

Effect of a GnRH Antagonist on Anti-Müllerian Hormone and Follicular Population in Anestrous Bitches
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Treatment Protocol and Follow-Up
2.2. Blood Sampling and Hormone Measurements
2.3. Ovariectomies, and Gross Histological Evaluation of the Gonads
2.4. Statistical Analysis
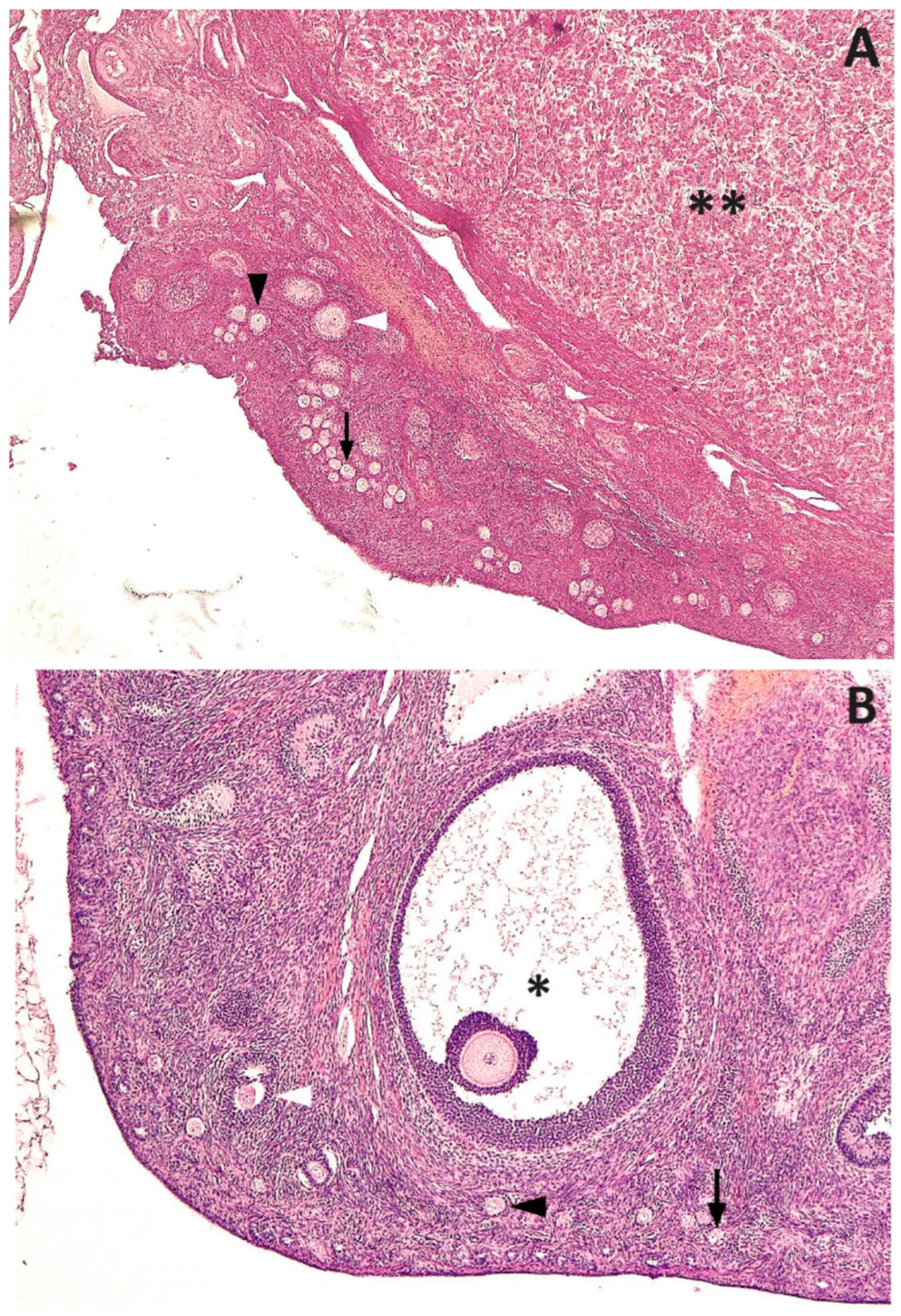
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gobello, C. Effects of GnRH Antagonists vs. Agonists in Domestic Carnivores, a Review. Reprod. Domest. Anim. 2012, 47 (Suppl. S6), 373–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiente, C.; Corrada, Y.; de la Sota, P.E.; Blanco, P.G.; Arias, D.; Gobello, C. Comparison of Two Doses of the GnRH Antagonist, Acyline, for Pregnancy Termination in Bitches. Reprod. Domest. Anim. 2009, 44 (Suppl. S2), 156–159. [Google Scholar] [CrossRef] [PubMed]
- Valiente, C.; Diaz, J.D.; Rosa, D.E.; Mattioli, G.; García Romero, G.; Gobello, C. Effect of a GnRH Antagonist on GnRH Agonist-Implanted Anestrous Bitches. Theriogenology 2009, 72, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Batista, P.R.; Blanco, P.G.; Gobello, C. Treatment of Canine Pyometra with the Gonadotropin-Releasing Hormone Antagonist Acyline: A Case Series. Top. Companion Anim. Med. 2015, 30, 25–27. [Google Scholar] [CrossRef]
- García Romero, G.; Valiente, C.; Aquilano, D.; Corrada, Y.; Gobello, C. Endocrine Effects of the GnRH Antagonist, Acyline, in Domestic Dogs. Theriogenology 2009, 71, 1234–1237. [Google Scholar] [CrossRef]
- Peng, P.; Mo, Y.; Yang, C.; Zhang, W.; Zhang, Y.; Liu, J.; Ye, C.; Yang, D. Effects of gonadotroph-releasing hormone analogues on follicle apoptosis in rats with chemotherapy-induced ovarian damage. Nan Fang Yi Ke Da Xue Xue Bao 2010, 30, 106–110. [Google Scholar]
- Ron-El, R.; Raziel, A.; Schachter, M.; Strassburger, D.; Kasterstein, E.; Friedler, S. Induction of Ovulation after GnRH Antagonists. Hum. Reprod. Update 2000, 6, 318–321. [Google Scholar] [CrossRef] [Green Version]
- Pelican, K.M.; Brown, J.L.; Wildt, D.E.; Ottinger, M.A.; Howard, J.G.; Pelican, K.M.; Brown, J.L.; Wildt, D.E.; Ottinger, M.A.; Howard, J.G. Ovarian suppression with the progestin levonorgestrel improves ovulation induction for artificial insemination in the domestic cat. Reprod. Fertil. Dev. 2003, 16, 131. [Google Scholar] [CrossRef]
- Visser, J.A.; de Jong, F.H.; Laven, J.S.E.; Themmen, A.P.N. Anti-Müllerian Hormone: A New Marker for Ovarian Function. Reproduction 2006, 131, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ireland, J.J.; Smith, G.W.; Scheetz, D.; Jimenez-Krassel, F.; Folger, J.K.; Ireland, J.L.H.; Mossa, F.; Lonergan, P.; Evans, A.C.O. Does Size Matter in Females? An Overview of the Impact of the High Variation in the Ovarian Reserve on Ovarian Function and Fertility, Utility of Anti-Müllerian Hormone as a Diagnostic Marker for Fertility and Causes of Variation in the Ovarian Reserve in Cattle. Reprod. Fertil. Dev. 2011, 23, 1–14. [Google Scholar] [CrossRef]
- Walter, B.; Feulner, H.; Otzdorff, C.; Klein, R.; Reese, S.; Meyer-Lindenberg, A. Changes in Anti-Müllerian Hormone Concentrations in Bitches throughout the Oestrous Cycle. Theriogenology 2019, 127, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, J.B.; Hansen, B.S.; Songsasen, N.; Travis, A.J.; Place, N.J. Anti-Müllerian Hormone in the Domestic Dog during the Anestrus to Oestrous Transition. Reprod. Domest. Anim. 2016, 51, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Themmen, A.P.N.; Kalra, B.; Visser, J.A.; Kumar, A.; Savjani, G.; de Gier, J.; Jaques, S. The Use of Anti-Müllerian Hormone as Diagnostic for Gonadectomy Status in Dogs. Theriogenology 2016, 86, 1467–1474. [Google Scholar] [CrossRef]
- de Gier, J.; Beijerink, N.J.; Kooistra, H.S.; Okkens, A.C. Physiology of the Canine Anoestrus and Methods for Manipulation of Its Length. Reprod. Domest. Anim. 2008, 43 (Suppl. S2), 157–164. [Google Scholar] [CrossRef] [PubMed]
- England, G.C.W.; Russo, M.; Freeman, S.L. Follicular Dynamics, Ovulation and Conception Rates in Bitches. Reprod. Domest. Anim. 2009, 44 (Suppl. S2), 53–58. [Google Scholar] [CrossRef] [PubMed]
- Concannon, P.W. Endocrinologic Control of Normal Canine Ovarian Function. Reprod. Domest. Anim. 2009, 44 (Suppl. S2), 3–15. [Google Scholar] [CrossRef]
- Arnold, E. Ovary and Uterus. In Textbook of Small Animal Surgery, 3rd ed.; Slatter, D., Ed.; Saunders RW: Philadelphia, PA, USA, 2002. [Google Scholar]
- Songsasen, N.; Fickes, A.; Pukazhenthi, B.S.; Wildt, D.E. Follicular Morphology, Oocyte Diameter and Localisation of Fibroblast Growth Factors in the Domestic Dog Ovary. Reprod. Domest. Anim. 2009, 44 (Suppl. S2), 65–70. [Google Scholar] [CrossRef] [Green Version]
- Gougeon, A.; Chainy, G.B. Morphometric Studies of Small Follicles in Ovaries of Women at Different Ages. J. Reprod. Fertil. 1987, 81, 433–442. [Google Scholar] [CrossRef]
- Marchetti, C.; Faya, M.; Priotto, M.; Grisolia-Romero, M.; Gobello, C. Long-term Reproductive Evaluation of a Bitch Neonatally Treated with a Gonadotropin-releasing Hormone Agonist. Vet. Rec. Case Rep. 2021, 9, e212. [Google Scholar] [CrossRef]
- Lunardon, N.T.; Silva-Santos, K.C.; Justino, R.C.; Dessunti, G.T.; Seneda, M.M.; Martins, M.I.M. Population Estimate of the Preantral Follicles and Frequency of Multioocyte Follicles in Prepubertal and Adult Bitches. Theriogenology 2015, 83, 1015–1020. [Google Scholar] [CrossRef]
- Jivago, J.L.P.R.; Paulini, F.; Silva, R.C.; Araujo, M.S.; Marinho, A.P.S.; Lucci, C.M. Cryopreservation and Characterization of Canine Preantral Follicles. Cryobiology 2018, 81, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Zysling, D.A.; Park, S.-U.; McMillan, E.L.; Place, N.J. Photoperiod-Gonadotropin Mismatches Induced by Treatment with Acyline or FSH in Siberian Hamsters: Impacts on Ovarian Structure and Function. Reproduction 2012, 144, 603–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, B.K.; Dobson, H.; Scaramuzzi, R.J. Ovarian Function in Ewes Made Hypogonadal with GnRH Antagonist and Stimulated with FSH in the Presence or Absence of Low Amplitude LH Pulses. J. Endocrinol. 1998, 156, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Krassel, F.; Ireland, J.L.H.; Kronemeyer, C.; Wilson-Alvarado, A.; Ireland, J.J. Development of the “Waveless” Bovine Model. Anim. Reprod. Sci. 2018, 195, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Place, N.J.; Crosier, A.E.; Comizzoli, P.; Nagashima, J.B.; Haefele, H.; Schmidt-Küntzel, A.; Marker, L.L. Age-Associated and Deslorelin-Induced Declines in Serum Anti-Müllerian Hormone Concentrations in Female Cheetahs, Acinonyx Jubatus. Gen. Comp. Endocrinol. 2017, 250, 54–57. [Google Scholar] [CrossRef]
- Ferré-Dolcet, L.; Ferro, S.; Contiero, B.; Fontaine, C.; Badon, T.; Gelli, D.; Romagnoli, S. Clinical Use of Anti-Müllerian Hormone to Monitor Resumption of Ovarian Activity Following Removal of a 4.7 Mg Deslorelin Implant in Queens. Vet. Res. Commun. 2022, 46, 917–924. [Google Scholar] [CrossRef]
- Hariton, E.; Shirazi, T.N.; Douglas, N.C.; Hershlag, A.; Briggs, S.F. Anti-Müllerian Hormone Levels among Contraceptive Users: Evidence from a Cross-Sectional Cohort of 27,125 Individuals. Am. J. Obs. Gynecol. 2021, 225, 515.e1–515.e10. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Mean ± SEM | |
|---|---|
| Body weight (kg) | 8.37 ± 0.9 |
| Ovarian volume (cm2) | 0.5 ± 0.07 |
| Ovarian weight (g) | 0.4 ± 0.07 |
| Gonado-somatic index (%) | 0.01 ± 0.00 |
| Mean ± SEM | % | |
|---|---|---|
| Primordial | 80,931.08 ± 23,107.77 | 84.13 |
| Primary | 10,927.42 ± 2536.89 | 11.36 |
| Secondary | 7505.95 ± 3263.16 | 7.80 |
| Antral | 5.55 ± 5.55 | 0.01 |
| Total | 96,200.10 ± 26,125.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchetti, C.; Grisolia Romero, M.; Priotto, M.; Faya, M.; Gobello, C. Effect of a GnRH Antagonist on Anti-Müllerian Hormone and Follicular Population in Anestrous Bitches. Animals 2023, 13, 2511. https://doi.org/10.3390/ani13152511
Marchetti C, Grisolia Romero M, Priotto M, Faya M, Gobello C. Effect of a GnRH Antagonist on Anti-Müllerian Hormone and Follicular Population in Anestrous Bitches. Animals. 2023; 13(15):2511. https://doi.org/10.3390/ani13152511
Chicago/Turabian StyleMarchetti, Cynthia, Mariela Grisolia Romero, Marcelo Priotto, Marcela Faya, and Cristina Gobello. 2023. "Effect of a GnRH Antagonist on Anti-Müllerian Hormone and Follicular Population in Anestrous Bitches" Animals 13, no. 15: 2511. https://doi.org/10.3390/ani13152511