint on evolutionary response. Evol. Int. J. Org. Evol. 1994, 48, 1080–1088. [Google Scholar] [CrossRef]
- Blom, J.; Lilja, C. A comparative study of embryonic development of some bird species with different patterns of postnatal growth. Zoology 2005, 108, 81–95. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Li, X.; Shen, X.; Xu, D.; Tian, Y.; Huang, Y. Transcriptomic investigation of embryonic pectoral muscle reveals increased myogenic processes in Shitou geese compared to Wuzong geese. Brit Poult. Sci 2021, 62, 650–657. [Google Scholar] [CrossRef]
- Ren, L.T.; Liu, A.F.; Wang, Q.G.; Wang, H.G.; Dong, D.Q.; Liu, L.B. Transcriptome analysis of embryonic muscle development in Chengkou Mountain Chicken. BMC Genom. 2021, 22, 12. [Google Scholar] [CrossRef]
- Li, C.; Xiong, T.; Zhou, M.F.; Wan, L.; Xi, S.W.; Liu, Q.H.; Chen, Y.; Mao, H.R.; Liu, S.F.; Chen, B. Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck. Animasl 2020, 10, 1417. [Google Scholar] [CrossRef]
- Zhu, M.T.; Wang, M.Y.; Shao, Y.Y.; Nan, Y.; Blair, H.T.; Morris, S.T.; Zhao, Z.S.; Zhang, H.M. Characterization of muscle development and gene expression in early embryos of chicken, quail, and their hybrids. Gene 2021, 768, 7. [Google Scholar] [CrossRef] [PubMed]
- Weng, K.Q.; Huo, W.R.; Li, Y.; Zhang, Y.; Zhang, Y.; Chen, G.H.; Xu, Q. Fiber characteristics and meat quality of different muscular tissues from slow- and fast-growing broilers. Poult. Sci 2022, 101, 8. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.T.; Kim, G.D.; Hwang, Y.H.; Ryu, Y.C. Control of fresh meat quality through manipulation of muscle fiber characteristics. Meat Sci. 2013, 95, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.R.; Weng, K.Q.; Li, Y.; Zhang, Y.; Zhang, Y.; Xu, Q.; Chen, G.H. Comparison of muscle fiber characteristics and glycolytic potential between slow- and fast-growing broilers. Poult. Sci 2022, 101, 9. [Google Scholar] [CrossRef]
- Tejeda, O.J.; Calderon, A.J.; Arana, J.A.; Meloche, K.J.; Starkey, J.D. Broiler chicken myofiber morphometrics and myogenic stem cell population heterogeneity. Poult. Sci 2019, 98, 4123–4130. [Google Scholar] [CrossRef]
- Li, H.F.; Shu, J.T.; Shan, Y.J.; Chen, W.F.; Song, C.; Xu, W.J. Myofiber development during embryonic to neonatal development in duck breeds differing in muscle growth rates. J. Integr. Agr. 2016, 15, 403–413. [Google Scholar] [CrossRef]
- Uni, Z.; Ferket, P.R.; Tako, E.; Kedar, O. In ovo feeding improves energy status of late-term chicken embryos. Poult. Sci 2005, 84, 764–770. [Google Scholar] [CrossRef]
- Foye, O.T.; Ferket, P.R.; Uni, Z. Ontogeny of energy and carbohydrate utilisation of the precocial avian embryo and hatchling. Avian Poult. Biol. Rev. 2007, 18, 93–101. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Christensen, V.L. Dietary carbohydrate level and glucose metabolism in turkey poults. Comp. Biochem. Physiol. A Comp. Physiol. 1991, 98, 347–350. [Google Scholar] [CrossRef]
- Liu, J.; Lei, Q.X.; Li, F.W.; Zhou, Y.; Gao, J.B.; Liu, W.; Han, H.X.; Cao, D.G. Dynamic Transcriptomic Analysis of Breast Muscle Development from the Embryonic to Post-hatching Periods in Chickens. Front. Genet. 2020, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Ban, Q.; Liang, Y.W.; Zhao, Z.S.; Liu, X.J.; Li, Q.F. Differential Expression Levels of Genes Related to Myogenesis During Embryogenesis of Quail and Chicken. Pak. Vet. J. 2013, 33, 317–320. [Google Scholar]
- Li, L.A.; Liu, H.H.; Xu, F.; Si, J.M.; Jia, J.; Wang, J.W. MyoD expression profile and developmental differences of leg and breast muscle in Peking duck (Anas platyrhynchos Domestica) during embryonic to neonatal stages. Micron 2010, 41, 847–852. [Google Scholar] [CrossRef]
- Fredette, B.J.; Landmesser, L.T. A reevaluation of the role of innervation in primary and secondary myogenesis in developing chick muscle. Dev. Biol. 1991, 143, 19–35. [Google Scholar] [CrossRef]
- Messina, G.; Cossu, G. The origin of embryonic and fetal myoblasts: A role of Pax3 and Pax7. Genes Dev. 2009, 23, 902–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, R.P.; Zhao, Y.M.; Li, Q.Q.; Yan, X.P.; Liu, J.Y.; Gou, H.; Li, L. Effects of Pax3 and Pax7 expression on muscle mass in the Pekin duck (Anas platyrhynchos domestica). Genet. Mol. Res. 2015, 14, 11495–11504. [Google Scholar] [CrossRef] [PubMed]
- Picard, C.A.; Marcelle, C. Two distinct muscle progenitor populations coexist throughout amniote development. Dev. Biol. 2013, 373, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.H.; Gi, G.G.; Tao, Z.Y.; Song, C.; Zhu, W.Q.; Song, W.T.; Li, H.F. Development of skeletal muscle and expression of myogenic regulatory factors during embryonic development in Jinding ducks (Anas platyrhynchos domestica). Poult. Sci 2014, 93, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhang, S.; Gilbert, E.R.; Siegel, P.B.; Zhu, Q.; Wong, E.A. Expression profiles of muscle genes in postnatal skeletal muscle in lines of chickens divergently selected for high and low body weight. Poult. Sci 2014, 93, 147–154. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Primer Sequence (5′→3′) | Product Length (bp) | Annealing Temperature (°C) | Accession No. |
|---|---|---|---|---|
| MyoD | F: ACTACAGCGGGGAGTCAGAT | 149 | 57 | NM_204214.3 |
| R: CCCATGCTTTGGGTCATTTGG | ||||
| Pax7 | F: TCGATTAGCCGTGTGCTACG | 108 | 59.2 | NM_205065.1 |
| R: GCCATCTATGCTGTGCTTGG | ||||
| GAPDH | F: GGGGAAAGTCATCCCTGAGC | 145 | 60 | NM_204305.1 |
| R: AGCAGCCTTCACTACCCTCT |
| Variable | Diameter (µm) | Cross-Sectional Area (µm2) | Density (N/mm2) | Breast Weight (g) | Embryo Weight (g) |
|---|---|---|---|---|---|
| MyoD | −0.601 | −0.638 | 0.976 ** | −0.822 * | −0.936 ** |
| Pax7 | 0.136 | 0.120 | 0.564 | −0.188 | −0.528 |
| Diameter (µm) | 0.997 ** | −0.687 | 0.898 ** | 0.690 | |
| Cross-sectional area (µm2) | −0.711 | 0.904 ** | 0.706 | ||
| Density (N/mm2) | −0.887 ** | −0.965 ** |
| Variable | Diameter (µm) | Cross-Sectional Area (µm2) | Density (N/mm2) | Leg Weight (g) | Embryo Weight (g) |
|---|---|---|---|---|---|
| MyoD | −0.799 * | −0.811 * | 0.952 ** | −0.866 * | −0.825 * |
| Pax7 | −0.284 | −0.314 | 0.453 | −0.564 | −0.435 |
| Diameter (µm) | 0.999 ** | −0.923 ** | 0.941 ** | 0.981 ** | |
| Cross-sectional area (µm2) | −0.930 ** | 0.949 ** | 0.985 ** | ||
| Density (N/mm2) | −0.939 ** | −0.940 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Yin, L.; Yuan, Y.; Tang, Y.; Lin, Z.; Liu, Y.; Yang, J. Developmental Characteristics of Skeletal Muscle during the Embryonic Stage in Chinese Yellow Quail (Coturnix japonica). Animals 2023, 13, 2317. https://doi.org/10.3390/ani13142317
Liu L, Yin L, Yuan Y, Tang Y, Lin Z, Liu Y, Yang J. Developmental Characteristics of Skeletal Muscle during the Embryonic Stage in Chinese Yellow Quail (Coturnix japonica). Animals. 2023; 13(14):2317. https://doi.org/10.3390/ani13142317
Chicago/Turabian StyleLiu, Li, Lingqian Yin, Yaohan Yuan, Yuan Tang, Zhongzhen Lin, Yiping Liu, and Jiandong Yang. 2023. "Developmental Characteristics of Skeletal Muscle during the Embryonic Stage in Chinese Yellow Quail (Coturnix japonica)" Animals 13, no. 14: 2317. https://doi.org/10.3390/ani13142317
APA StyleLiu, L., Yin, L., Yuan, Y., Tang, Y., Lin, Z., Liu, Y., & Yang, J. (2023). Developmental Characteristics of Skeletal Muscle during the Embryonic Stage in Chinese Yellow Quail (Coturnix japonica). Animals, 13(14), 2317. https://doi.org/10.3390/ani13142317