
Optimizing Feeding Strategies for Growing Rabbits: Impact of Timing and Amount on Health and Circadian Rhythms

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
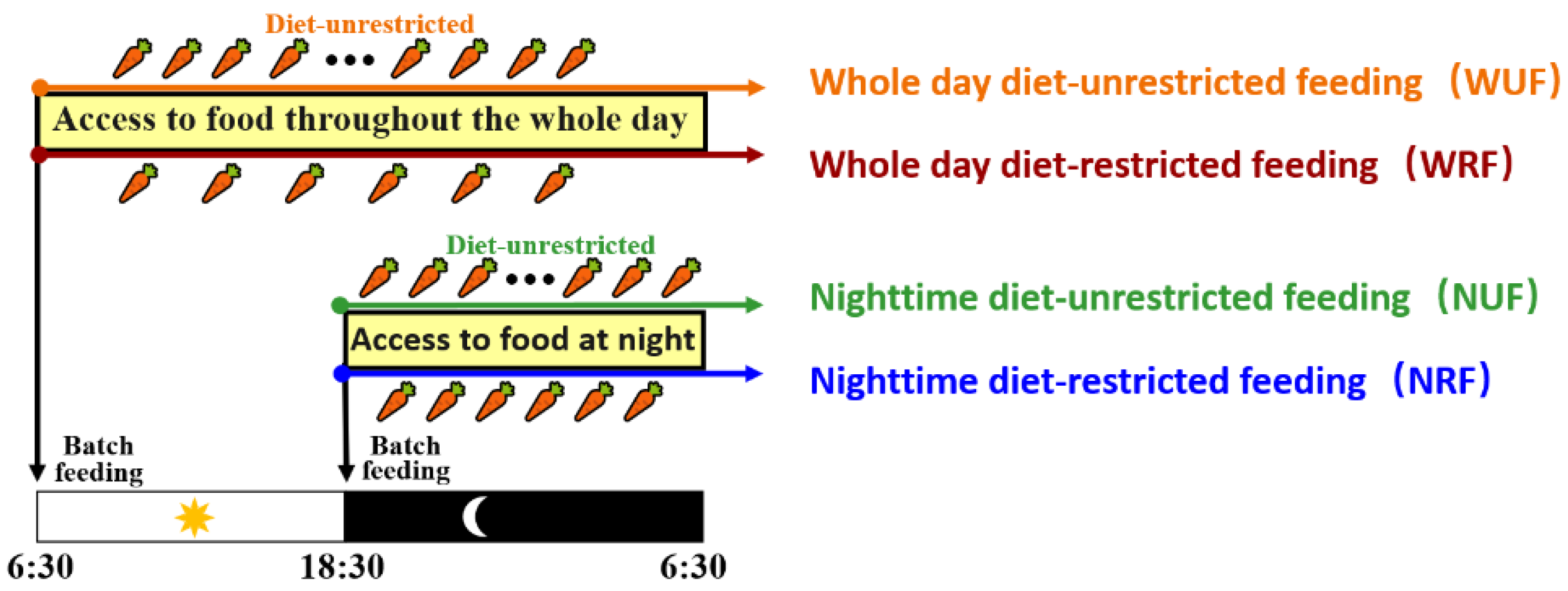
2.1. Experimental Animals and Feeding Management
2.2. Measurement of Production Performance Indicators
2.3. Experimental Environment, Behavior and Body Temperature Monitoring
2.4. Statistical Analysis
3. Results
3.1. Impact of Feeding Amount and Feeding Time on Rabbit Performance
3.2. Impact of Feeding Amount and Feeding Time on the Health of Rabbits
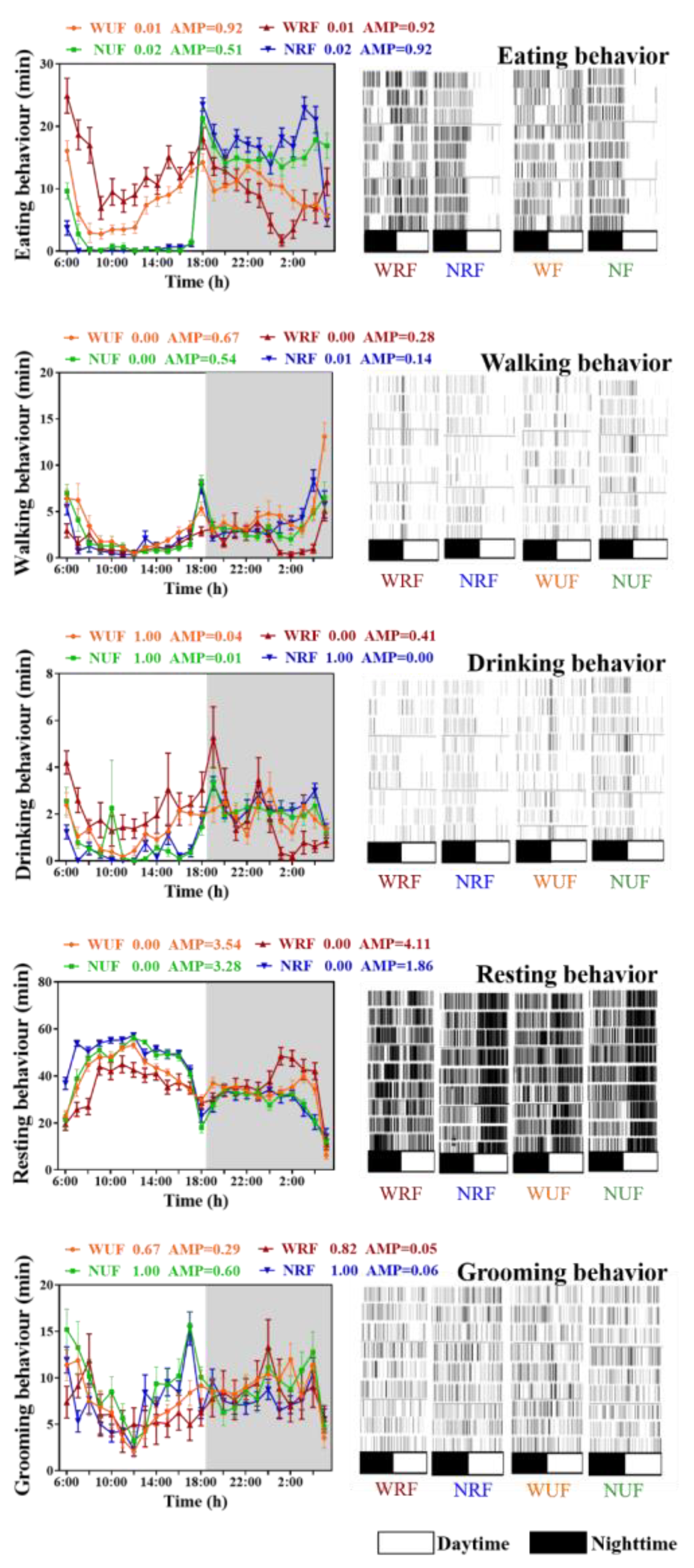
3.3. Impact of Feeding Amount and Feeding Time on Behavioral Rhythm of Rabbits
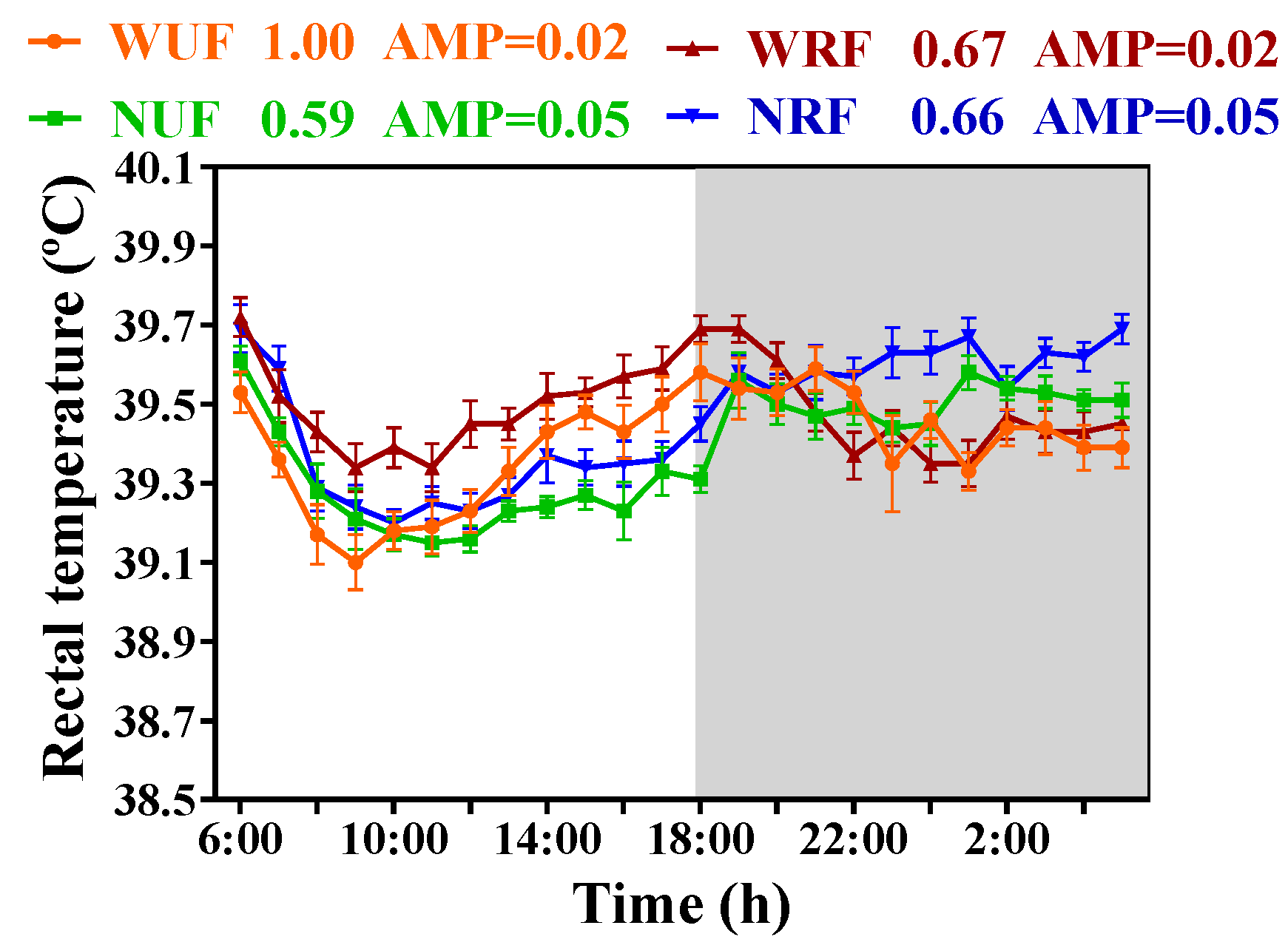
3.4. Impact of Feeding Amount and Feeding Time on Rectal Temperature of Rabbits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Erp, R.; de Vries, S.; van Kempen, T.; Den Hartog, L.A.; Gerrits, W. Circadian misalignment imposed by nocturnal feeding tends to increase fat deposition in pigs. Br. J. Nutr. 2020, 123, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Salfer, I.J.; Harvatine, K.J. Night-restricted feeding of dairy cows modifies daily rhythms of feed intake, milk synthesis and plasma metabolites compared with day-restricted feeding. Br. J. Nutr. 2020, 123, 849–858. [Google Scholar] [CrossRef]
- Guo, Y.; Yao, C.Y.; Wang, Q.J.; Li, Q.; Liu, Z.Y.; Zhang, K.H.; Geng, S.X.; She, D.Y.; Wu, Z.H. Effects of eating time on behavioral rhythm and performance of pigs under an intelligent electronic feeding system. Trans. CSAE 2022, 38, 206–214. [Google Scholar]
- Reznick, J.; Preston, E.; Wilks, D.L.; Beale, S.M.; Turner, N.; Cooney, G.J. Altered feeding differentially regulates circadian rhythms and energy metabolism in liver and muscle of rats. Biochim. Biophys. Acta 2013, 1832, 228–238. [Google Scholar] [CrossRef]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell. Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Cisse, Y.M.; Borniger, J.C.; Lemanski, E.; Walker, W.N.; Nelson, R.J. Time-restricted feeding alters the innate immune response to bacterial endotoxin. J. Immunol. 2018, 200, 681–687. [Google Scholar] [CrossRef]
- Buhr, E.D.; Yoo, S.H.; Takahashi, J.S. Temperature as a universal resetting cue for mammalian circadian oscillators. Science 2010, 330, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, Q.J.; Zhang, K.H.; Yao, C.Y.; Huang, J.; Li, Q.; Liu, Z.Y.; Zhang, Y.; Shan, C.H.; Liu, P.; et al. Night-restricted feeding improves locomotor activity rhythm and modulates nutrient utilization to accelerate growth in rabbits. FASEB J. 2021, 35, e21166. [Google Scholar] [CrossRef]
- Feillet, C.A.; Mendoza, J.; Pévet, P.; Challet, E. Restricted feeding restores rhythmicity in the pineal gland of arrhythmic suprachiasmatic-lesioned rats. Eur. J. Neurosci. 2008, 28, 2451–2458. [Google Scholar] [CrossRef] [PubMed]
- Elmowalid, G.A.E.; Ahmad, A.A.M.; El-Hamid, M.I.A.; Ibrahim, D.; Wahdan, A.; El Oksh, A.S.A.; Yonis, A.E.; Elkady, M.A.; Ismail, T.A.; Alkhedaide, A.Q.; et al. Nigella sativa extract potentially inhibited methicillin resistant induced infection in rabbits: Potential immunomodulatory and growth promoting properties. Animals 2022, 12, 2635. [Google Scholar] [CrossRef]
- Wang, Q.; Fu, W.; Guo, Y.; Tang, Y.; Du, H.; Wang, M.; Liu, Z.; Li, Q.; An, L.; Tian, J.; et al. Drinking warm water improves growth performance and optimizes the gut microbiota in early postweaning rabbits during winter. Animals 2019, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Gidenne, T.; Feugier, A. Feed restriction strategy in the growing rabbit. 1. Impact on digestion, rate of passage and microbial activity. Animal 2009, 3, 501–508. [Google Scholar] [CrossRef]
- Fernández-Carmona, J.; Solar, A.; Pascual, J.J.; Blas, E.; Cervera, C. The behaviour of farm rabbit does around parturition and during lactation. World Rabbit Sci. 2010, 13, 253–277. [Google Scholar] [CrossRef]
- Trocino, A.; Filiou, E.; Zomeno, C.; Birolo, M.; Bertotto, D. Behaviour and reactivity of female and male rabbits housed in collective pens: Effects of floor type and stocking density at different ages. World Rabbit Sci. 2018, 26, 135–147. [Google Scholar] [CrossRef]
- Fernández-Carmona, J.; Cervera, C.; Blas, E. Prediction of the energy value of rabbit feeds varying widely in fibre content. Anim. Feed. Sci. Technol. 1996, 64, 61–75. [Google Scholar] [CrossRef]
- Gordon, C.J. Twenty-four hour rhythms of selected ambient temperature in rat and hamster. Physiol. Behav. 1993, 53, 257–263. [Google Scholar] [CrossRef]
- Connolly, M.S.; Lynch, C.B. Circadian variation of strain differences in body temperature and activity in mice. Physiol. Behav. 1981, 27, 1045–1049. [Google Scholar] [CrossRef] [PubMed]
- Caldelas, I.; Gonzalez, B.; Montufar-Chaveznava, R.; Hudson, R. Endogenous clock gene expression in the suprachiasmatic nuclei of previsual newborn rabbits is entrained by nursing. Dev. Neurobiol. 2009, 69, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Van Someren, E.J.; Raymann, R.J.; Scherder, E.J.; Daanen, H.A.; Swaab, D.F. Circadian and age-related modulation of thermoreception and temperature regulation: Mechanisms and functional implications. Ageing Res. Rev. 2002, 1, 721–778. [Google Scholar] [CrossRef]
- Yoda, T.; Crawshaw, L.I.; Yoshida, K.; Su, L.; Hosono, T.; Shido, O.; Sakurada, S.; Fukuda, Y.; Kanosue, K. Effects of food deprivation on daily changes in body temperature and behavioral thermoregulation in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R134–R139. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.; Pirovano, C.; Van Someren, E.J.; Buijs, R.M. Environmental light and suprachiasmatic nucleus interact in the regulation of body temperature. Neuroscience 2005, 132, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Hodge, B.A.; Wen, Y.; Riley, L.A.; Zhang, X.; England, J.H.; Harfmann, B.D.; Schroder, E.A.; Esser, K.A. The endogenous molecular clock orchestrates the temporal separation of substrate metabolism in skeletal muscle. Skelet. Muscle 2015, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Rousso-Noori, L.; Zwighaft, Z.; Neufeld-Cohen, A.; Golik, M.; Kraut-Cohen, J.; Wang, M.; Han, X.; Asher, G. Circadian clocks and feeding time regulate the oscillations and levels of hepatic triglycerides. Cell. Metab. 2014, 19, 319–330. [Google Scholar] [CrossRef]
- Saini, C.; Morf, J.; Stratmann, M.; Gos, P.; Schibler, U. Simulated body temperature rhythms reveal the phase-shifting behavior and plasticity of mammalian circadian oscillators. Genes. Dev. 2012, 26, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.A.; Zumbrunn, G.; Fleury-Olela, F.; Preitner, N.; Schibler, U. Rhythms of mammalian body temperature can sustain peripheral circadian clocks. Curr. Biol. 2002, 12, 1574–1583. [Google Scholar] [CrossRef]
- Mandal, S.; Freije, W.A.; Guptan, P.; Banerjee, U. Metabolic control of G1-S transition: Cyclin E degradation by p53-induced activation of the ubiquitin-proteasome system. J. Cell. Biol. 2010, 188, 473–479. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, Y.; Zhang, K.; Zhang, L.; Geng, S.; Shan, C.; Liu, P.; Zhu, M.; Jin, Q.; Liu, Z.; et al. Night-restricted feeding improves gut health by synchronizing microbe-driven serotonin rhythm and eating activity-driven body temperature oscillations in growing rabbits. Front. Cell. Infect. Microbiol. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Tahara, Y.; Yamazaki, M.; Sukigara, H.; Motohashi, H.; Sasaki, H.; Miyakawa, H.; Haraguchi, A.; Ikeda, Y.; Fukuda, S.; Shibata, S. Gut microbiota-derived short chain fatty acids induce circadian clock entrainment in mouse peripheral tissue. Sci. Rep. 2018, 8, 1395. [Google Scholar] [CrossRef] [PubMed]
- Frazier, K.; Chang, E.B. Intersection of the gut microbiome and circadian rhythms in metabolism. Trends Endocrinol. Metab. 2020, 31, 25–36. [Google Scholar] [CrossRef]
- Bo, T.B.; Zhang, X.Y.; Wen, J.; Deng, K.; Qin, X.W.; Wang, D.H. The microbiota-gut-brain interaction in regulating host metabolic adaptation to cold in male Brandt’s voles (Lasiopodomys brandtii). ISME J. 2019, 13, 3037–3053. [Google Scholar] [CrossRef] [PubMed]
- Tuganbaev, T.; Mor, U.; Bashiardes, S.; Liwinski, T.; Nobs, S.P.; Leshem, A.; Dori-Bachash, M.; Thaiss, C.A.; Pinker, E.Y.; Ratiner, K.; et al. Diet diurnally regulates small intestinal microbiome-epithelial-immune homeostasis and enteritis. Cell 2020, 182, 1441–1459.e21. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, C.; Stojanovic, O.; Colin, D.J.; Suarez-Zamorano, N.; Tarallo, V.; Veyrat-Durebex, C.; Rigo, D.; Fabbiano, S.; Stevanovic, A.; Hagemann, S.; et al. Gut microbiota orchestrates energy homeostasis during cold. Cell 2015, 163, 1360–1374. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Materials | Content | Nutritional Ingredients | Content |
|---|---|---|---|
| Alfalfa meal | 31.80 | Dry matter (g/kg feed) | 868.00 |
| Corn | 27.60 | Crude protein (g/kg DM) | 202.00 |
| Soybean meal | 17.70 | Neutral detergent fibre (g/kg DM) | 318.00 |
| Wheat bran | 20.00 | Ether extract (g/kg DM) | 30.00 |
| Premix | 0.15 | Digestible energy (MJ/kg DM) | 11.89 |
| Salt | 0.40 | Ca (g/kg DM) | 10.40 |
| Limestone | 0.39 | Total phosphorus (g/kg DM) | 7.40 |
| DL-Methionine | 0.41 | Lysine (g/kg DM) | 9.80 |
| Lysine | 0.05 | Methionine + cysteine (g/kg DM) | 9.30 |
| Calcium hydrophosphate | 1.30 | Threonine (g/kg DM) | 9.20 |
| Threonine | 0.2 | ||
| Total | 100.00 |
| Performance Indicators | Age | WRF | NRF | WUF | NUF | p Value | ||
|---|---|---|---|---|---|---|---|---|
| Feed Amount | Feed Time | Feed Amount × Feed Time | ||||||
| Body weight (kg) | 35 | 1.03 ± 0.00 a | 1.02 ± 0.00 a | 1.03 ± 0.00 a | 1.02 ± 0.00 a | 0.837 | 0.057 | 0.898 |
| 42 | 1.30 ± 0.01 b | 1.27 ± 0.01 c | 1.33 ± 0.01 a | 1.25 ± 0.01 d | 0.581 | <0.001 | 0.001 | |
| 49 | 1.64 ± 0.01 a | 1.54 ± 0.01 c | 1.65 ± 0.01 a | 1.59 ± 0.01 b | 0.001 | <0.001 | 0.060 | |
| 56 | 1.95 ± 0.01 a | 1.84 ± 0.01 c | 1.96 ± 0.01 a | 1.90 ± 0.01 b | 0.014 | <0.001 | 0.024 | |
| 63 | 2.14 ± 0.01 b | 2.14 ± 0.01 b | 2.21 ± 0.01 a | 2.18 ± 0.01 a | <0.001 | 0.223 | 0.197 | |
| 70 | 2.43 ± 0.01 bc | 2.42 ± 0.01 c | 2.49 ± 0.01 a | 2.47 ± 0.01 ab | <0.001 | 0.240 | 0.582 | |
| 77 | 2.75 ± 0.01 ab | 2.71 ± 0.01 b | 2.77 ± 0.02 a | 2.74 ± 0.02 ab | 0.077 | 0.016 | 0.827 | |
| 84 | 3.10 ± 0.02 a | 3.03 ± 0.02 b | 3.14 ± 0.02 a | 3.05 ± 0.02 b | 0.219 | <0.001 | 0.567 | |
| Daily gain weight (g) | 36–42 | 38.84 ± 0.74 b | 35.18 ± 0.87 c | 42.39 ± 0.8 a | 32.73 ± 0.85 d | 0.415 | <0.001 | <0.001 |
| 43–49 | 48.54 ± 0.74 a | 37.79 ± 0.86 b | 46.44 ± 0.77 a | 47.47 ± 0.75 a | <0.001 | <0.001 | <0.001 | |
| 50–56 | 44.91 ± 0.81 a | 45.37 ± 0.71 a | 42.26 ± 0.98 b | 46.01 ± 0.77 a | 0.207 | 0.014 | 0.043 | |
| 57–63 | 26.32 ± 0.96 c | 42.81 ± 0.84 a | 36.35 ± 0.86 b | 41.24 ± 0.82 a | <0.001 | <0.001 | <0.001 | |
| 64–70 | 40.62 ± 0.88 ab | 41.63 ± 0.69 ab | 39.41 ± 0.73 b | 42.59 ± 0.76 a | 0.969 | 0.005 | 0.167 | |
| 71–77 | 45.18 ± 1.03 a | 40.82 ± 0.77 bc | 42.67 ± 0.80 b | 38.90 ± 0.90 c | 0.018 | <0.001 | 0.504 | |
| 78–84 | 49.37 ± 0.88 b | 46.45 ± 0.93 c | 52.45 ± 1.16 a | 43.92 ± 1.12 c | 0.652 | <0.001 | 0.018 | |
| 36–84 | 41.66 ± 0.47 b | 41.23 ± 0.29 b | 42.96 ± 0.42 a | 41.26 ± 0.40 b | 0.088 | 0.010 | 0.149 | |
| Daily feed intake (g) | 36–42 | 97.29 ± 0.16 b | 90.79 ± 0.51 c | 113.69 ± 0.79 a | 87.63 ± 0.75 d | <0.001 | <0.001 | <0.001 |
| 43–49 | 114.02 ± 0.38 c | 114.43 ± 0.23 c | 136.9 ± 1.04 a | 127.42 ± 1.02 b | <0.001 | <0.001 | <0.001 | |
| 50–56 | 145.63 ± 0.34 b | 137.66 ± 0.79 c | 151.81 ± 1.17 a | 150.46 ± 0.93 a | < 0.001 | <0.001 | <0.001 | |
| 57–63 | 171.30 ± 0.75 b | 173.248 ± 0.56 b | 176.74 ± 1.53 a | 173.50 ± 1.19 b | 0.785 | 0.002 | <0.001 | |
| 64–70 | 195.60 ± 0.45 a | 191.31 ± 0.78 b | 191.82 ± 1.12 b | 194.74 ± 0.99 a | 0.845 | 0.437 | <0.001 | |
| 71–77 | 205.62 ± 0.61 a | 198.92 ± 1.14 b | 200.22 ± 1.32 b | 193.63 ± 1.29 c | <0.001 | <0.001 | 0.962 | |
| 78–84 | 214.57 ± 0.50 b | 207.28 ± 1.05 c | 221.13 ± 1.61 a | 200.78 ± 1.35 d | 0.983 | <0.001 | <0.001 | |
| 36–84 | 162.24 ± 0.29 b | 157.57 ± 0.49 d | 168.76 ± 0.69 a | 160.07 ± 0.78 c | <0.001 | <0.001 | 0.001 | |
| Feed conversion ratio | 36–42 | 2.61 ± 0.06 a | 2.81 ± 0.09 a | 2.67 ± 0.05 a | 2.77 ± 0.07 a | 0.883 | 0.025 | 0.462 |
| 43–49 | 2.44 ± 0.05 d | 3.08 ± 0.07 a | 2.93 ± 0.05 b | 2.62 ± 0.04 c | 0.785 | 0.002 | <0.001 | |
| 50–56 | 3.33 ± 0.07 b | 3.09 ± 0.08 c | 3.80 ± 0.09 a | 3.34 ± 0.07 b | <0.001 | <0.001 | 0.162 | |
| 57–63 | 6.94 ± 0.26 a | 4.24 ± 0.11 c | 4.95 ± 0.11 b | 4.26 ± 0.09 c | <0.001 | <0.001 | <0.001 | |
| 64–70 | 4.65 ± 0.09 b | 4.68 ± 0.08 b | 5.05 ± 0.10 a | 4.78 ± 0.10 b | 0.007 | 0.193 | 0.101 | |
| 71–77 | 4.63 ± 0.10 b | 4.99 ± 0.10 a | 4.87 ± 0.10 ab | 5.04 ± 0.11 a | 0.160 | 0.011 | 0.353 | |
| 78–84 | 4.30 ± 0.09 b | 4.47 ± 0.10 ab | 4.28 ± 0.11 b | 4.61 ± 0.11 a | 0.506 | 0.004 | 0.354 | |
| 36–84 | 3.87 ± 0.05 b | 3.83 ± 0.02 b | 3.97 ± 0.04 a | 3.86 ± 0.03 b | 0.041 | 0.026 | 0.218 | |
| Behaviors | Time Period (min/24 h) | WRF | NRF | WUF | NUF | p Value | ||
|---|---|---|---|---|---|---|---|---|
| Feed Amount | Feed Time | Feed Amount × Feed Time | ||||||
| Eating | Whole day | 248.06 ± 6.97 a | 184.47 ± 6.61 b | 192.04 ± 6.10 b | 191.27 ± 5.80 b | 0.064 | 0.017 | 0.020 |
| Daytime | 134.88 ± 2.82 aA | 9.21 ± 0.80 cB | 76.86 ± 1.47 bB | 20.96 ± 1.47 cB | <0.001 | <0.001 | <0.001 | |
| Nighttime | 119.26 ± 2.51 bA | 175.26 ± 3.94 aA | 115.19 ± 3.19 bA | 170.31 ± 3.25 aA | 0.962 | <0.001 | 0.629 | |
| Drinking | Whole day | 38.04 ± 2.16 a | 27.84 ± 1.09 a | 32.56 ± 2.27 a | 35.73 ± 3.37 a | 0.801 | 0.463 | 0.167 |
| Daytime | 16.81 ± 0.66 aA | 4.52 ± 0.17 bB | 12.44 ± 0.74 abA | 11.90 ± 1.56 abA | 0.570 | 0.020 | 0.032 | |
| Nighttime | 25.79 ± 1.33 aA | 23.32 ± 0.68 aA | 20.11 ± 1.08 aA | 23.42 ± 1.37 aA | 0.380 | 0.897 | 0.397 | |
| Walking | Whole day | 54.64 ± 2.42 c | 85.66 ± 7.14 b | 131.32 ± 7.8 a | 121.88 ± 5.62 a | <0.001 | 0.631 | 0.002 |
| Daytime | 21.76 ± 1.75 bcA | 28.79 ± 0.48 cB | 46.36 ± 2.25 aB | 47.27 ± 2.09 bB | <0.001 | 0.549 | 0.003 | |
| Nighttime | 33.90 ± 3.47 bA | 45.85 ± 5.15 bA | 84.96 ± 3.49 aA | 75.55 ± 3.11 abA | 0.221 | 0.163 | 0.027 | |
| Grooming | Whole day | 158.33 ± 24.10 a | 175.54 ± 5.11 a | 173.09 ± 3.04 a | 216.12 ± 13.74 a | 0.337 | 0.296 | 0.652 |
| Daytime | 24.12 ± 3.29 cB | 76.48 ± 1.36 bB | 71.80 ± 2.25 bB | 99.44 ± 4.07 aB | 0.002 | 0.001 | 0.760 | |
| Nighttime | 134.21 ± 16.57 aA | 99.06 ± 1.53 aA | 101.29 ± 2.49 aA | 116.28 ± 6.23 aA | 0.505 | 0.407 | 0.459 | |
| Lying | Whole day | 1005.38 ± 9.02 a | 978.11 ± 13.39 ab | 902.13 ± 10.24 b | 904.46 ± 16.54 b | 0.001 | 0.625 | 0.562 |
| Daytime | 520.04 ± 2.97 bA | 607.59 ± 2.66 aA | 512.23 ± 2.02 bA | 553.92 ± 6.71 bA | 0.004 | 0.001 | 0.020 | |
| Nighttime | 485.34 ± 9.23 aB | 370.52 ± 7.03 abB | 389.90 ± 6.00 abB | 350.54 ± 8.23 bB | 0.564 | 0.023 | 0.144 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Wang, Q.; Zhang, K.; He, S.; Liu, Z.; Li, M.; Liu, M.; Guo, Y.; Wu, Z. Optimizing Feeding Strategies for Growing Rabbits: Impact of Timing and Amount on Health and Circadian Rhythms. Animals 2023, 13, 2742. https://doi.org/10.3390/ani13172742
Huang J, Wang Q, Zhang K, He S, Liu Z, Li M, Liu M, Guo Y, Wu Z. Optimizing Feeding Strategies for Growing Rabbits: Impact of Timing and Amount on Health and Circadian Rhythms. Animals. 2023; 13(17):2742. https://doi.org/10.3390/ani13172742
Chicago/Turabian StyleHuang, Jie, Qiangjun Wang, Kehao Zhang, Shuai He, Zhongying Liu, Mingyong Li, Man Liu, Yao Guo, and Zhonghong Wu. 2023. "Optimizing Feeding Strategies for Growing Rabbits: Impact of Timing and Amount on Health and Circadian Rhythms" Animals 13, no. 17: 2742. https://doi.org/10.3390/ani13172742
APA StyleHuang, J., Wang, Q., Zhang, K., He, S., Liu, Z., Li, M., Liu, M., Guo, Y., & Wu, Z. (2023). Optimizing Feeding Strategies for Growing Rabbits: Impact of Timing and Amount on Health and Circadian Rhythms. Animals, 13(17), 2742. https://doi.org/10.3390/ani13172742