
Melatonin Supplementation during In Vitro Maturation of Porcine Oocytes Alleviates Oxidative Stress and Endoplasmic Reticulum Stress Induced by Imidacloprid Exposure
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Porcine Oocyte Collection and IVM
2.2. Imidacloprid and Melatonin Treatments
2.3. Oocyte Parthenogenetic Activation and In Vitro Culturing
2.4. Intracellular ROS and GSH Level Assays
2.5. Measurement of Cathepsin B Activity
2.6. Assessment of Blastocyst Total Cell Numbers
2.7. Relative Gene Expression Measurement Using Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.8. Statistical Analysis
3. Results
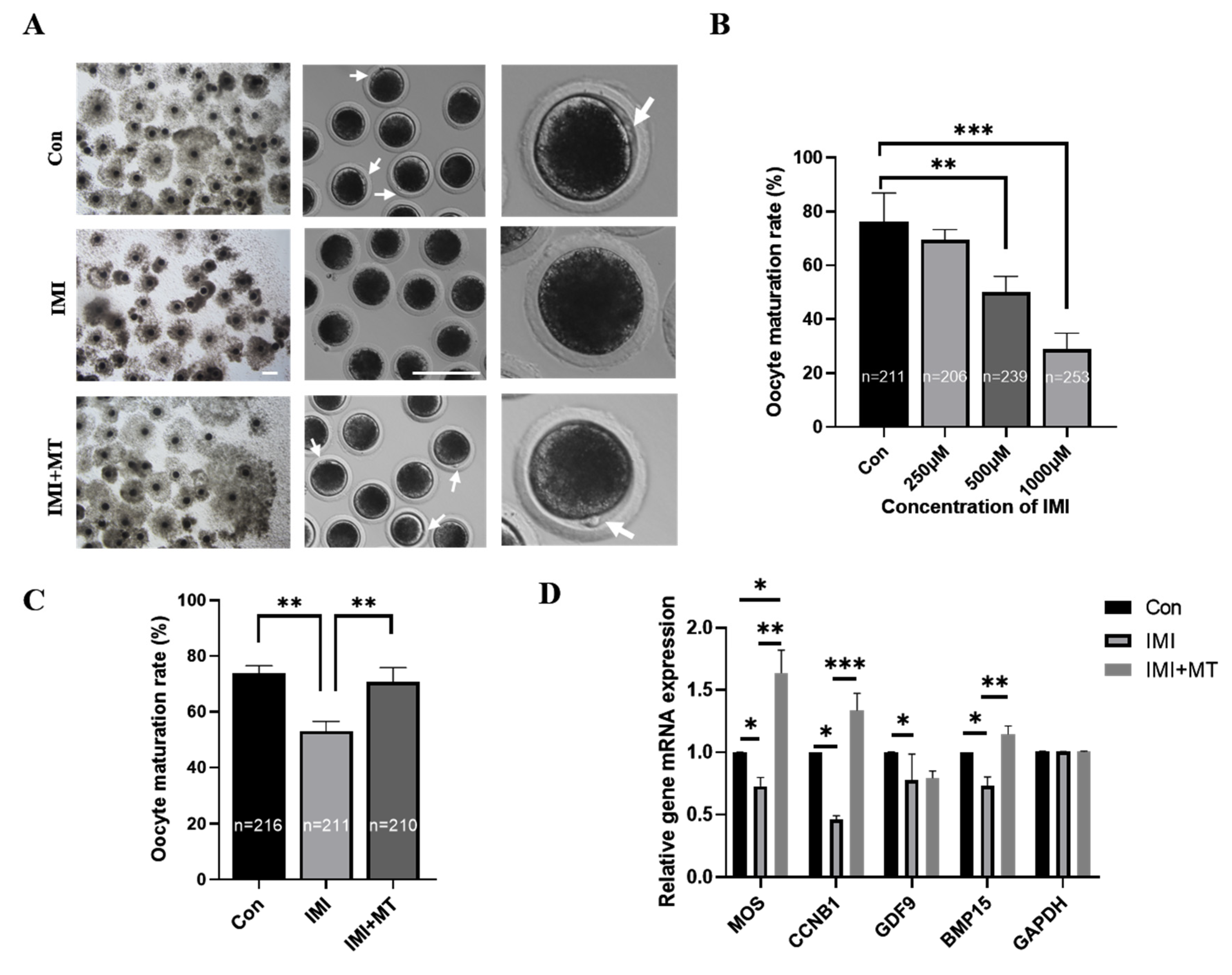
3.1. MT Alleviates Maturation Rate and Oocyte Quality Reductions in IMI-Exposed Porcine Oocytes
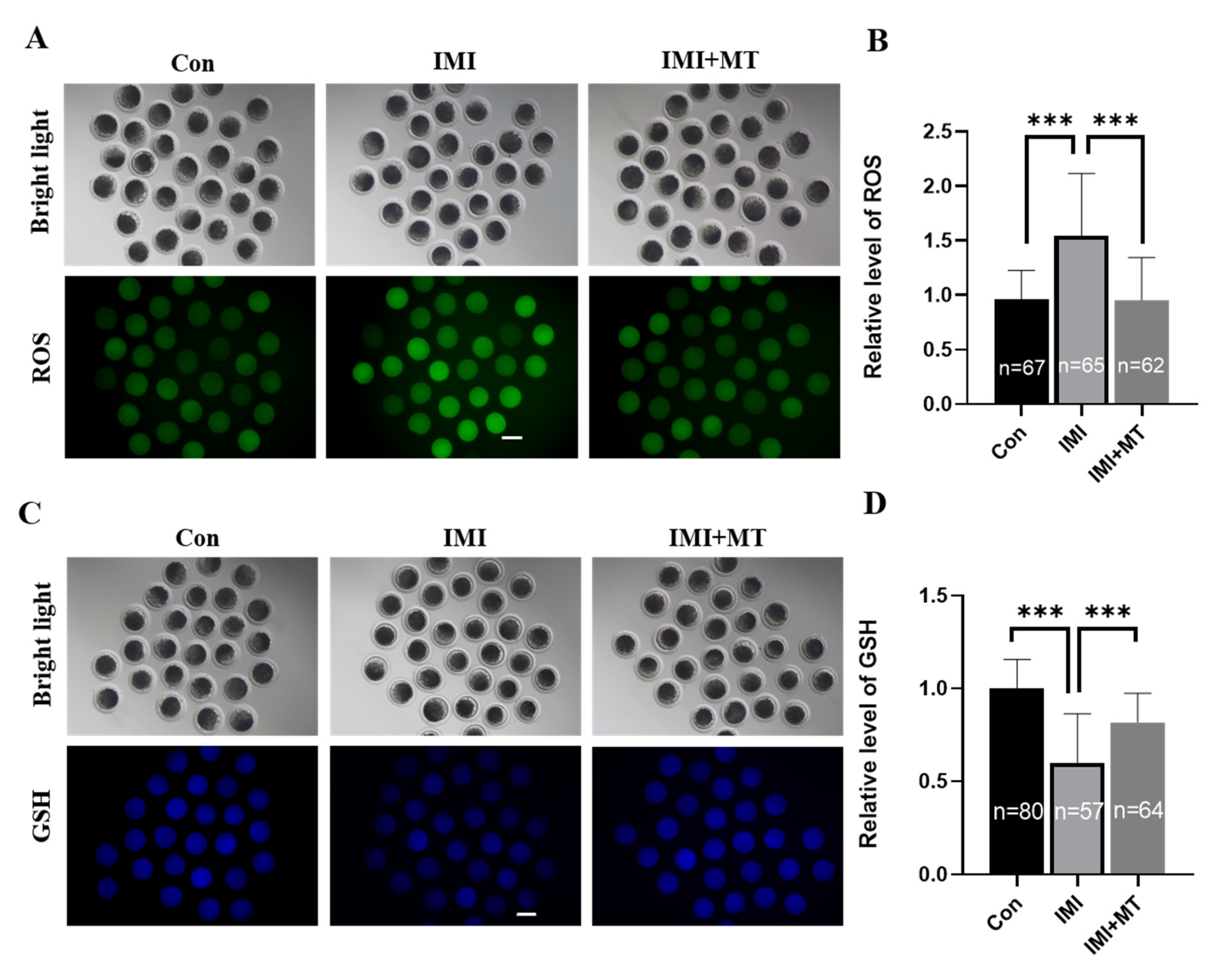
3.2. MT Reduces Oxidative Stress and Endoplasmic Reticulum Stress in IMI-Exposed Porcine Oocytes
3.3. MT Reduces Apoptosis in IMI-Exposed Porcine Oocytes
3.4. MT Promotes Blastocyst Formation in IMI-Exposed Porcine Embryos after Parthenogenetic Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hatırnaz, Ş.; Ata, B.; Hatırnaz, E.S.; Dahan, M.H.; Tannus, S.; Tan, J.; Tan, S.L. Oocyte in vitro maturation: A sytematic review. Turk. J. Obstet. Gynecol. 2018, 15, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Rhind, S.M.; Evans, N.P.; Bellingham, M.; Sharpe, R.M.; Cotinot, C.; Mandon-Pepin, B.; Loup, B.; Sinclair, K.D.; Lea, R.G.; Pocar, P.; et al. Effects of environmental pollutants on the reproduction and welfare of ruminants. Anim. Int. J. Anim. Biosci. 2010, 4, 1227–1239. [Google Scholar] [CrossRef]
- Jiang, W.J.; Liu, W.; Li, Y.H.; Jiang, H.; Xu, Y.N.; Kim, N.H. Citrinin impairs pig oocyte maturation by inducing oxidative stress and apoptosis. Toxicon 2022, 205, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.I.; Hassold, T.J.; Hunt, P.A. Human aneuploidy: Mechanisms and new insights into an age-old problem. Nat. Rev. Genet. 2012, 13, 493–504. [Google Scholar] [CrossRef]
- Vieira, C.E.D.; Pérez, M.R.; Acayaba, R.D.; Raimundo, C.C.M.; Dos Reis Martinez, C.B. DNA damage and oxidative stress induced by imidacloprid exposure in different tissues of the Neotropical fish Prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef]
- Wei, F.; Wang, D.; Li, H.; Xia, P.; Ran, Y.; You, J. Toxicogenomics provides insights to toxicity pathways of neonicotinoids to aquatic insect, Chironomus dilutus. Environ. Pollut. 2020, 260, 114011. [Google Scholar] [CrossRef]
- Lv, Y.; Bing, Q.; Lv, Z.; Xue, J.; Li, S.; Han, B.; Yang, Q.; Wang, X.; Zhang, Z. Imidacloprid-induced liver fibrosis in quails via activation of the TGF-β1/Smad pathway. Sci. Total Environ. 2020, 705, 135915. [Google Scholar] [CrossRef]
- Khandia, R.; Pathe, C.S.; Vishwakarma, P.; Dhama, K.; Munjal, A. Evaluation of the ameliorative effects of Phyllanthus niruri on the deleterious insecticide imidacloprid in the vital organs of chicken embryos. J. Ayurveda Integr. Med. 2020, 11, 495–501. [Google Scholar] [CrossRef]
- Abou-Donia, M.B.; Goldstein, L.B.; Bullman, S.; Tu, T.; Khan, W.A.; Dechkovskaia, A.M.; Abdel-Rahman, A.A. Imidacloprid induces neurobehavioral deficits and increases expression of glial fibrillary acidic protein in the motor cortex and hippocampus in offspring rats following in utero exposure. J. Toxicol. Environ. Health Part A 2008, 71, 119–130. [Google Scholar] [CrossRef]
- Abu Zeid, E.H.; Alam, R.T.M.; Ali, S.A.; Hendawi, M.Y. Dose-related impacts of imidacloprid oral intoxication on brain and liver of rock pigeon (Columba livia domestica), residues analysis in different organs. Ecotoxicol. Environ. Saf. 2019, 167, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; Wang, M.; Chen, A.; Zhang, C.; Lin, L.; Zhang, Z.; Chen, A. Protective effect of caffeic acid phenethyl ester against imidacloprid-induced hepatotoxicity by attenuating oxidative stress, endoplasmic reticulum stress, inflammation and apoptosis. Pestic. Biochem. Physiol. 2020, 164, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Duan, J.; Chen, L.; Wang, Y.; Qin, Q.; Dang, X.; Zhou, Z. Melatonin enhances the antioxidant capacity to rescue the honey bee Apis mellifera from the ecotoxicological effects caused by environmental imidacloprid. Ecotoxicol. Environ. Saf. 2022, 239, 113622. [Google Scholar] [CrossRef]
- Chern, C.M.; Liao, J.F.; Wang, Y.H.; Shen, Y.C. Melatonin ameliorates neural function by promoting endogenous neurogenesis through the MT2 melatonin receptor in ischemic-stroke mice. Free Radic. Biol. Med. 2012, 52, 1634–1647. [Google Scholar] [CrossRef]
- Xu, C.L.; Tan, Q.Y.; Yang, H.; Li, C.Y.; Wu, Z.; Ma, Y.F. Melatonin enhances spermatogonia activity through promoting KIAA1429-mediated m6A deposition to activate the PI3K/AKT signaling. Reprod. Biol. 2022, 22, 100681. [Google Scholar] [CrossRef]
- Kazemzadeh, S.; Mohammadpour, S.; Madadi, S.; Babakhani, A.; Shabani, M.; Khanehzad, M. Melatonin in cryopreservation media improves transplantation efficiency of frozen-thawed spermatogonial stem cells into testes of azoospermic mice. Stem Cell Res. Ther. 2022, 13, 346. [Google Scholar] [CrossRef] [PubMed]
- Lan, M.; Zhang, Y.; Wan, X.; Pan, M.H.; Xu, Y.; Sun, S.C. Melatonin ameliorates ochratoxin A-induced oxidative stress and apoptosis in porcine oocytes. Environ. Pollut. 2020, 256, 113374. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Jiang, H.; Gao, Q.; Li, Y.H.; Xu, Y.N.; Kim, N.H. Melatonin alleviates defects induced by zearalenone during porcine embryo development. Theriogenology 2020, 151, 66–73. [Google Scholar] [CrossRef]
- Liang, S.; Jin, Y.X.; Yuan, B.; Zhang, J.B.; Kim, N.H. Melatonin enhances the developmental competence of porcine somatic cell nuclear transfer embryos by preventing DNA damage induced by oxidative stress. Sci. Rep. 2017, 7, 11114. [Google Scholar] [CrossRef]
- Nikmard, F.; Hosseini, E.; Bakhtiyari, M.; Ashrafi, M.; Amidi, F.; Aflatoonian, R. Effects of melatonin on oocyte maturation in PCOS mouse model. Anim. Sci. J. Nihon Chikusan Gakkaiho 2017, 88, 586–592. [Google Scholar] [CrossRef]
- Wang, T.; Gao, Y.Y.; Chen, L.; Nie, Z.W.; Cheng, W.; Liu, X.; Schatten, H.; Zhang, X.; Miao, Y.L. Melatonin prevents postovulatory oocyte aging and promotes subsequent embryonic development in the pig. Aging 2017, 9, 1552–1564. [Google Scholar] [CrossRef]
- Zhao, Z.; Yang, L.; Zhang, D.; Zheng, Z.; Li, N.; Li, Q.; Cui, M. Elevation of MPF and MAPK gene expression, GSH content and mitochondrial distribution quality induced by melatonin promotes porcine oocyte maturation and development in vitro. PeerJ 2020, 8, e9913. [Google Scholar] [CrossRef]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.M.; Tian, J.H.; Liu, G.S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar] [CrossRef]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Lee, B.C. Synergistic effects of resveratrol and melatonin on in vitro maturation of porcine oocytes and subsequent embryo development. Theriogenology 2018, 114, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Field, L.M. Neonicotinoids. Curr. Biol. 2018, 28, R772–R773. [Google Scholar] [CrossRef]
- Wang, J.; Yin, R.; Liu, Y.; Wang, B.; Wang, N.; Xiao, P.; Xiao, T.; Hirai, H. Meta-analysis of neonicotinoid insecticides in global surface waters. Environ. Sci. Pollut. Res. Int. 2022, 30, 1039–1047. [Google Scholar] [CrossRef]
- Harada, K.H.; Tanaka, K.; Sakamoto, H.; Imanaka, M.; Niisoe, T.; Hitomi, T.; Kobayashi, H.; Okuda, H.; Inoue, S.; Kusakawa, K.; et al. Biological Monitoring of Human Exposure to Neonicotinoids Using Urine Samples, and Neonicotinoid Excretion Kinetics. PLoS ONE 2016, 11, e0146335. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Z.; Chang, C.H.; Lou, J.L.; Zhao, M.R.; Lu, C. Potential human exposures to neonicotinoid insecticides: A review. Environ. Pollut. 2018, 236, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.A.; Lehmler, H.J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; LeFevre, G.H.; Peeples, T.L.; Poch, M.C.; LaDuca, L.E.; et al. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health. Environ. Sci. Process. Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef]
- Zhao, G.P.; Li, J.W.; Yang, F.W.; Yin, X.F.; Ren, F.Z.; Fang, B.; Pang, G.F. Spermiogenesis toxicity of imidacloprid in rats, possible role of CYP3A4. Chemosphere 2021, 282, 131120. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razik, R.K.; Mosallam, E.M.; Hamed, N.A.; Badawy, M.E.I.; Abo-El-Saad, M.M. Testicular deficiency associated with exposure to cypermethrin, imidacloprid, and chlorpyrifos in adult rats. Environ. Toxicol. Pharmacol. 2021, 87, 103724. [Google Scholar] [CrossRef]
- Kapoor, U.; Srivastava, M.K.; Srivastava, L.P. Toxicological impact of technical imidacloprid on ovarian morphology, hormones and antioxidant enzymes in female rats. Food Chem. Toxicol. 2011, 49, 3086–3089. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Y.; Hu, L.L.; Xing, C.H.; Lu, X.; Sun, S.C.; Wei, Y.X.; Ren, Y.P. Acrylamide Exposure Destroys the Distribution and Functions of Organelles in Mouse Oocytes. Front. Cell Dev. Biol. 2022, 10, 834964. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, C.H.; Chen, S.; Sun, S.C. Zearalenone exposure impairs organelle function during porcine oocyte meiotic maturation. Theriogenology 2022, 177, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.O.; Marinho, L.; da Rosa, P.; De Cesaro, M.P.; Lunardelli, P.A.; Silva-Santos, K.C.; Basso, A.C.; Bordignon, V.; Seneda, M.M. Molecular characteristics of granulosa and cumulus cells and oocyte competence in Nelore cows with low and high numbers of antral follicles. Reprod. Domest. Anim. 2018, 53, 921–929. [Google Scholar] [CrossRef]
- Schoevers, E.J.; Colenbrander, B.; Roelen, B.A. Developmental stage of the oocyte during antral follicle growth and cumulus investment determines in vitro embryo development of sow oocytes. Theriogenology 2007, 67, 1108–1122. [Google Scholar] [CrossRef]
- Solak, K.A.; Santos, R.R.; van den Berg, M.; Blaauboer, B.J.; Roelen, B.A.; van Duursen, M.B. Naringenin (NAR) and 8-prenylnaringenin (8-PN) reduce the developmental competence of porcine oocytes in vitro. Reprod. Toxicol. 2014, 49, 1–11. [Google Scholar] [CrossRef]
- Luo, D.; Zhang, J.B.; Li, S.P.; Liu, W.; Yao, X.R.; Guo, H.; Jin, Z.L.; Jin, Y.X.; Yuan, B.; Jiang, H.; et al. Imperatorin Ameliorates the Aging-Associated Porcine Oocyte Meiotic Spindle Defects by Reducing Oxidative Stress and Protecting Mitochondrial Function. Front. Cell Dev. Biol. 2020, 8, 592433. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Liang, S.; Jin, Y.X.; Zhang, M.J.; Zhang, J.B.; Kim, N.H. Toxic effects of atrazine on porcine oocytes and possible mechanisms of action. PLoS ONE 2017, 12, e0179861. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Pérez, T.; Hayward, D.; Holder, J.; Gruneberg, U.; Barr, F.A. MAD1-dependent recruitment of CDK1-CCNB1 to kinetochores promotes spindle checkpoint signaling. J. Cell Biol. 2019, 218, 1108–1117. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, T.; Lan, M.; Zang, X.W.; Li, Y.L.; Cui, X.S.; Kim, N.H.; Sun, S.C. Melatonin protects oocytes from MEHP exposure-induced meiosis defects in porcine. Biol. Reprod. 2018, 98, 286–298. [Google Scholar] [CrossRef]
- Hao, T.; Xu, X.; Hao, H.; Du, W.; Pang, Y.; Zhao, S.; Zou, H.; Yang, S.; Zhu, H.; Yang, Y.; et al. Melatonin improves the maturation and developmental ability of bovine oocytes by up-regulating GJA4 to enhance gap junction intercellular communication. Reprod. Fertil. Dev. 2021, 33, 760–771. [Google Scholar] [CrossRef]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef]
- Jeong, P.S.; Lee, S.; Park, S.H.; Kim, M.J.; Kang, H.G.; Nanjidsuren, T.; Son, H.C.; Song, B.S.; Koo, D.B.; Sim, B.W.; et al. Butylparaben Is Toxic to Porcine Oocyte Maturation and Subsequent Embryonic Development Following In Vitro Fertilization. Int. J. Mol. Sci. 2020, 21, 3692. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Falcone, T.; Mohamed, M.S.; Aleem, A.A.; Sharma, R.K.; Worley, S.E.; Thornton, J.; Agarwal, A. Differential growth of human embryos in vitro: Role of reactive oxygen species. Fertil. Steril. 2004, 82, 593–600. [Google Scholar] [CrossRef]
- Hu, Y.; Betzendahl, I.; Cortvrindt, R.; Smitz, J.; Eichenlaub-Ritter, U. Effects of low O2 and ageing on spindles and chromosomes in mouse oocytes from pre-antral follicle culture. Hum. Reprod. 2001, 16, 737–748. [Google Scholar] [CrossRef] [PubMed]
- De Matos, D.G.; Furnus, C.C.; Moses, D.F.; Baldassarre, H. Effect of cysteamine on glutathione level and developmental capacity of bovine oocyte matured in vitro. Mol. Reprod. Dev. 1995, 42, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Martins-Gomes, C.; Ferreira, S.S.; Souto, E.B.; Andreani, T. Molecular Physicochemical Properties of Selected Pesticides as Predictive Factors for Oxidative Stress and Apoptosis-Dependent Cell Death in Caco-2 and HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 8107. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Miao, Z.; Wang, S.; Wu, H.; Xu, S. Exposure to imidacloprid induce oxidative stress, mitochondrial dysfunction, inflammation, apoptosis and mitophagy via NF-kappaB/JNK pathway in grass carp hepatocytes. Fish Shellfish Immunol. 2022, 120, 674–685. [Google Scholar] [CrossRef]
- Baihetiyaer, B.; Jiang, N.; Li, X.; He, B.; Wang, J.; Fan, X.; Sun, H.; Yin, X. Oxidative stress and gene expression induced by biodegradable microplastics and imidacloprid in earthworms (Eisenia fetida) at environmentally relevant concentrations. Environ. Pollut. 2023, 323, 121285. [Google Scholar] [CrossRef] [PubMed]
- Nikmard, F.; Hosseini, E.; Bakhtiyari, M.; Ashrafi, M.; Amidi, F.; Aflatoonian, R. The boosting effects of melatonin on the expression of related genes to oocyte maturation and antioxidant pathways: A polycystic ovary syndrome- mouse model. J. Ovarian Res. 2022, 15, 11. [Google Scholar] [CrossRef]
- El-Sheikh, M.; Mesalam, A.A.; Kang, S.M.; Joo, M.D.; Soliman, S.S.; Khalil, A.A.K.; Ahn, M.J.; Kong, I.K. Modulation of Apoptosis and Autophagy by Melatonin in Juglone-Exposed Bovine Oocytes. Animals 2023, 13, 1475. [Google Scholar] [CrossRef]
- Xing, C.H.; Wang, Y.; Liu, J.C.; Pan, Z.N.; Zhang, H.L.; Sun, S.C.; Zhang, Y. Melatonin reverses mitochondria dysfunction and oxidative stress-induced apoptosis of Sudan I-exposed mouse oocytes. Ecotoxicol. Environ. Saf. 2021, 225, 112783. [Google Scholar] [CrossRef]
- Kato, H.; Nishitoh, H. Stress responses from the endoplasmic reticulum in cancer. Front. Oncol. 2015, 5, 93. [Google Scholar] [CrossRef]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Qi, Z.; Chen, L. Endoplasmic Reticulum Stress and Autophagy. Adv. Exp. Med. Biol. 2019, 1206, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Guzel, E.; Arlier, S.; Guzeloglu-Kayisli, O.; Tabak, M.S.; Ekiz, T.; Semerci, N.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Endoplasmic Reticulum Stress and Homeostasis in Reproductive Physiology and Pathology. Int. J. Mol. Sci. 2017, 18, 792. [Google Scholar] [CrossRef]
- Fang, X.; Han, Q.; Li, S.; Luo, A. Melatonin attenuates spatial learning and memory dysfunction in developing rats by suppressing isoflurane-induced endoplasmic reticulum stress via the SIRT1/Mfn2/PERK signaling pathway. Heliyon 2022, 8, e10326. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.Z.; Cai, H.; He, C.; Yang, D.H.; Sun, J.; He, W.L.; Li, B.L.; Hua, J.L.; Peng, S. Melatonin relieves heat-induced spermatocyte apoptosis in mouse testes by inhibition of ATF6 and PERK signaling pathways. Zool. Res. 2021, 42, 514–524. [Google Scholar] [CrossRef]
- Zhou, R.; Ma, Y.; Tao, Z.; Qiu, S.; Gong, Z.; Tao, L.; Zhu, Y. Melatonin Inhibits Glucose-Induced Apoptosis in Osteoblastic Cell Line Through PERK-eIF2α-ATF4 Pathway. Front. Pharmacol. 2020, 11, 602307. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wei, Y.; Lu, Q.; Qin, D.; Zhang, M.; Du, X.; Xu, W.; Yu, X.; He, C.; Li, N.; et al. Melatonin alleviates LPS-induced endoplasmic reticulum stress and inflammation in spermatogonial stem cells. J. Cell. Physiol. 2021, 236, 3536–3551. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, J.; Zhao, Q.; Chen, Q.; Sun, Y.; Jin, Y.; Wu, J. Melatonin Induces Anti-Inflammatory Effects to Play a Protective Role via Endoplasmic Reticulum Stress in Acute Pancreatitis. Cell. Physiol. Biochem. 2016, 40, 1094–1104. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences 5′–3′ | Product Size (bp) | Accession Number |
|---|---|---|---|
| GAPDH | F: TTCCACGGCACAGTCAAG | 117 | NM_001206359.1 |
| R: ATACTCAGCACCAGCATCG | |||
| MOS | F: GGTGGTGGCCTACAATCTCC | 136 | NM_001113219.1 |
| R: TCAGCTTGTAGAGCGCGAAG | |||
| CCNB1 | F: CCAACTGGTTGGTGTCACTG | 195 | NM_001170768.1 |
| R: GCTCTCCGAAGAAAATGCAG | |||
| BMP15 | F: ATGCTGGAGTTGTACCAGCG | 87 | NM_001005155.2 |
| R: CTGAGAGGCCTTGCTCCATT | |||
| GDF9 | F: CCCCAAAGCCAACAGAAGTCA R: TGATGGAAGGGTTCCTGTCACC | 85 | NM_001001909.1 |
| CAT | F: AACTGTCCCTTCCGTGCTA R: CCTGGGTGACATTATCTTCG | 83 | XM_021081498.1 |
| SIRT1 | F: GAGAAGGAAACAATGGGCCG R: ACCAAACAGAAGGTTATCTCGGT | 150 | NM_001145750.2 |
| SOD1 | F: CAAAGGATCAAGAGAGGCACG R: CGAGAGGGCGATCACAGAAT | 84 | NM_001190422.1 |
| BAX | F: GCTTCAGGGTTTCATCCAGGATCG R: ACTCGCTCAACTTCTTGGTAGATC | 107 | XM_003127290.5 |
| BCL2 | F: GGATAACGGGAGGCTGGGATG R: TTATGGCCCAGATAGGCACC | 148 | XM_021099593.1 |
| CASPASE-3 | F: TGTGGGATTGAGACGGACAG R: TTTCGCCAGGAATAGTAACCAGG | 116 | NM_214131.1 |
| GRP78 | F: CGGAGGAGGAGGACAAGAAGGAG R: ATATGACGGCGTGATGCGGTTG | 143 | XM_001927795.7 |
| IRE1 | F: ACCGTGGTGTCTCAGGATGTGG R: CCAGCCAATGAGCAGGAAGGTG | 126 | XM_005668695.3 |
| JNK | F: CTCGCTACTACAGAGCACCTG R: TTCTCCCATAATGCACCCCAC | 85 | XM_021073087.1 |
| XBP1 | F: GGAGTTAAGACAGCGCTTGG R: GAGATGTTCTGGAGGGGTGA | 142 | NM_001271738.1 |
| CHOP | F: TCTGGCTTGGCTGACTGAGGAG | 139 | NM_001144845.1 |
| R: TTTCCGTTTCCTGGGTCTTCTTTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Wang, X.-Q.; Liu, R.-P.; Li, Y.-H.; Yao, X.-R.; Kim, N.-H.; Xu, Y.-N. Melatonin Supplementation during In Vitro Maturation of Porcine Oocytes Alleviates Oxidative Stress and Endoplasmic Reticulum Stress Induced by Imidacloprid Exposure. Animals 2023, 13, 2596. https://doi.org/10.3390/ani13162596
Wang J, Wang X-Q, Liu R-P, Li Y-H, Yao X-R, Kim N-H, Xu Y-N. Melatonin Supplementation during In Vitro Maturation of Porcine Oocytes Alleviates Oxidative Stress and Endoplasmic Reticulum Stress Induced by Imidacloprid Exposure. Animals. 2023; 13(16):2596. https://doi.org/10.3390/ani13162596
Chicago/Turabian StyleWang, Jing, Xin-Qin Wang, Rong-Ping Liu, Ying-Hua Li, Xue-Rui Yao, Nam-Hyung Kim, and Yong-Nan Xu. 2023. "Melatonin Supplementation during In Vitro Maturation of Porcine Oocytes Alleviates Oxidative Stress and Endoplasmic Reticulum Stress Induced by Imidacloprid Exposure" Animals 13, no. 16: 2596. https://doi.org/10.3390/ani13162596