


Tend and Befriend in Horses: Partner Preferences, Lateralization, and Contextualization of Allogrooming in Two Socially Stable Herds of Quarter Horse Mares

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Management
2.2. Videos Collection
2.3. Statistical Analysis
3. Results
3.1. Allogrooming Measurements in Confined Settings
3.2. Allogrooming Measurements in the Pasture Settings
3.3. Comparison between the Two Settings in Allogrooming Measurements
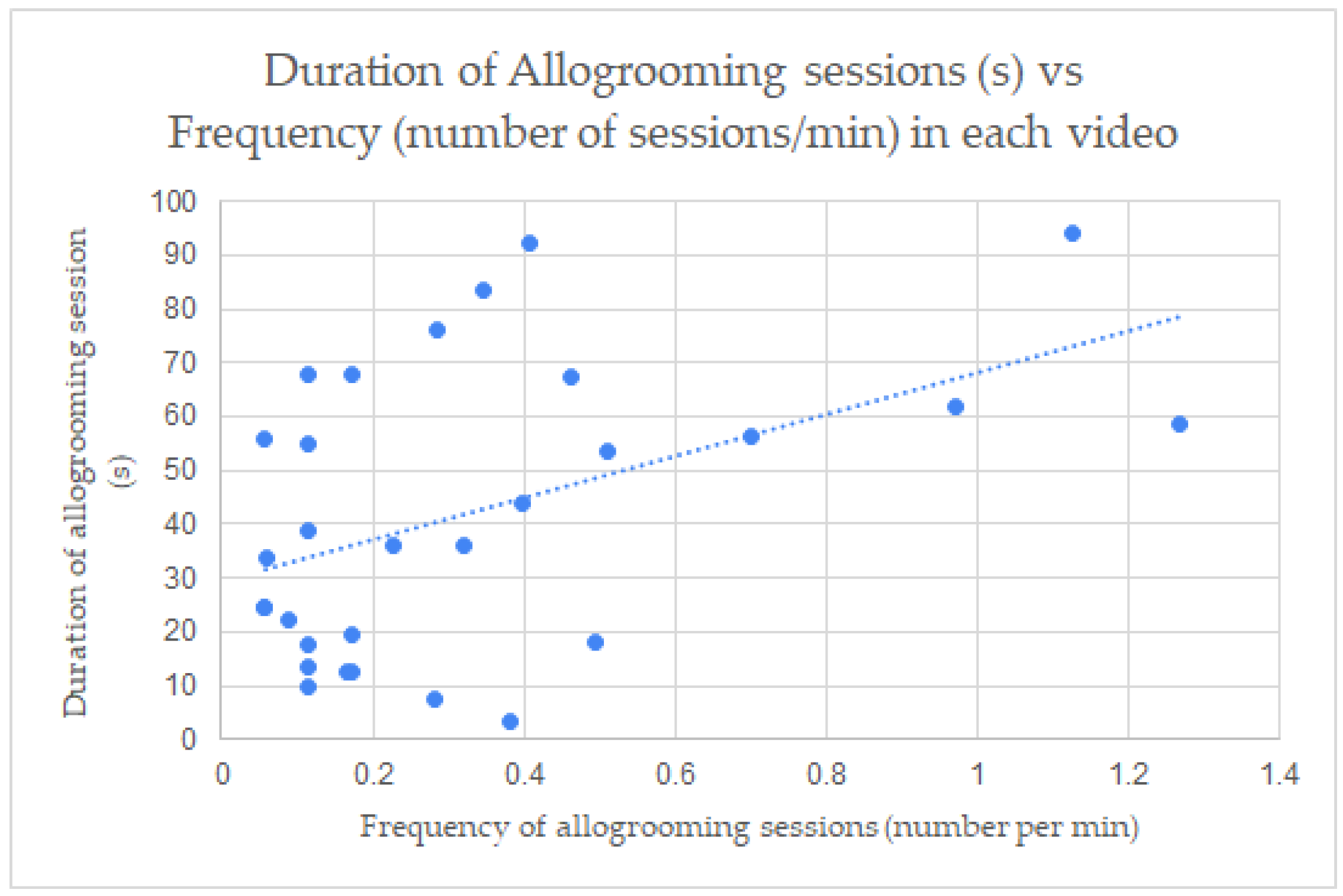
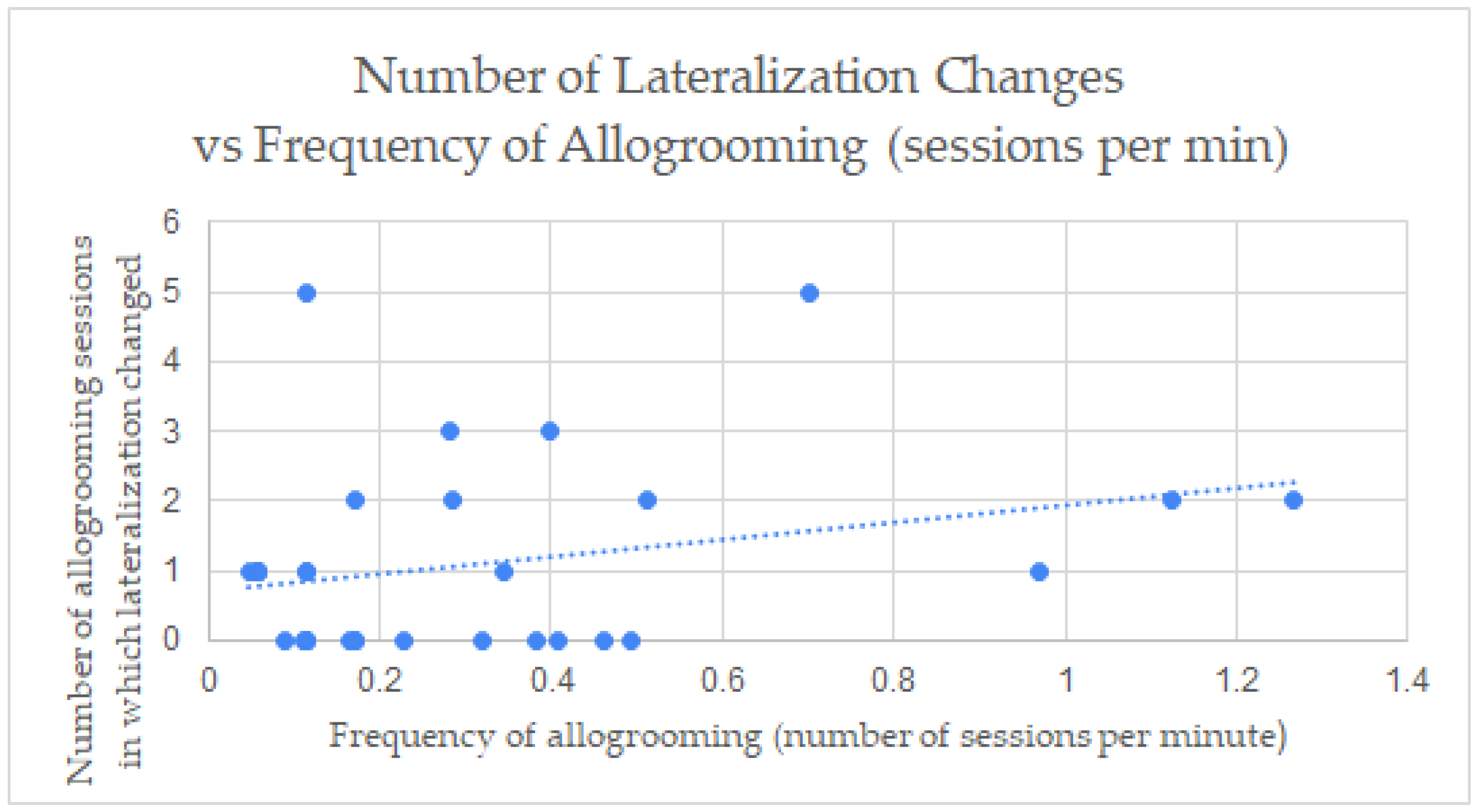
3.4. Correlation between the Allogrooming Measurements
4. Discussion
4.1. Partner Preferences
4.2. Number of Pairs, Frequency, and Duration
4.3. Lateralization
4.4. Welfare
4.5. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Massen, J.J.M.; Sterck, E.H.M.; De Vos, H. Close social associations in animals and humans: Functions and mechanisms of friendship. Behaviour 2010, 147, 1379–1412. [Google Scholar] [CrossRef]
- Wasilewski, A. “ Friendship “ in Ungulates?-Socio-Positive Relationships between Non-Related Herd Members of the Same Species. Ph.D. Thesis, Fachbereich Biologie, Philipps University of Marburg, Marburg, Germany, 2003. [Google Scholar]
- Ramírez-i-Ollé, M. Friendship as a scientific method. Sociol. Rev. 2019, 67, 299–317. [Google Scholar] [CrossRef]
- Wispé, L.G. Positive forms of social behavior: An overview. J. Soc. Issues 1972, 28, 1–19. [Google Scholar] [CrossRef]
- Rault, J.L. Be kind to others: Prosocial behaviours and their implications for animal welfare. Appl. Anim. Behav. Sci. 2019, 210, 113–123. [Google Scholar] [CrossRef]
- Raspa, F.; Tarantola, M.; Muca, E.; Bergero, D.; Soglia, D.; Cavallini, D.; Vervuert, I.; Bordin, C.; De Palo, P.; Valle, E. Does Feeding Management Make a Difference to Behavioural Activities and Welfare of Horses Reared for Meat Production? Animals 2022, 12, 1740. [Google Scholar] [CrossRef]
- Bartlett, E.; Cameron, L.J.; Freeman, M.S. A preliminary comparison between proximity and interaction-based methods to construct equine (Equus caballus) social networks. J. Vet. Behav. 2022, 50, 36–45. [Google Scholar] [CrossRef]
- Schneider, G.; Krueger, K. Third-party interventions keep social partners from exchanging affiliative interactionswith others. Anim. Behav. 2012, 83, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Stanley, C.R.; Mettke-Hofmann, C.; Hager, R.; Shultz, S. Social stability in semiferal ponies: Networks show interannual stability alongside seasonal flexibility. Anim. Behav. 2018, 136, 175–184. [Google Scholar] [CrossRef] [Green Version]
- VanDierendonck, M.C.; de Vries, H.; Schilder, M.B.H.; Colenbrander, B.; Thorhallsdóttir, A.G.; Sigurjónsdóttir, H. Interventions in social behaviour in a herd of mares and geldings. Appl. Anim. Behav. Sci. 2009, 116, 67–73. [Google Scholar] [CrossRef]
- Maeda, T.; Ochi, S.; Ringhofer, M.; Sosa, S.; Sueur, C.; Hirata, S.; Yamamoto, S. Aerial drone observations identified a multilevel society in feral horses. Sci. Rep. 2021, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Bouskila, A.; Lourie, E.; Sommer, S.; De Vries, H.; Hermans, Z.M.; Van Dierendonck, M. Similarity in sex and reproductive state, but not relatedness, influence the strength of association in the social network of feral horses in the Blauwe Kamer Nature Reserve. Isr. J. Ecol. Evol. 2015, 61, 106–113. [Google Scholar] [CrossRef]
- Heitor, F.; do Mar Oom, M.; Vicente, L. Social relationships in a herd of Sorraia horses. Part II. Factors affecting affiliative relationships and sexual behaviours. Behav. Process. 2006, 73, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Kieson, E.; Sams, J. A Preliminary Investigation of Preferred Affiliative Interactions within and between Select Bonded Pairs of Horses: A First Look at Equine “ Love Languages ”. Int. J. Zool. Anim. Biol. 2021, 4, 000318. [Google Scholar] [CrossRef]
- Wolter, R.; Stefanski, V.; Krueger, K. Parameters for the analysis of social bonds in horses. Animals 2018, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, J.W.; Ladewig, J.; Søndergaard, E.; Malmkvist, J. Effects of individual versus group stabling on social behaviour in domestic stallions. Appl. Anim. Behav. Sci. 2002, 75, 233–248. [Google Scholar] [CrossRef]
- Pierard, M.; McGreevy, P.; Geers, R. Effect of density and relative aggressiveness on agonistic and affiliative interactions in a newly formed group of horses. J. Vet. Behav. 2019, 29, 61–69. [Google Scholar] [CrossRef]
- VanDierendonck, M.C.; Spruijt, B.M. Coping in groups of domestic horses-Review from a social and neurobiological perspective. Appl. Anim. Behav. Sci. 2012, 138, 194–202. [Google Scholar] [CrossRef]
- Pierard, M.; McGreevy, P.; Geers, R. Reliability of a descriptive reference ethogram for equitation science. J. Vet. Behav. 2019, 29, 118–127. [Google Scholar] [CrossRef]
- Feh, C.; de Mazierès, J. Grooming at a preferred site reduces heart rate in horses. Anim. Behav. 1993, 46, 1191–1194. [Google Scholar] [CrossRef] [Green Version]
- Kimura, R. Mutual grooming and preferred associate relationships in a band of free-ranging horses. Appl. Anim. Behav. Sci. 1998, 59, 265–276. [Google Scholar] [CrossRef]
- Van Dierendonck, M.C.; Sigurjónsdóttir, H.; Colenbrander, B.; Thorhallsdóttir, A.G. Differences in social behaviour between late pregnant, post-partum and barren mares in a herd of Icelandic horses. Appl. Anim. Behav. Sci. 2004, 89, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Shimada, M.; Suzuki, N. The contribution of mutual grooming to affiliative relationships in a feral misaki horse herd. Animals 2020, 10, 1564. [Google Scholar] [CrossRef]
- Baragli, P.; Scopa, C.; Felici, M.; Reddon, A.R. Horses show individual level lateralisation when inspecting an unfamiliar and unexpected stimulus. PLoS ONE 2021, 16, e0255688. [Google Scholar] [CrossRef]
- Bloom, J.S.; Hynd, G.W. The role of the corpus callosum in interhemispheric transfer of information: Excitation or inhibition? Neuropsychol. Rev. 2005, 15, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Van der Knaap, L.J.; Van der Ham, I.J.M. How does the corpus callosum mediate interhemispheric transfer? A review. Behav. Brain Res. 2011, 223, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Letzner, S.; Simon, A.; Güntürkün, O. Connectivity and neurochemistry of the commissura anterior of the pigeon (Columba livia). J. Comp. Neurol. 2016, 524, 343–361. [Google Scholar] [CrossRef]
- Farmer, K.; Krüger, K.; Byrne, R.W.; Marr, I. Sensory laterality in affiliative interactions in domestic horses and ponies (Equus caballus). Anim. Cogn. 2018, 21, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Crosby, A. A study of lateralized behaviours in domestic horses (Equus caballus). Master’s Thesis, Linköping University, Department of Physics, Chemistry and Biology, Linköping, Sweden, 2021. [Google Scholar]
- Harrison, K.E.; Byrne, R.W. Hand preferences in unimanual and bimanual feeding by wild vervet monkeys (Cercopithecus aethiops). J. Comp. Psychol. 2000, 114, 13–21. [Google Scholar] [CrossRef]
- Rogers, L.J. Brain Lateralization and Cognitive Capacity. Animals 2021, 11, 1996. [Google Scholar] [CrossRef]
- Berretz, G.; Packheiser, J.; Wolf, O.T.; Ocklenburg, S. Dichotic listening performance and interhemispheric integration after stress exposure. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Austin, N.P.; Rogers, L.J. Asymmetry of flight and escape turning responses in horses. Laterality 2007, 12, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Austin, N.P.; Rogers, L.J. Limb preferences and lateralization of aggression, reactivity and vigilance in feral horses, Equus caballus. Anim. Behav. 2012, 83, 239–247. [Google Scholar] [CrossRef]
- Torcivia, C.; McDonnell, S. Equine discomfort ethogram. Animals 2021, 11, 580. [Google Scholar] [CrossRef]
- Luz, M.P.F.; Maia, C.M.; Pantoja, J.C.F.; Neto, M.C.; Puoli Filho, J.N.P. Feeding Time and Agonistic Behavior in Horses: Influence of Distance, Proportion, and Height of Troughs. J. Equine Vet. Sci. 2015, 35, 843–848.e1. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.E. Taylor Tend and Befriend-Biobehavioral Bases of Affiliation Under Stress.pdf. Curr. Dir. Psychol. Sci. 2006, 15, 273–277. [Google Scholar] [CrossRef]
- Taylor, S.E.; Master, S.L. Social responses to stress: The tend-and-befriend model. Handb. Stress Sci. Biol. Psychol. Health 2011, 101, 109. [Google Scholar]
- Decety, J.; Bartal, I.B.A.; Uzefovsky, F.; Knafo-Noam, A. Empathy as a driver of prosocial behaviour: Highly conserved neurobehavioural mechanisms across species. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondrakiewicz, K.; Kostecki, M.; Szadzińska, W.; Knapska, E. Ecological validity of social interaction tests in rats and mice. Genes Brain Behav. 2019, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Scheggia, D.; Papaleo, F. Social Neuroscience: Rats Can Be Considerate to Others. Curr. Biol. 2020, 30, R274–R276. [Google Scholar] [CrossRef]
- Steinbeis, N.; Engert, V.; Linz, R.; Singer, T. The effects of stress and affiliation on social decision-making: Investigating the tend-and-befriend pattern. Psychoneuroendocrinology 2015, 62, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.; Valkanas, H.; Serravalle, L.; Ellenbogen, M.A. Oxytocin and social context moderate social support seeking in women during negative memory recall. Psychoneuroendocrinology 2016, 70, 63–69. [Google Scholar] [CrossRef]
- Hartmann, E.; Søndergaard, E.; Keeling, L.J. Keeping horses in groups: A review. Appl. Anim. Behav. Sci. 2012, 136, 77–87. [Google Scholar] [CrossRef]
- Rose-Meierhöfer, S.; Klaer, S.; Ammon, C.; Brunsch, R.; Hoffmann, G. Activity behavior of horses housed in different open barn systems. J. Equine Vet. Sci. 2010, 30, 624–634. [Google Scholar] [CrossRef]
- Topczewska, J. An attempt to assess the welfare of horses maintained in herd systems. Arch. Anim. Breed. 2014, 57, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Majecka, K.; Klawe, A. Influence of Paddock Size on Social Relationships in Domestic Horses. J. Appl. Anim. Welf. Sci. 2018, 21, 8–16. [Google Scholar] [CrossRef]
- Christensen, J.W.; Søndergaard, E.; Thodberg, K.; Halekoh, U. Effects of repeated regrouping on horse behaviour and injuries. Appl. Anim. Behav. Sci. 2011, 133, 199–206. [Google Scholar] [CrossRef]
- Heitor, F.; Oom, M.; do, M.; Vicente, L. Social relationships in a herd of Sorraia horses. Part I. Correlates of social dominance and contexts of aggression. Behav. Process. 2006, 73, 170–177. [Google Scholar] [CrossRef]
- Heitor, F.; Vicente, L. Dominance relationships and patterns of aggression in a bachelor group of Sorraia horses (Equus caballus). J. Ethol. 2010, 28, 35–44. [Google Scholar] [CrossRef]
- Basile, M.; Boivin, S.; Boutin, A.; Blois-Heulin, C.; Hausberger, M.; Lemasson, A. Socially dependent auditory laterality in domestic horses (Equus caballus). Anim. Cogn. 2009, 12, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.L.; Vallortigara, G. When and why did brains break symmetry? Symmetry 2015, 7, 2181–2194. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.J. A matter of degree: Strength of brain asymmetry and behaviour. Symmetry 2017, 9, 57. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Cantalupo, C. Theoretical speculations on the evolutionary origins of hemispheric specialization. Curr. Dir. Psychol. Sci. 2008, 17, 233–237. [Google Scholar] [CrossRef]
- Dharmaretnam, M.; Rogers, L.J. Hemispheric specialization and dual processing in strongly versus weakly lateralized chicks. Behav. Brain Res. 2005, 162, 62–70. [Google Scholar] [CrossRef]
- Lippolis, G.; Bizazza, A.; Rogers, R.J.; Vallortigara, G. Lateralization of predator avoidance responses in three species of toads. Laterality 2002, 7, 163–183. [Google Scholar] [CrossRef] [PubMed]
- De Boyer Des Roches, A.; Richard-Yris, M.A.; Henry, S.; Ezzaouia, M.; Hausberger, M. Laterality and emotions: Visual laterality in the domestic horse (Equus caballus) differs with objects’ emotional value. Physiol. Behav. 2008, 94, 487–490. [Google Scholar] [CrossRef]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, I.; Wiśniewska, M.; Wilk, I.; Liss, M.; Wnuk-Pawlak, E.; Dybczyńska, M. Assessment of affiliative behaviour in mares. Rocz. Nauk. Pol. Tow. Zootech. 2019, 15, 49–58. [Google Scholar] [CrossRef]
- Mellor, D.J.; Burns, M. Using the Five Domains Model to develop welfare assessment guidelines for Thoroughbred horses in New Zealand. N. Z. Vet. J. 2020, 68, 150–156. [Google Scholar] [CrossRef]
- Placci, M.; Marliani, G.; Sabioni, S.; Gabai, G.; Mondo, E.; Borghetti, P.; De Angelis, E.; Accorsi, P.A. Natural horse boarding vs. traditional stable: A comparison of hormonal, hematological and immunological parameters. J. Appl. Anim. Welf. Sci. 2020, 23, 366–377. [Google Scholar] [CrossRef]
- Mellor, D.J. Enhancing animal welfare by creating opportunities for positive affective engagement. N. Z. Vet. J. 2015, 63, 3–8. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Videos | Number | Length |
|---|---|---|
| Total videos | 185 | 33 min/video |
| No horse and/or undesignated herds | 32 | Excluded from Study |
| Total videos for herd 1 and 2: confined + pasture settings | 153 | Total: 4782.11 min |
| Combined videos for Herd 1 and 2: confined setting | 33 | Total: 411.64 min, Mean: 12.47 min |
| Combined videos for Herd 1 and 2: pasture setting | 120 | Total: 1915.08 min, Mean: 15.96 min |
| Total videos for Herd 1: confined and pasture settings | 94 | Total: 1553.54 min, Mean: 14.66 min |
| Total videos for Herd 2: confined and pasture settings | 59 | Total: 901.85 min, Mean: 15.29 min |
| Total videos for Herd 1: confined setting | 21 | Total: 282.97 min, Mean: 13.47 min |
| Total videos for Herd 1: pasture setting | 73 | Total: 1141.90 min, Mean: 15.64 min |
| Total videos for Herd 2: confined setting | 12 | Total: 128.67 min, Mean: 10.72 min |
| Total videos for Herd 2: pasture setting | 47 | Total: 773.18 min, Mean: 16.45 min |
| Total number of videos in which allogrooming appeared | 33 | Total: 462.13 min |
| Total number of of videos in which allogrooming appeared for Herd 1 and 2: confined setting | 29 | Total: 388.55 min, Mean: 13.03 min |
| Total number of of videos in which allogrooming appeared for Herd 1 and 2: pasture setting | 4 | Total: 73.58 min, Mean: 18.39 min |
| Total number of of videos in which allogrooming appeared for Herd 1: confined setting | 18 | Total: 262.29 min, Mean: 14.57 min |
| Total number of of videos in which allogrooming appeared for Herd 1: pasture setting | 4 | Total: 73.58 min, Mean 18.39 min |
| Total number of of videos in which allogrooming appeared for Herd 2: confined setting | 11 | Total: 126.30 min, Mean 11.48 min |
| Total number of of videos in which allogrooming appeared for Herd 2: pasture setting | 0 | 0 |
| Item | Total | Confined Environment (Higher Stress) | Pasture Environment (Low Stress) |
|---|---|---|---|
| Total number of allogrooming sessions | 124 | 118 | 6 |
| Average Frequency of allogrooming sessions (per minute of video) | 0.124 | 0.563 | 0.003 |
| Number of affiliative pairs allogrooming | 124 | 118 | 6 |
| Average duration of individual allogrooming sessions | 55.784 | 40.98s | 163.11s |
| Number of partners in affiliative interactions | 1 | 1 | 1 |
| Number of allogrooming sessions with single partner | 123 | 117 | 6 |
| Number of horses with two or more partners | 1 | 1 (One horse with three partners in one video) | 0 |
| Total Number of pairs engaged in left allogrooming | 69 * | 66 | 3 |
| Total number of pairs engaged in right allogrooming | 74 * | 71 | 3 |
| Confined | Pasture | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Item | Total | Mean (M) | SD | df | Mean (M) | SD | df | T-Value | p-Value |
| Duration (s) of allogrooming sessions | 1840.86 | 40.980 | 26.762 | 28 | 163.108 | 150.737 | 3 | 2.744 | <0.001 ** |
| Duration (s) of allogrooming sessions in Herd 1 | 1436.11 | 43.538 | 24.938 | 17 | 163.108 | 150.737 | 3 | 2.845 | 0.003 * |
| Duration (s) of allogrooming sessions in Herd 2 | 404.75 | 36.795 | 30.284 | 11 | N/A | N/A | N/A | N/A | N/A |
| Total number of pairs allogrooming (single partners) | 124 | 3.576 | 4.486 | 32 | 0.050 | 0.219 | 119 | 2.609 | <0.001 ** |
| Number of pairs allogrooming in Herd 1 (single partner) | 93 | 4.143 | 5.369 | 20 | 0.050 | 0.219 | 72 | 2.630 | <0.001 ** |
| Number of pairs allogrooming in Herd 2 (single partner) | 33 | 2.583 | 2.109 | 12 | 0.000 | 0.000 | 47 | 2.665 | <0.001 ** |
| Confined | Pasture | |||||||
|---|---|---|---|---|---|---|---|---|
| Item | Mean (M) | SD | df | Mean (M) | SD | df | T-Value | p-Value |
| Herd 1 | 0.716 | 1.905 | 21 | 0.005 | 0.020 | 72 | 2.630 | 0.002 * |
| Herd 2 | 0.295 | 0.255 | 12 | 0.000 | 0.000 | 46 | 2.665 | <0.0001 ** |
| Combined (Herd 1 and Herd 2) | 0.563 | 1.528 | 32 | 0.003 | 0.016 | 119 | 2.609 | <0.0001 ** |
| Herd 1 | Herd 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Item | Mean (M) | SD | df | Mean (M) | SD | df | T-Value | p-Value |
| Confined | 0.716 | 1.905 | 21 | 0.295 | 0.255 | 72 | 2.733 | 0.454 |
| Pasture | 0.005 | 0.020 | 12 | 0.000 | 0.000 | 46 | 2.618 | 0.129 |
| Item | Confined | Pasture | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean (M) | SD | df | Mean (M) | SD | df | T-Value | p-Value | |
| Left allogrooming total | 2.2 | 2.024 | 29 | 0.600 | 0.548 | 4 | 2.733 | 0.092 |
| Right allogrooming total | 2.367 | 3.409 | 29 | 0.600 | 0.548 | 4 | 2.733 | 0.261 |
| Herd 1 Left allogrooming | 2.526 | 2.270 | 18 | 0.600 | 0.548 | 4 | 2.819 | 0.077 |
| Herd 1 Right allogrooming | 2.789 | 4.158 | 18 | 0.600 | 0.548 | 4 | 2.819 | 0.260 |
| Herd 2 Left allogrooming | 1.636 | 1.433 | 10 | N/A | N/A | N/A | N/A | N/A |
| Herd 2 Right allogrooming | 1.636 | 1.286 | 10 | N/A | N/A | N/A | N/A | N/A |
| Item | Left Allogrooming | Right Allogrooming | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean (M) | SD | df | Mean (M) | SD | df | T-Value | p-Value | |
| Herd 1 Confined | 2.526 | 2.270 | 18 | 2.789 | 4.158 | 18 | 2.719 | 0.810 |
| Herd 1 Pasture | 0.600 | 0.548 | 4 | 0.600 | 0.548 | 4 | 3.355 | 1.000 |
| Herd 2 Confined | 1.636 | 1.433 | 10 | 1.636 | 1.286 | 10 | 2.845 | 1.000 |
| Herd 2 Pasture | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| Both Herds Confined | 2.200 | 2.024 | 29 | 2.367 | 3.409 | 29 | 2.663 | 0.819 |
| Both Herds Pasture | 0.600 | 0.548 | 4 | 0.600 | 0.548 | 4 | 3.355 | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kieson, E.; Goma, A.A.; Radi, M. Tend and Befriend in Horses: Partner Preferences, Lateralization, and Contextualization of Allogrooming in Two Socially Stable Herds of Quarter Horse Mares. Animals 2023, 13, 225. https://doi.org/10.3390/ani13020225
Kieson E, Goma AA, Radi M. Tend and Befriend in Horses: Partner Preferences, Lateralization, and Contextualization of Allogrooming in Two Socially Stable Herds of Quarter Horse Mares. Animals. 2023; 13(2):225. https://doi.org/10.3390/ani13020225
Chicago/Turabian StyleKieson, Emily, Amira A. Goma, and Medhat Radi. 2023. "Tend and Befriend in Horses: Partner Preferences, Lateralization, and Contextualization of Allogrooming in Two Socially Stable Herds of Quarter Horse Mares" Animals 13, no. 2: 225. https://doi.org/10.3390/ani13020225
APA StyleKieson, E., Goma, A. A., & Radi, M. (2023). Tend and Befriend in Horses: Partner Preferences, Lateralization, and Contextualization of Allogrooming in Two Socially Stable Herds of Quarter Horse Mares. Animals, 13(2), 225. https://doi.org/10.3390/ani13020225