

Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of GPR84 in Grass Carp (Ctenopharyngodon Idella)
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents, and Plasmids
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Molecular Cloning of Grass Carp GPR84
2.4. Homology, Phylogenetic, and Chromosome Synteny Analysis of ciGPR84
2.5. Real-Time PCR for Tissue Expression
2.6. Cell Culture
2.7. Functional Characterization of ciGPR84 in Cultured HEK293 Cells
2.8. Statistical Analysis
3. Results
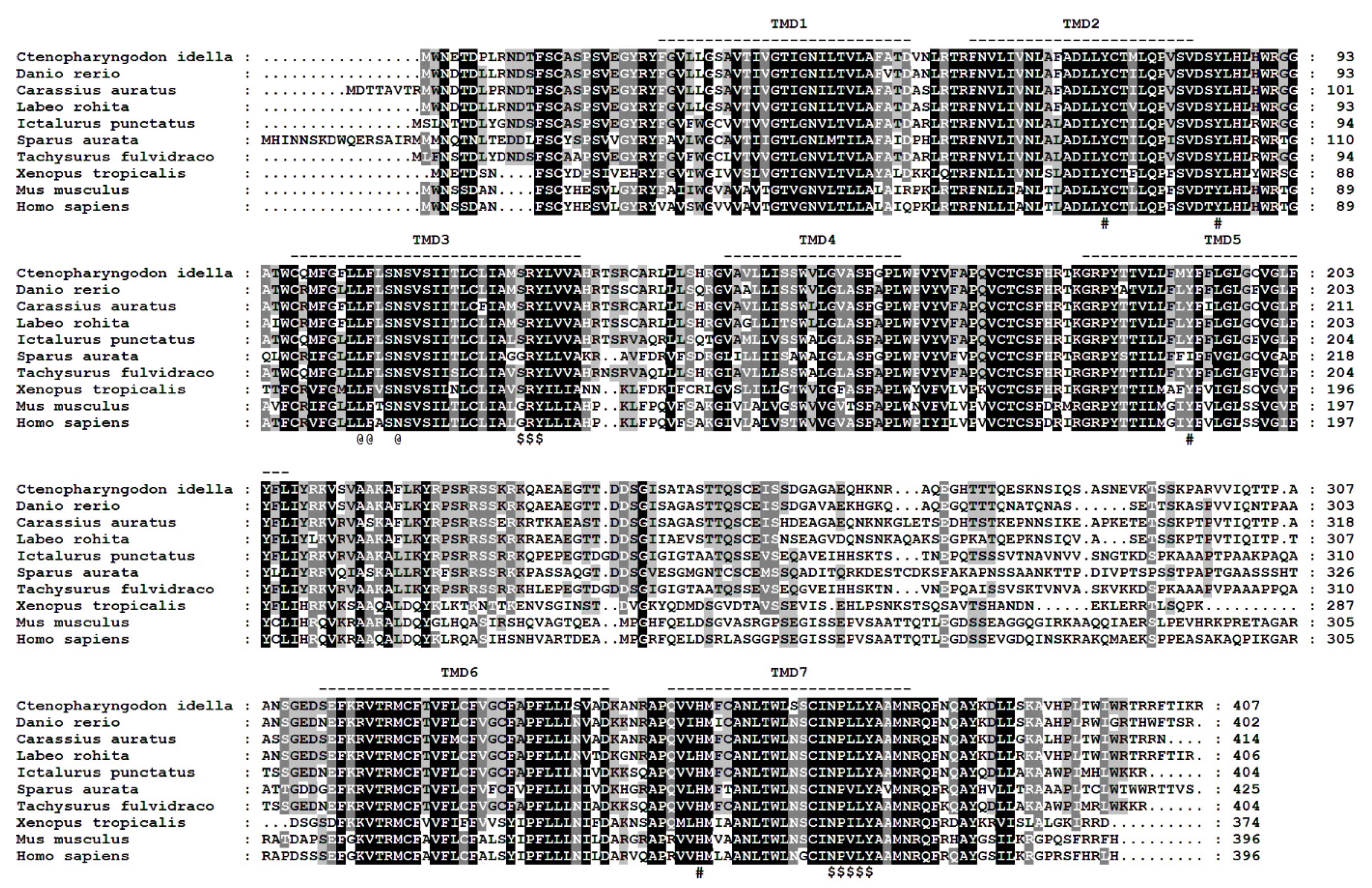
3.1. Analysis of the Full Length of the Sequence
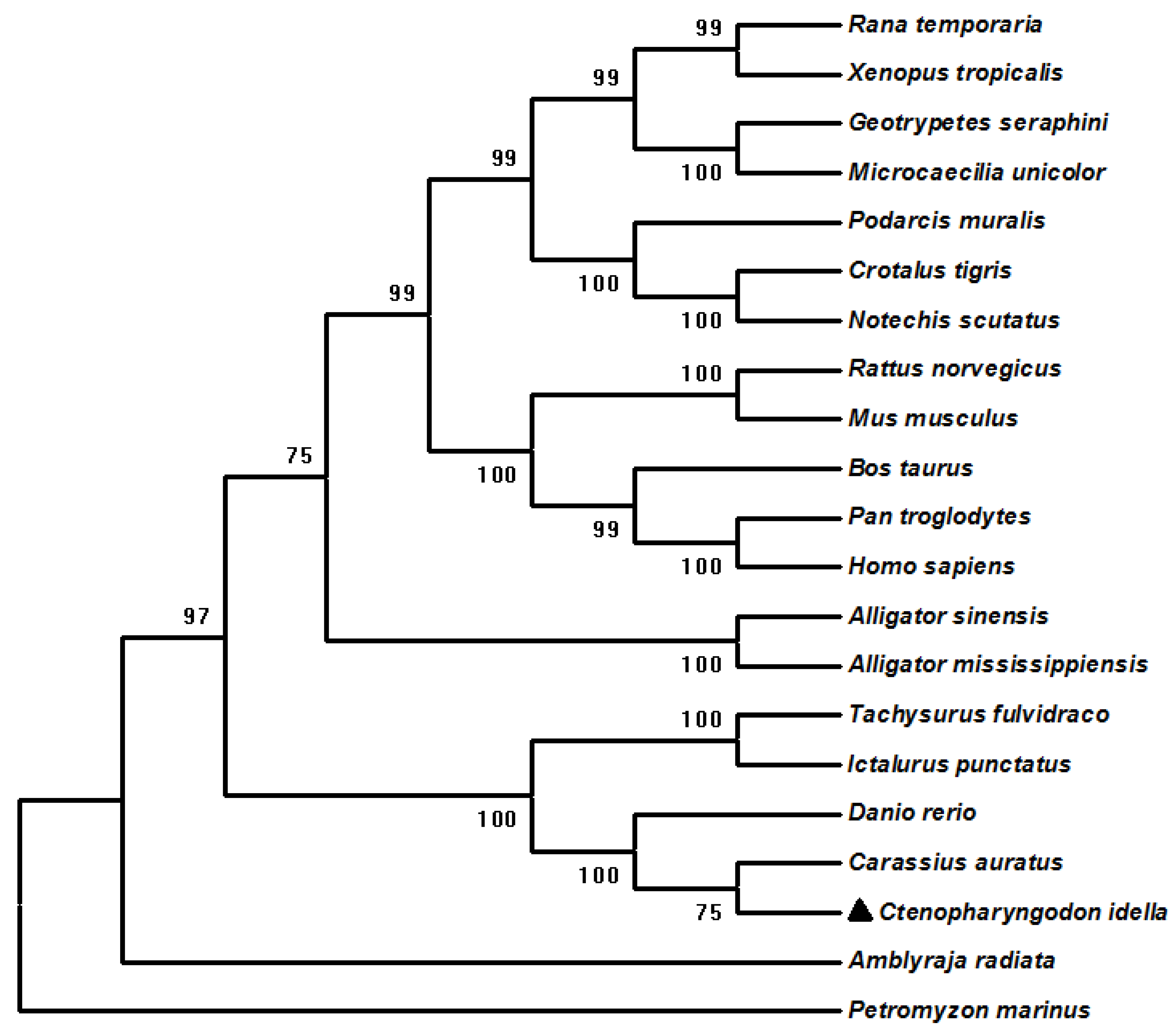
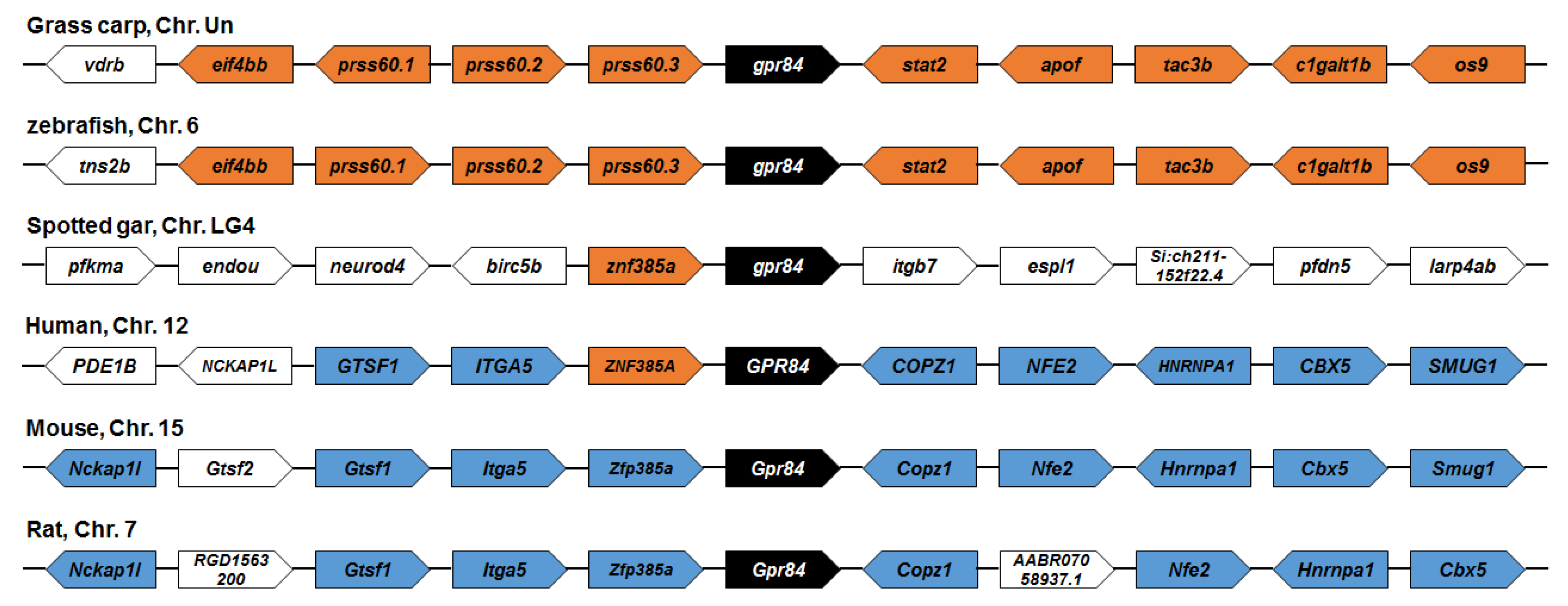
3.2. Phylogenetic and Synteny Analysis of ciGPR84
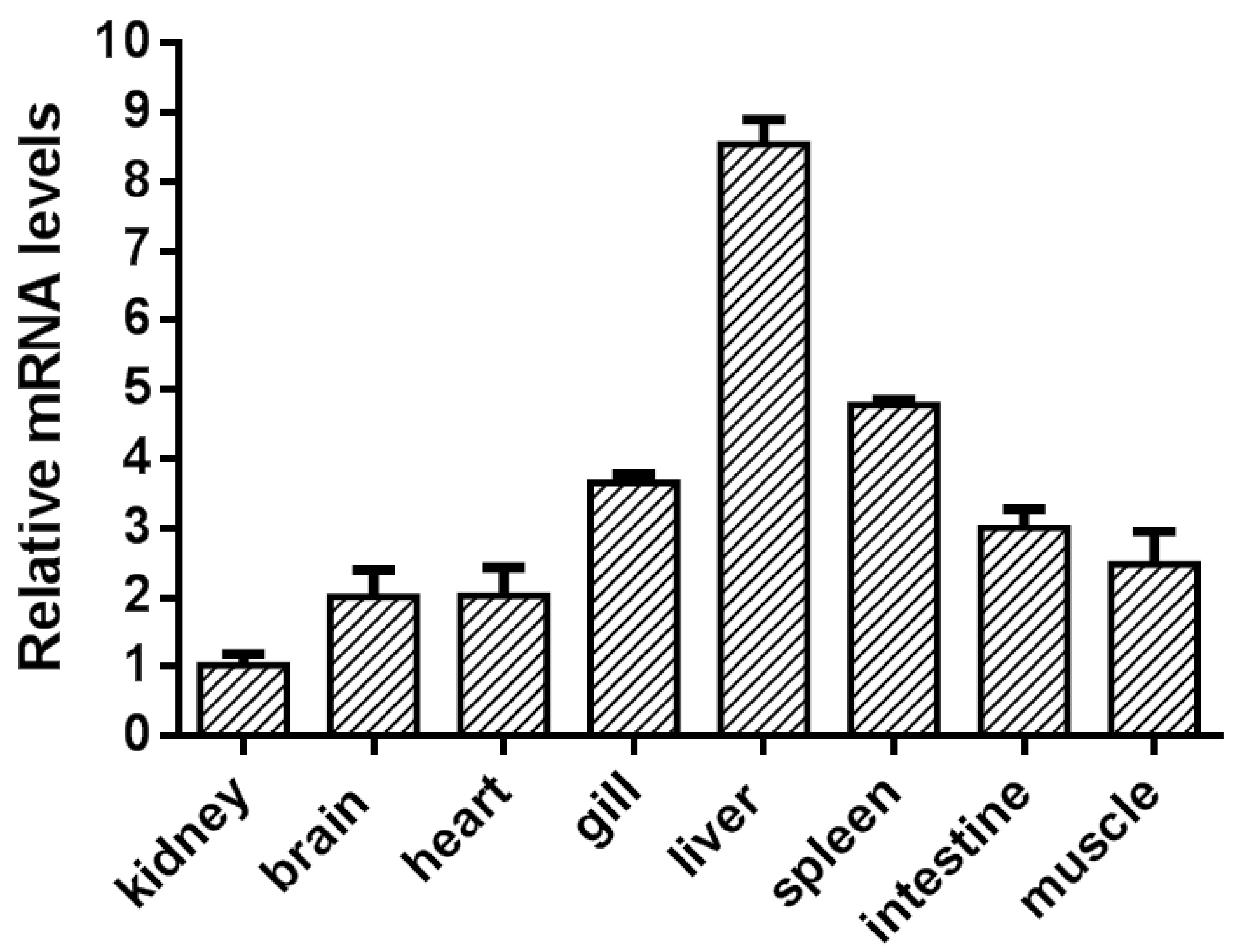
3.3. The Expression of Grass Carp GPR84 in Grass Carp
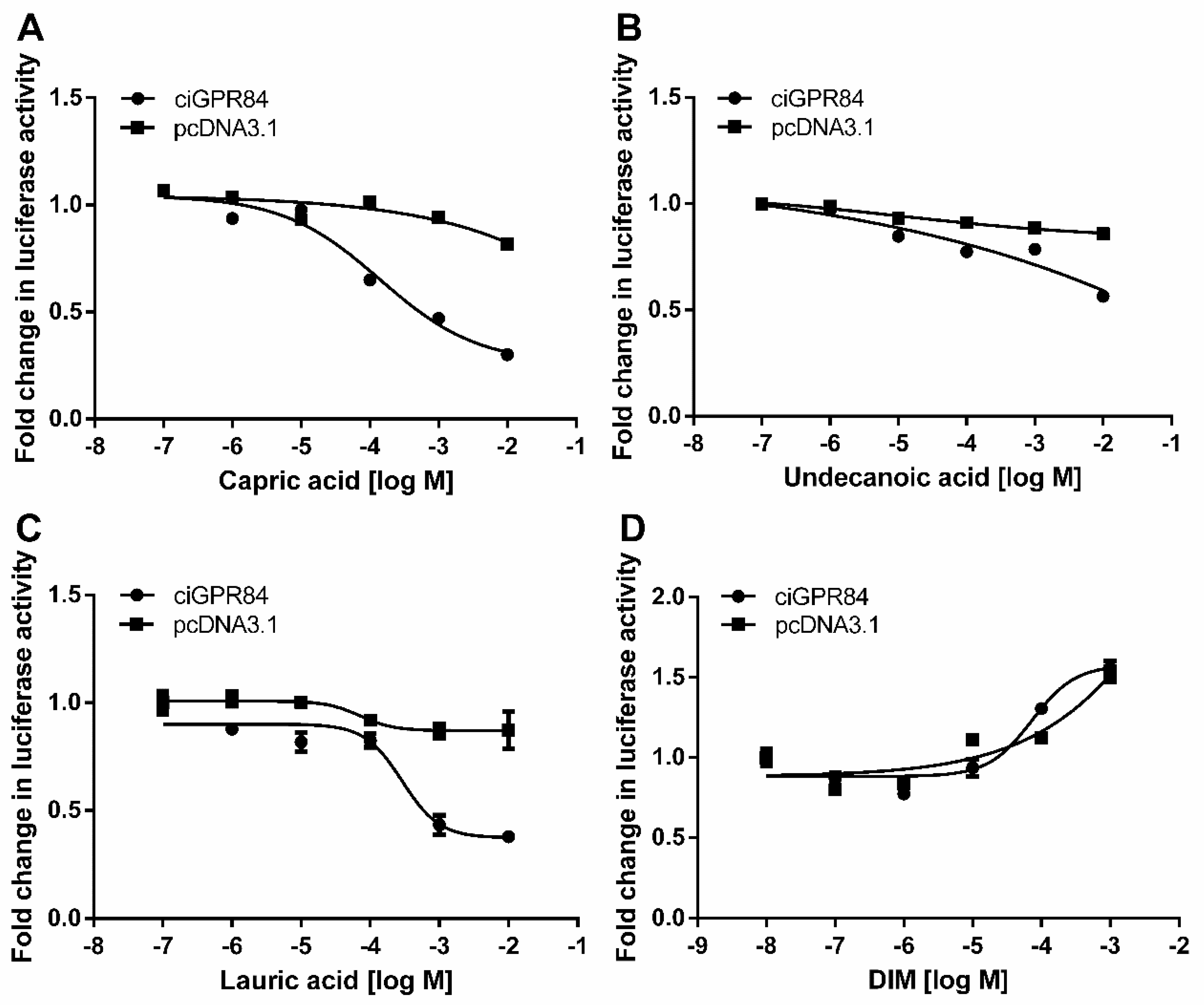
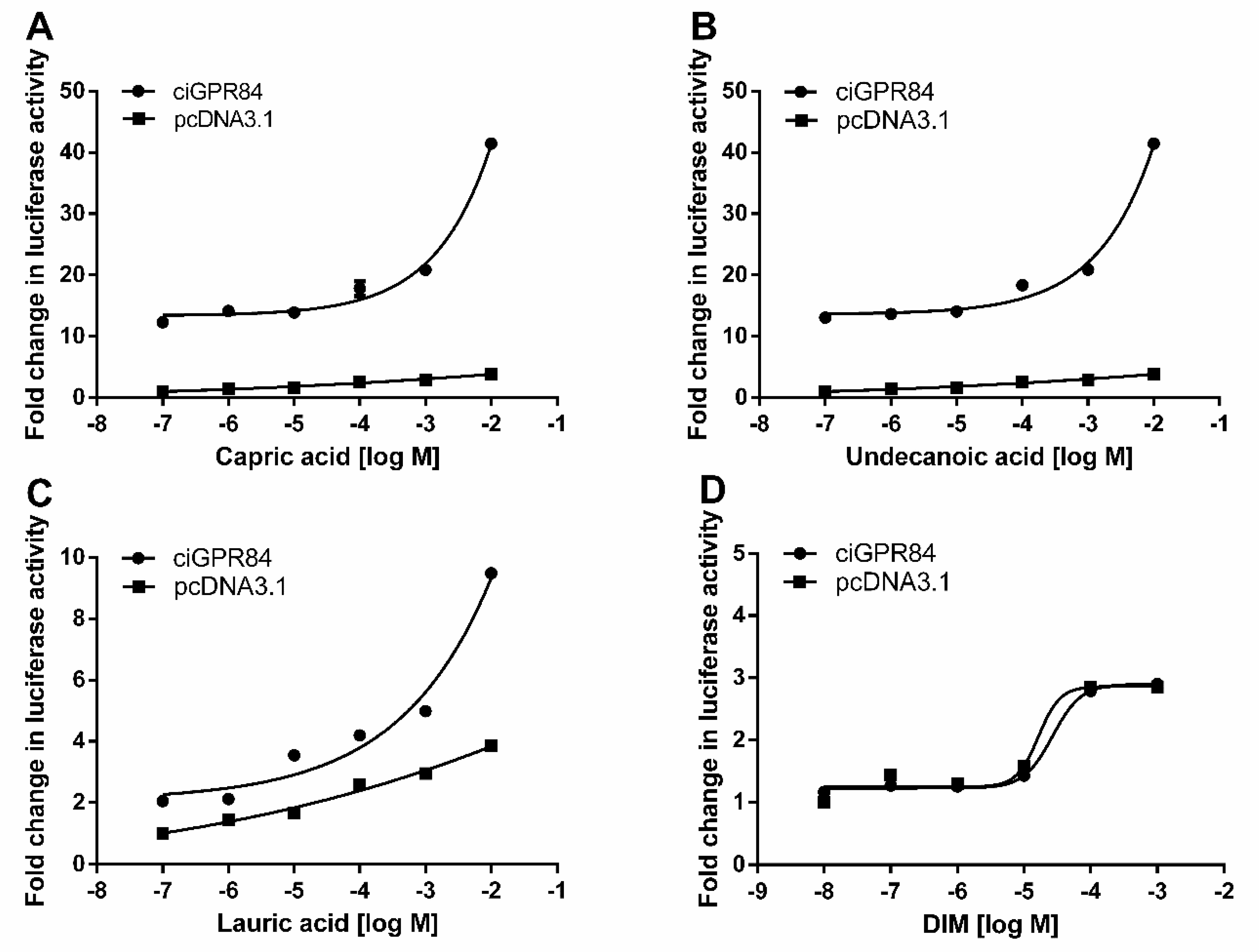
3.4. Functional Characterization of ciGPR84 in Cultured HEK293 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvarez-Curto, E.; Milligan, G. Metabolism Meets Immunity: The Role of Free Fatty Acid Receptors in the Immune System. Biochem. Pharmacol. 2016, 114, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Falomir-Lockhart, L.J.; Cavazzutti, G.F.; Giménez, E.; Toscani, A.M. Fatty Acid Signaling Mechanisms in Neural Cells: Fatty Acid Receptors. Front. Cell. Neurosci. 2019, 13, 162. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L. Gut Microbiota Metabolism of Dietary Fiber Influences Allergic Airway Disease and Hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short-and Medium-Chain Fatty Acids in Energy Metabolism: The Cellular Perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Saresella, M.; Marventano, I.; Barone, M.; La Rosa, F.; Piancone, F.; Mendozzi, L.; D’Arma, A.; Rossi, V.; Pugnetti, L.; Roda, G. Alterations in Circulating Fatty Acid Are Associated with Gut Microbiota Dysbiosis and Inflammation in Multiple Sclerosis. Front. Immunol. 2020, 11, 1390. [Google Scholar] [CrossRef]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.-H.; May, C.; Wilck, N. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The Neuropharmacology of Butyrate: The Bread and Butter of the Microbiota-Gut-Brain Axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Ferguson, G.J.; Kulkarni, S.; Damoulakis, G.; Anderson, K.; Bohlooly-Y, M.; Stephens, L.; Hawkins, P.T.; Curi, R. SCFAs Induce Mouse Neutrophil Chemotaxis through the GPR43 Receptor. PLoS ONE 2011, 6, e21205. [Google Scholar] [CrossRef]
- Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-Inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar]
- Wang, J.; Wu, X.; Simonavicius, N.; Tian, H.; Ling, L. Medium-Chain Fatty Acids as Ligands for Orphan G Protein-Coupled Receptor GPR84. J. Biol. Chem. 2006, 281, 34457–34464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, L.; Yang, H.; Fang, Y.; Wang, S.; Wang, M.; Yuan, Q.; Wu, W.; Zhang, Y.; Liu, Z.; et al. GPR84 Signaling Promotes Intestinal Mucosal Inflammation via Enhancing NLRP3 Inflammasome Activation in Macrophages. Acta Pharmacol. Sin. 2021, 43, 2042–2054. [Google Scholar] [CrossRef] [PubMed]
- Lattin, J.E.; Schroder, K.; Su, A.I.; Walker, J.R.; Zhang, J.; Wiltshire, T.; Saijo, K.; Glass, C.K.; Hume, D.A.; Kellie, S. Expression Analysis of G Protein-Coupled Receptors in Mouse Macrophages. Immunome Res. 2008, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Recio, C.; Lucy, D.; Purvis, G.S.D.; Iveson, P.; Zeboudj, L.; Iqbal, A.J.; Lin, D.; O’Callaghan, C.; Davison, L.; Griesbach, E. Activation of the Immune-Metabolic Receptor GPR84 Enhances Inflammation and Phagocytosis in Macrophages. Front. Immunol. 2018, 9, 1419. [Google Scholar] [CrossRef]
- Venkataraman, C.; Kuo, F. The G-Protein Coupled Receptor, GPR84 Regulates IL-4 Production by T Lymphocytes in Response to CD3 Crosslinking. Immunol. Lett. 2005, 101, 144–153. [Google Scholar] [CrossRef]
- Langmead, C.J.; Summers, R.J. Molecular Pharmacology of GPCRs. Br. J. Pharmacol. 2018, 175, 4005–4008. [Google Scholar] [CrossRef]
- Mafi, A.; Kim, S.-K.; Goddard, W.A., III. The Mechanism for Ligand Activation of the GPCR–G Protein Complex. Proc. Natl. Acad. Sci. USA 2022, 119, e2110085119. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Ringholm, A.; Schiöth, H.B.; Peter, R.E. Molecular Cloning, Pharmacological Characterization, and Brain Mapping of the Melanocortin 4 Receptor in the Goldfish: Involvement in the Control of Food Intake. Endocrinology 2003, 144, 2336–2349. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Z.; Wen, H.; Tao, Y. Characterization of Channel Catfish (Ictalurus punctatus) Melanocortin-3 Receptor Reveals a Potential Network in Regulation of Energy Homeostasis. Gen. Comp. Endocrinol. 2019, 277, 90–103. [Google Scholar] [CrossRef]
- Li, J.T.; Yang, Z.; Chen, H.P.; Zhu, C.H.; Deng, S.P.; Li, G.L.; Tao, Y.X. Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of Melanocortin-4 Receptor in Spotted Scat, Scatophagus argus. Gen. Comp. Endocrinol. 2016, 230–231, 143–152. [Google Scholar] [CrossRef]
- Li, L.; Yang, Z.; Zhang, Y.P.; He, S.; Liang, X.F.; Tao, Y.X. Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of Melanocortin-4 Receptor in Grass Carp (Ctenopharyngodon idella). Domest. Anim. Endocrinol. 2017, 59, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.-Y.; Liu, T.; Qin, C.-J.; Zou, Y.-C.; Wang, J.; Li, R.; Tao, Y.-X. MRAP2 Interaction with Melanocortin-4 Receptor in Snakehead (Channa argus). Biomolecules 2021, 11, 481. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yi, T.-L.; Yang, D.-Q.; Tao, Y.-X. Regulation of Melanocortin-5 Receptor Pharmacology by Two Isoforms of MRAP2 in Ricefield Eel (Monopterus albus). Gen. Comp. Endocrinol. 2021, 314, 113928. [Google Scholar] [CrossRef]
- Tao, M.; Ji, R.L.; Huang, L.; Fan, S.Y.; Liu, T.; Liu, S.J.; Tao, Y.X. Regulation of Melanocortin-4 Receptor Pharmacology by Two Isoforms of Melanocortin Receptor Accessory Protein 2 in Topmouth Culter (Culter alburnus). Front. Endocrinol. 2020, 11, 1489–1501. [Google Scholar] [CrossRef]
- Yu, H.; Li, Y.; Song, W.; Wang, H.; Mo, H.; Liu, Q.; Zhang, X.; Jiang, Z.; Wang, L. Functional Characterization of Melanocortin-3 Receptor in Rainbow Trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2022, 48, 241–252. [Google Scholar] [CrossRef]
- Barney, E.; Dores, M.R.; McAvoy, D.; Davis, P.; Racareanu, R.C.; Iki, A.; Hyodo, S.; Dores, R.M. Elephant Shark Melanocortin Receptors: Novel Interactions with MRAP1 and Implication for the HPI Axis. Gen. Comp. Endocrinol. 2019, 272, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liang, X.-F.; Li, G.-L.; Tao, Y.-X. Biased Signaling in Fish Melanocortin-4 Receptors (MC4Rs): Divergent Pharmacology of Four Ligands on Spotted Scat (Scatophagus argus) and Grass Carp (Ctenopharyngodon idella) MC4Rs. Mol. Cell. Endocrinol. 2020, 515, 110929. [Google Scholar] [CrossRef] [PubMed]
- Al Mahmud, Z.; Jenkins, L.; Ulven, T.; Labéguère, F.; Gosmini, R.; De Vos, S.; Hudson, B.D.; Tikhonova, I.G.; Milligan, G. Three Classes of Ligands Each Bind to Distinct Sites on the Orphan G Protein-Coupled Receptor GPR84. Sci. Rep. 2017, 7, 17953. [Google Scholar] [CrossRef]
- Gaidarov, I.; Anthony, T.; Gatlin, J.; Chen, X.; Mills, D.; Solomon, M.; Han, S.; Semple, G.; Unett, D.J. Embelin and Its Derivatives Unravel the Signaling, Proinflammatory and Antiatherogenic Properties of GPR84 Receptor. Pharmacol. Res. 2018, 131, 185–198. [Google Scholar] [CrossRef]
- Wei, L.; Tokizane, K.; Konishi, H.; Yu, H.-R.; Kiyama, H. Agonists for G-Protein-Coupled Receptor 84 (GPR84) Alter Cellular Morphology and Motility But Do Not Induce Pro-Inflammatory Responses in Microglia. J. Neuroinflamm. 2017, 14, 198. [Google Scholar] [CrossRef]
- Kuwabara, T.; Ishikawa, F.; Kondo, M.; Kakiuchi, T. The Role of IL-17 and Related Cytokines in Inflammatory Autoimmune Diseases. Mediat. Inflamm. 2017, 2017, 3908061. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Feng, D.; Liu, K.; Wang, P.; Xiao, H.; Wang, Y.; Zhang, S.; Liu, Z. A Medium-Chain Fatty Acid Receptor Gpr84 in Zebrafish: Expression Pattern and Roles in Immune Regulation. Dev. Comp. Immunol. 2014, 45, 252–258. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.; Zhang, S.; Liu, Z. Zebrafish Fatty Acids Receptor Gpr84 Enhances Macrophage Phagocytosis. Fish Shellfish Immunol. 2019, 84, 1098–1099. [Google Scholar] [CrossRef] [PubMed]
- Velasco, C.; Conde-Sieira, M.; Comesaña, S.; Chivite, M.; Míguez, J.M.; Soengas, J.L. Role of the G Protein-Coupled Receptors GPR84 and GPR119 in the Central Regulation of Food Intake in Rainbow Trout. J. Exp. Biol. 2021, 224, jeb242360. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.; Su, J. Insights into the Antiviral Immunity against Grass Carp (Ctenopharyngodon idella) Reovirus (GCRV) in Grass Carp. J. Immunol. Res. 2015, 2015, 670437. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shi, M.; Zhang, W.; Cheng, Y.; Wang, Y.; Xia, X.-Q. The Grass Carp Genome Database (GCGD): An Online Platform for Genome Features and Annotations. Database 2017, 2017, bax051. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Davis, P.; Franquemont, S.; Liang, L.; Angleson, J.K.; Dores, R.M. Evolution of the Melanocortin-2 Receptor in Tetrapods: Studies on Xenopus Tropicalis MC2R and Anolis Carolinensis MC2R. Gen. Comp. Endocrinol. 2013, 188, 75–84. [Google Scholar] [CrossRef]
- Yu, H.-X.; Li, Y.; Ezeorba, T.; Mo, H.-L.; Zhang, Z.-H.; Yang, Q.-Y.; Wang, L.-X. Molecular Characterization and Functional Exploration of GPR84 in Chinese Giant Salamander (Andrias davidianus). Dev. Comp. Immunol. 2022, 137, 104526. [Google Scholar] [CrossRef]
- Wu, L.; Yu, H.; Mo, H.; Lan, X.; Pan, C.; Wang, L.; Zhao, H.; Zhou, J.; Li, Y. Functional Characterization of Melanocortin-3 Receptor in a Hibernating Cavefish Onychostoma macrolepis. Animals 2021, 12, 38. [Google Scholar] [CrossRef]
- Lucy, D.; Purvis, G.S.D.; Zeboudj, L.; Chatzopoulou, M.; Recio, C.; Bataille, C.J.R.; Wynne, G.M.; Greaves, D.R.; Russell, A.J. A Biased Agonist at Immunometabolic Receptor GPR84 Causes Distinct Functional Effects in Macrophages. ACS Chem. Biol. 2019, 14, 2055–2064. [Google Scholar] [CrossRef]
- Takeda, S.; Yamamoto, A.; Okada, T.; Matsumura, E.; Nose, E.; Kogure, K.; Kojima, S.; Haga, T. Identification of Surrogate Ligands for Orphan G Protein-Coupled Receptors. Life Sci. 2003, 74, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.-L.; Yang, L.-K.; Ruan, G.-L.; Yang, D.-Q.; Tao, Y.-X. Melanocortin-4 Receptor in Swamp Eel (Monopterus albus): Cloning, Tissue Distribution, and Pharmacology. Gene 2018, 678, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.-Q.; Hou, Z.-S.; Wen, H.-S.; Li, Y.; Qi, X.; Li, W.-J.; Tao, Y.-X. Melanocortin-4 Receptor in Spotted Sea Bass, Lateolabrax Maculatus: Cloning, Tissue Distribution, Physiology, and Pharmacology. Front. Endocrinol. 2019, 10, 705. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Product Size (bp) | Ta 1 (°C) |
|---|---|---|---|
| ciGPR84-F | ATGTGGAACGAGACGGA | 1224 | 53 |
| ciGPR84-R | AGCTTTAGGTCATATTCGGA | ||
| qPCR-F | CTGATAGTAAATCTTGCCTTCGC | 194 | 60 |
| qPCR-R | ACAACCAGGTAACGGCTCATAG | ||
| ciβ-actin F | CGTGACATCAAGGAGAAG | 215 | 57 |
| ciβ-actin R | GAGTTGAAGGTGGTCTCAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Song, W.-J.; Yi, S.-K.; Yu, H.-X.; Mo, H.-L.; Yao, M.-X.; Tao, Y.-X.; Wang, L.-X. Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of GPR84 in Grass Carp (Ctenopharyngodon Idella). Animals 2023, 13, 3001. https://doi.org/10.3390/ani13193001
Li Y, Song W-J, Yi S-K, Yu H-X, Mo H-L, Yao M-X, Tao Y-X, Wang L-X. Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of GPR84 in Grass Carp (Ctenopharyngodon Idella). Animals. 2023; 13(19):3001. https://doi.org/10.3390/ani13193001
Chicago/Turabian StyleLi, Yang, Wei-Jia Song, Shao-Kui Yi, Hui-Xia Yu, Hao-Lin Mo, Ming-Xing Yao, Ya-Xiong Tao, and Li-Xin Wang. 2023. "Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of GPR84 in Grass Carp (Ctenopharyngodon Idella)" Animals 13, no. 19: 3001. https://doi.org/10.3390/ani13193001
APA StyleLi, Y., Song, W.-J., Yi, S.-K., Yu, H.-X., Mo, H.-L., Yao, M.-X., Tao, Y.-X., & Wang, L.-X. (2023). Molecular Cloning, Tissue Distribution, and Pharmacological Characterization of GPR84 in Grass Carp (Ctenopharyngodon Idella). Animals, 13(19), 3001. https://doi.org/10.3390/ani13193001