

Dietary Supplementation with Chlorogenic Acid Enhances Antioxidant Capacity, Which Promotes Growth, Jejunum Barrier Function, and Cecum Microbiota in Broilers under High Stocking Density Stress
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Treatment
2.2. Animals and Experimental Design
2.3. Growth Performance and Sample Collection
2.4. Antioxidant Capacity of the Serum
2.5. Intestinal Morphology
2.6. Quantiative Real-Time PCR (qRT-PCR) Analysis
2.7. The Cecal Bacterial Community
2.8. Statistical Analyses
3. Results
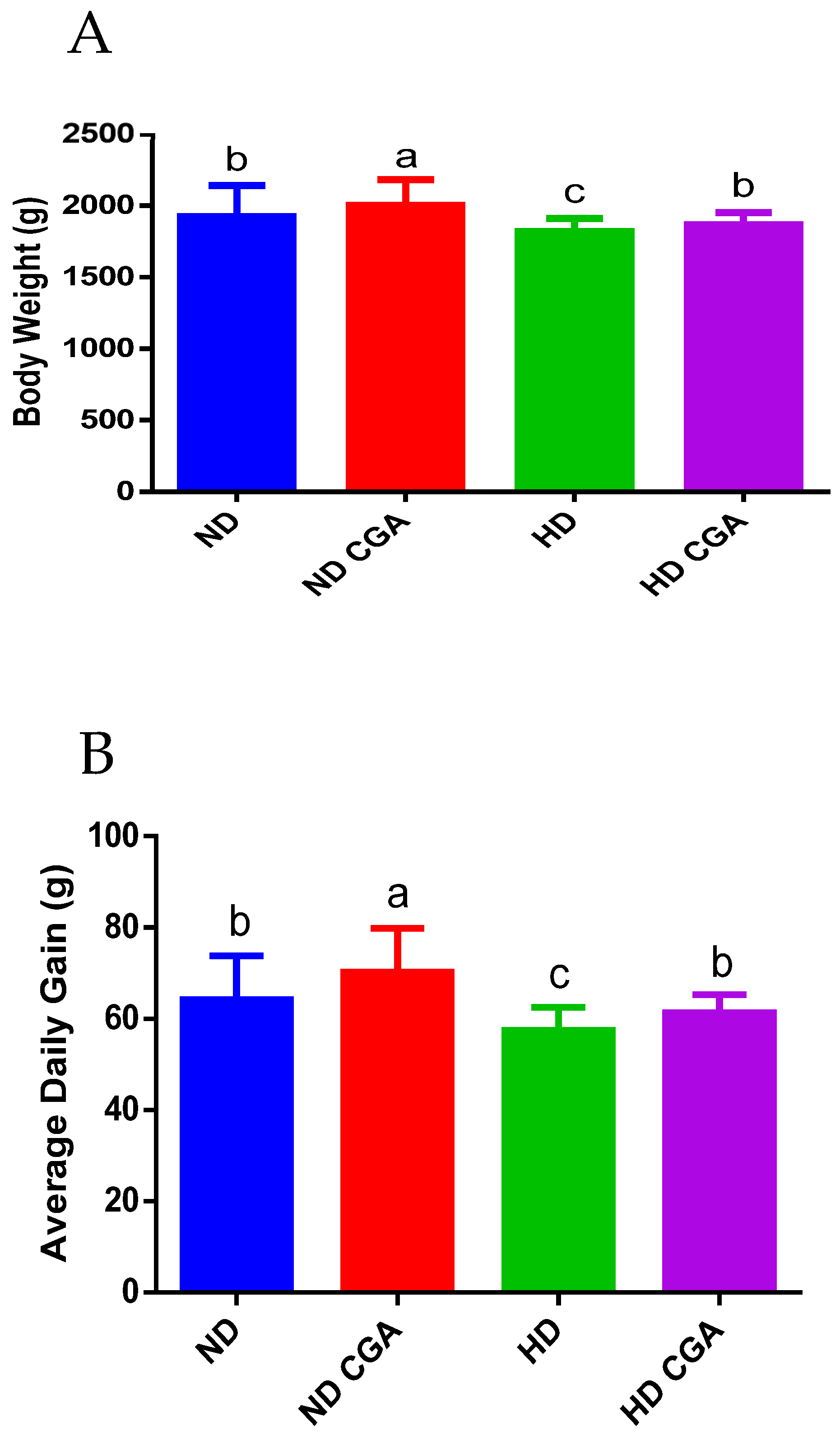
3.1. Effect of CGA on Growth Performance of HD Broilers
3.2. Effect of CGA on Antioxidant Capacity of HD Broilers
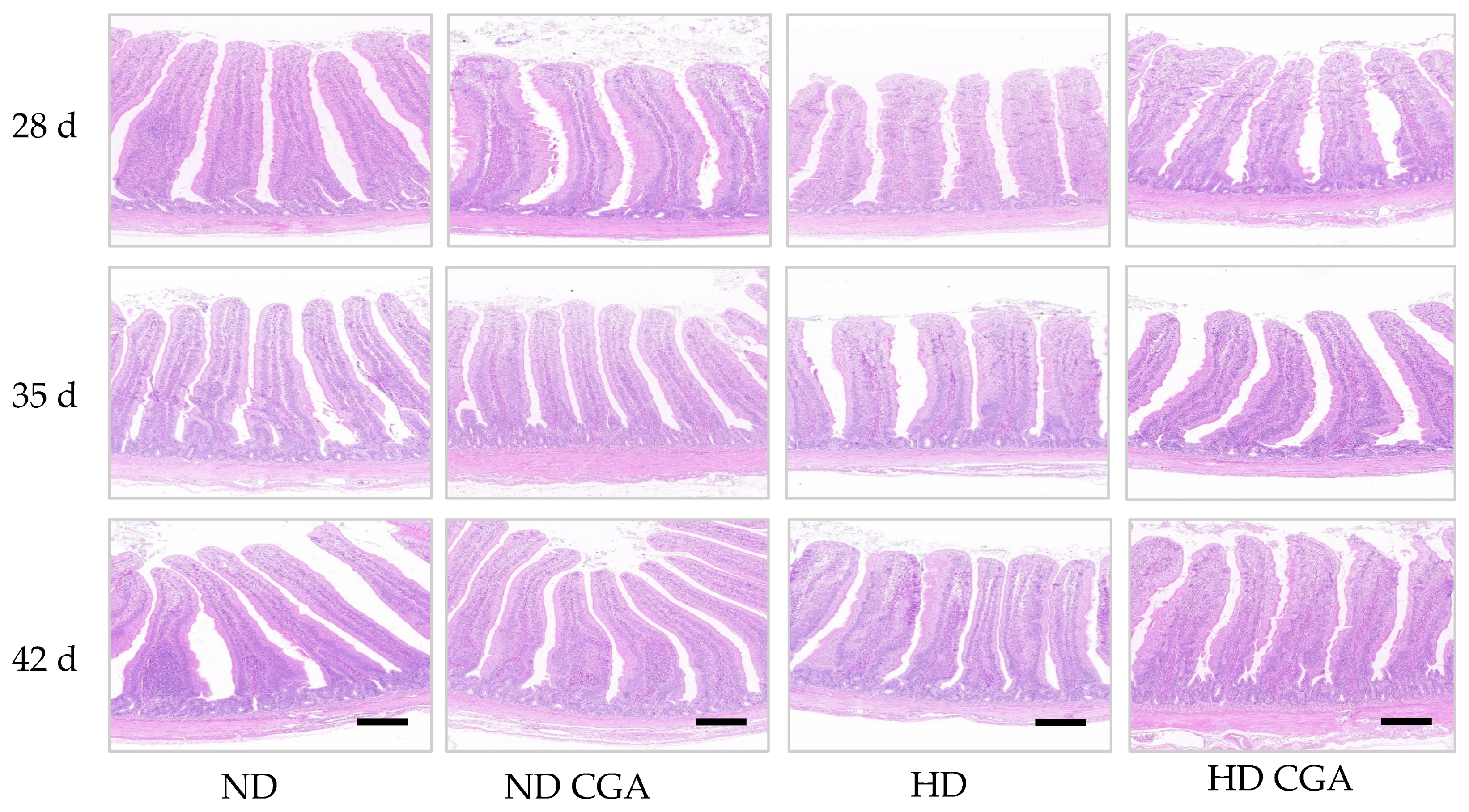
3.3. Effect of CGA on Jejunum Morphology in HD Broilers
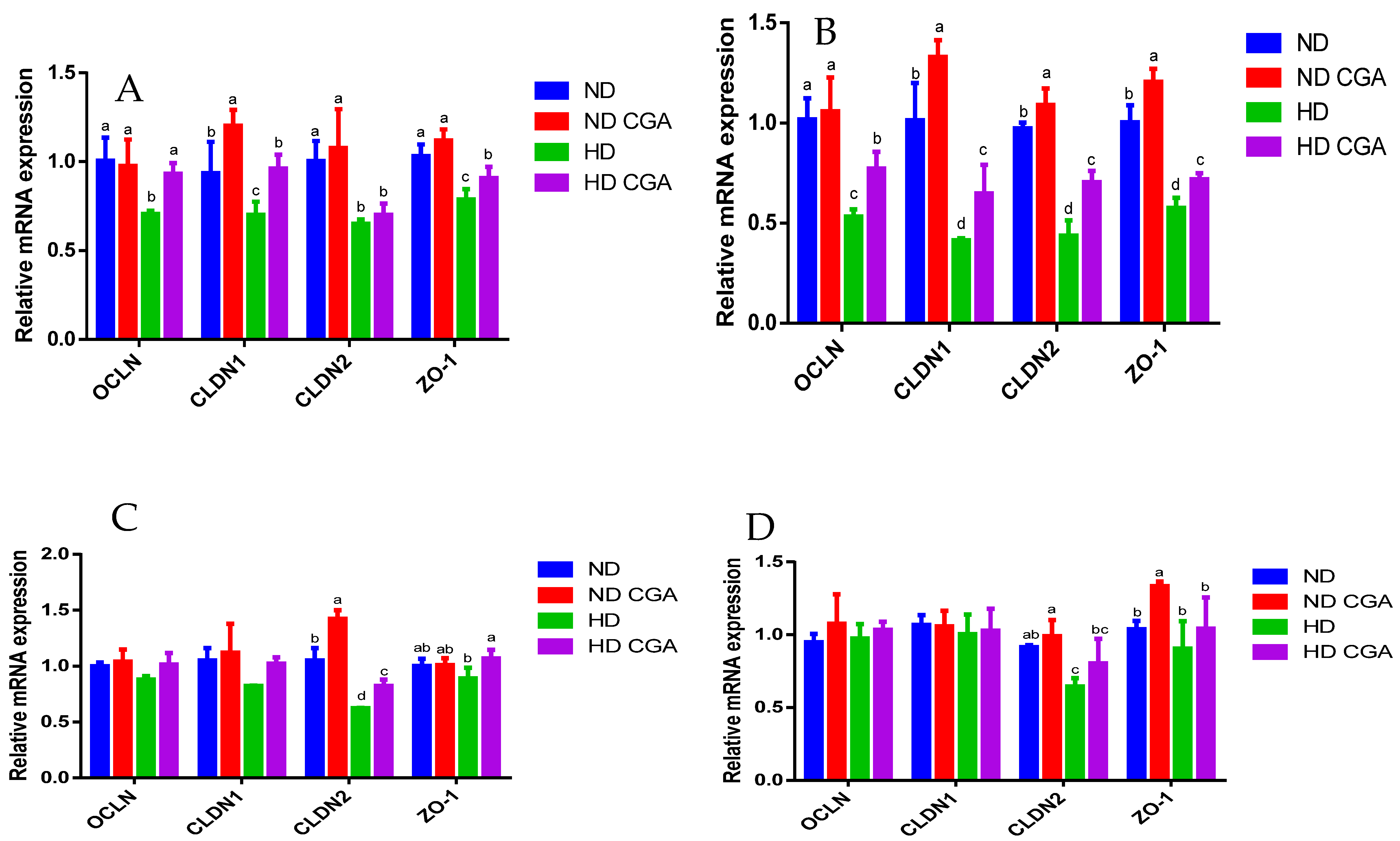
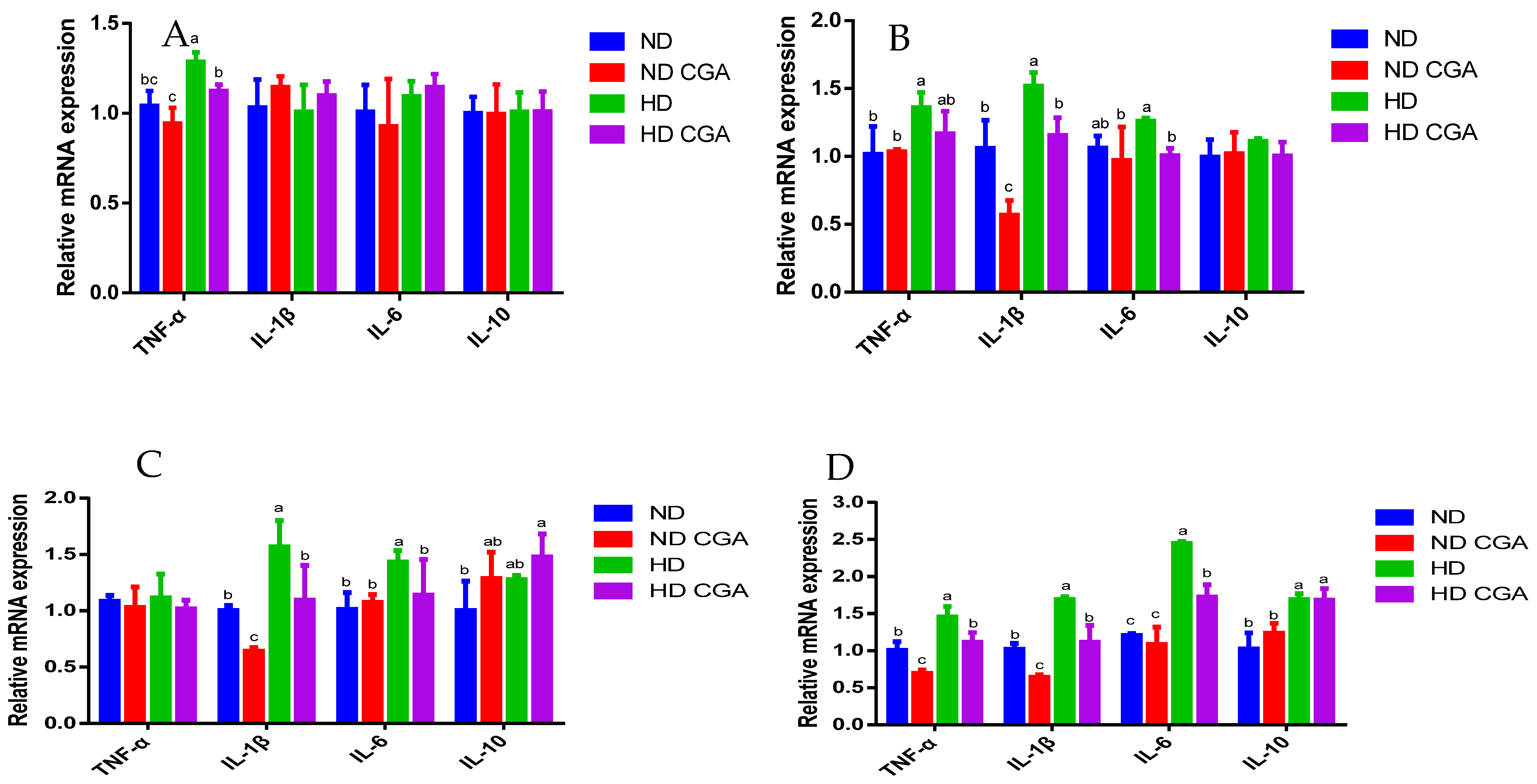
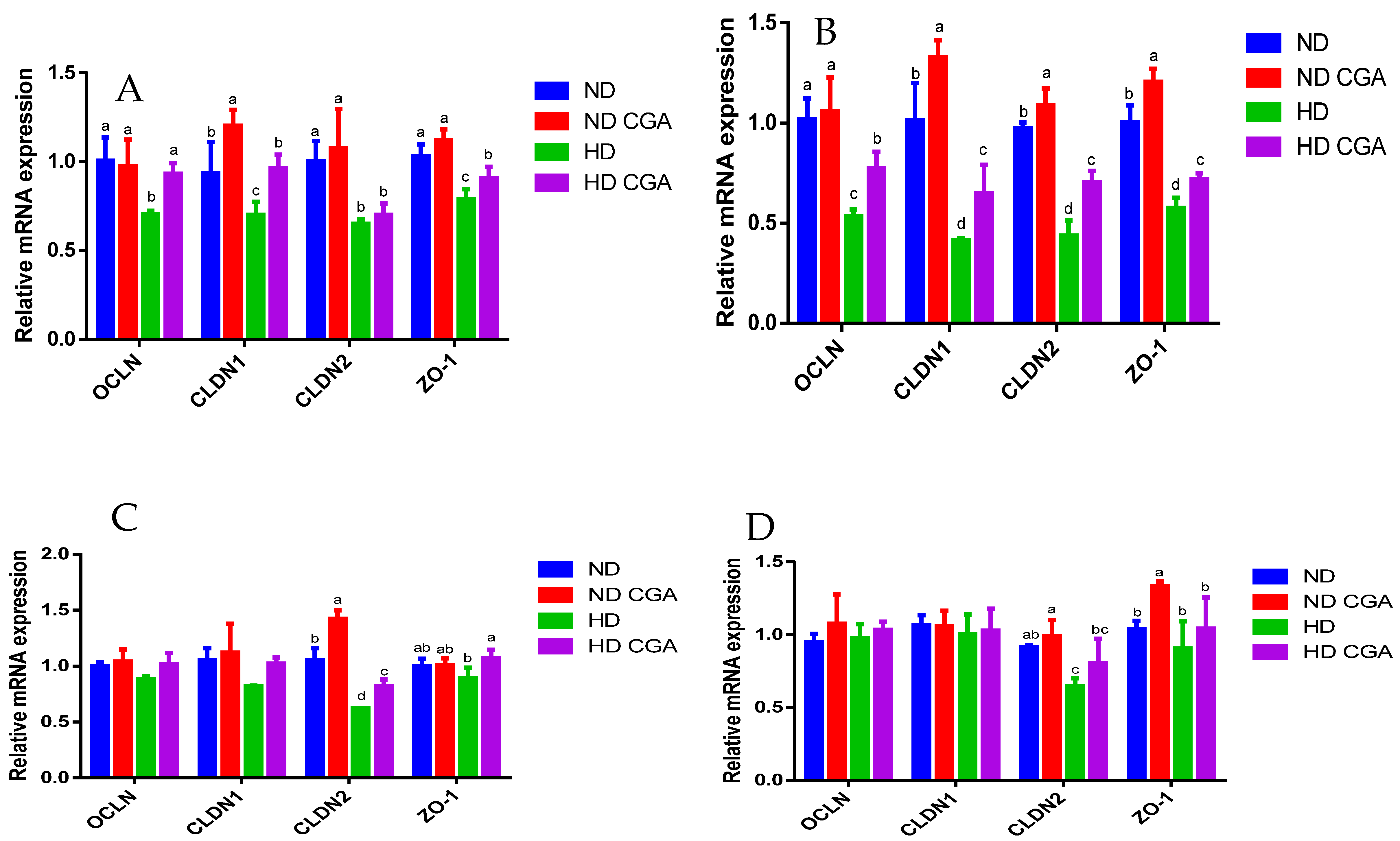
3.4. Effect of CGA on Jejunal mRNA Expression of Tight-Junction Genes and Immune Factors in HD Broilers
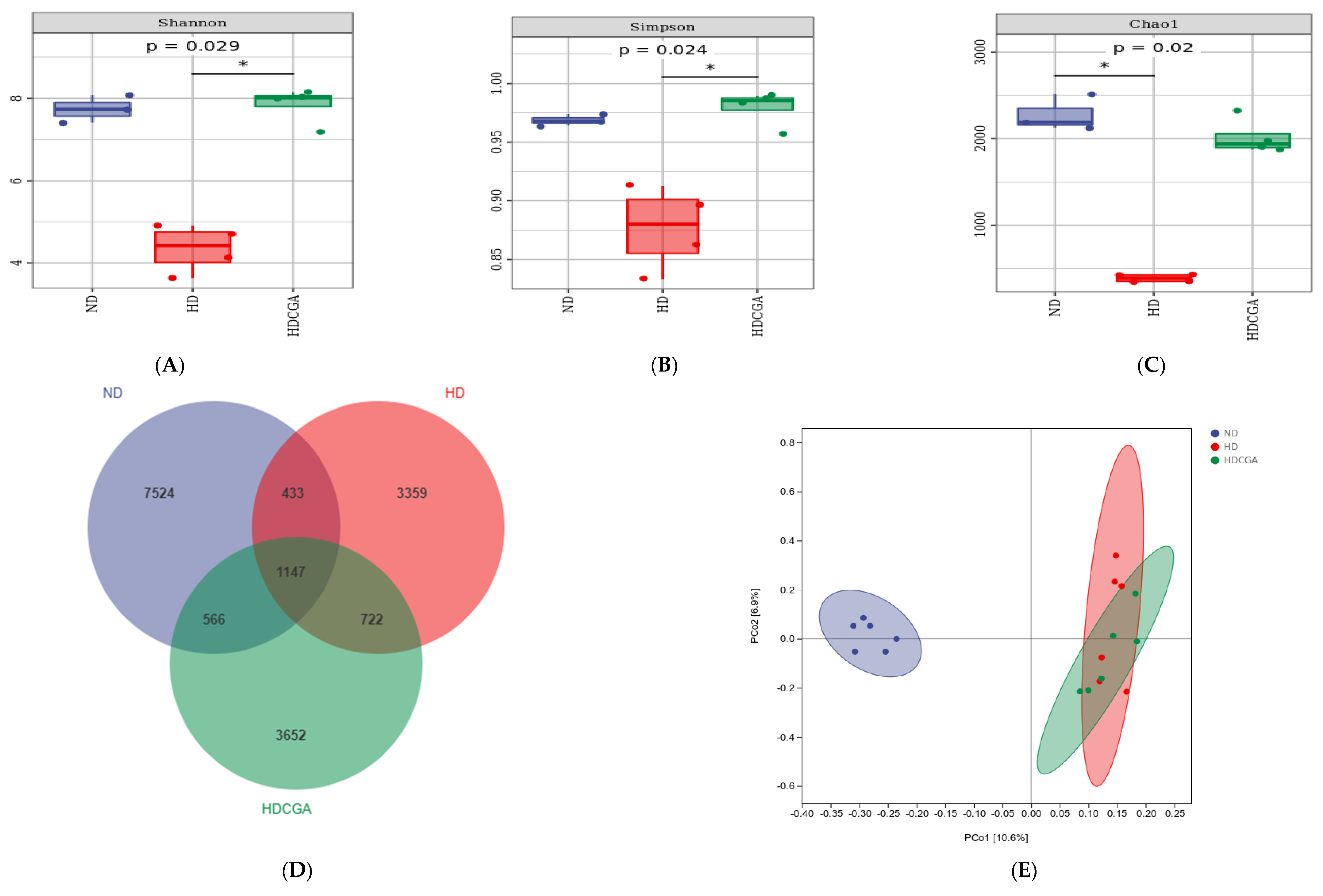
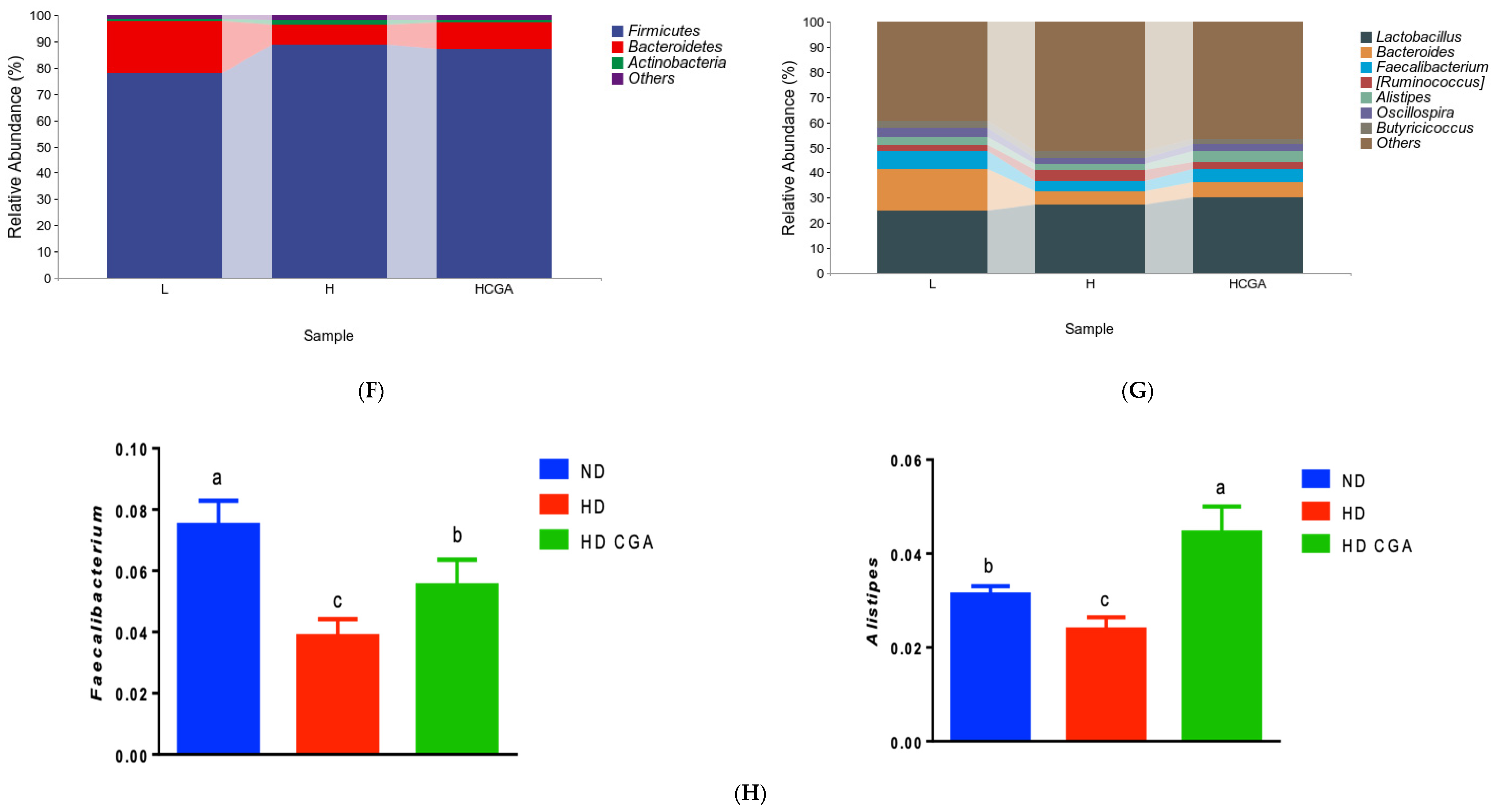
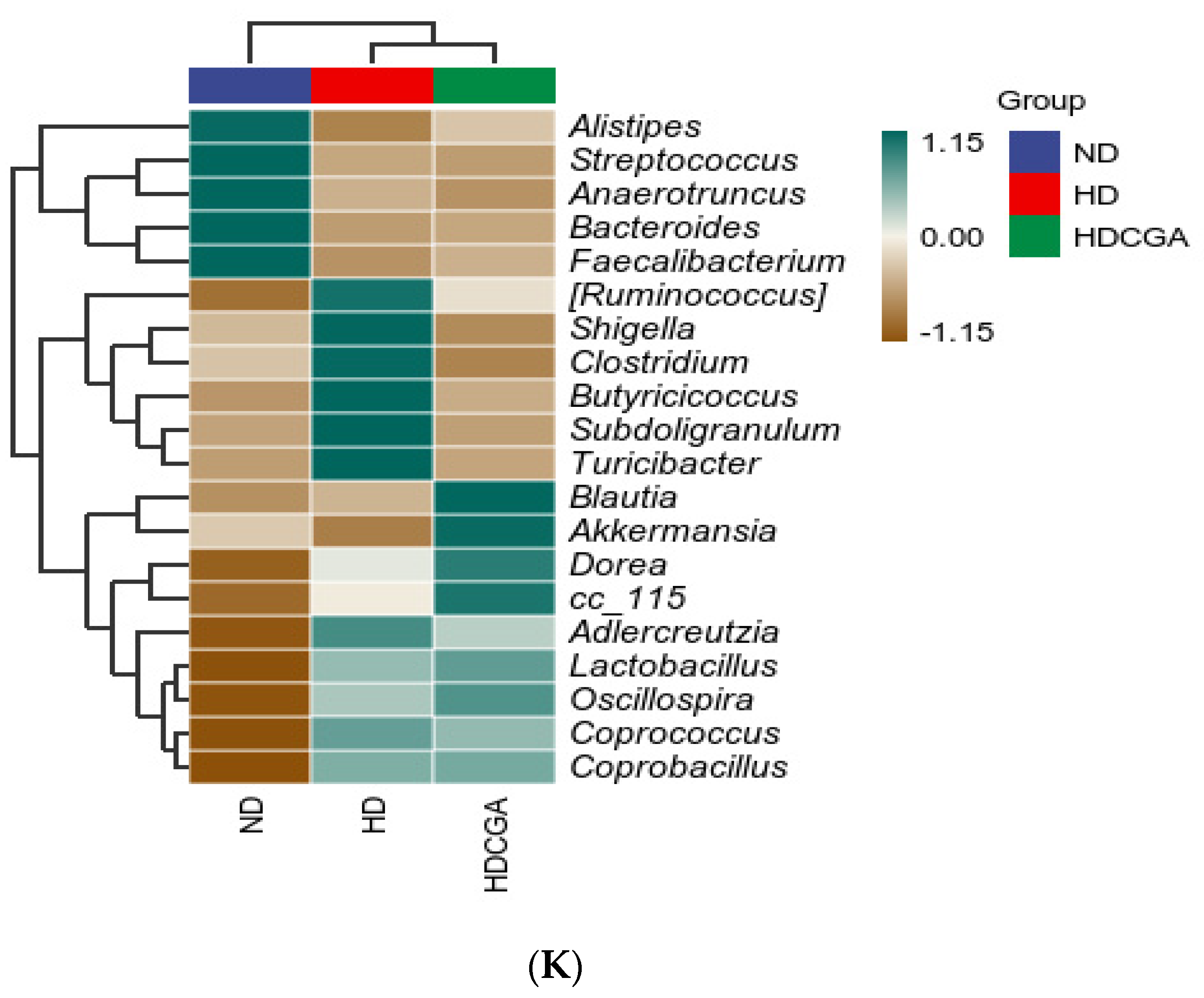
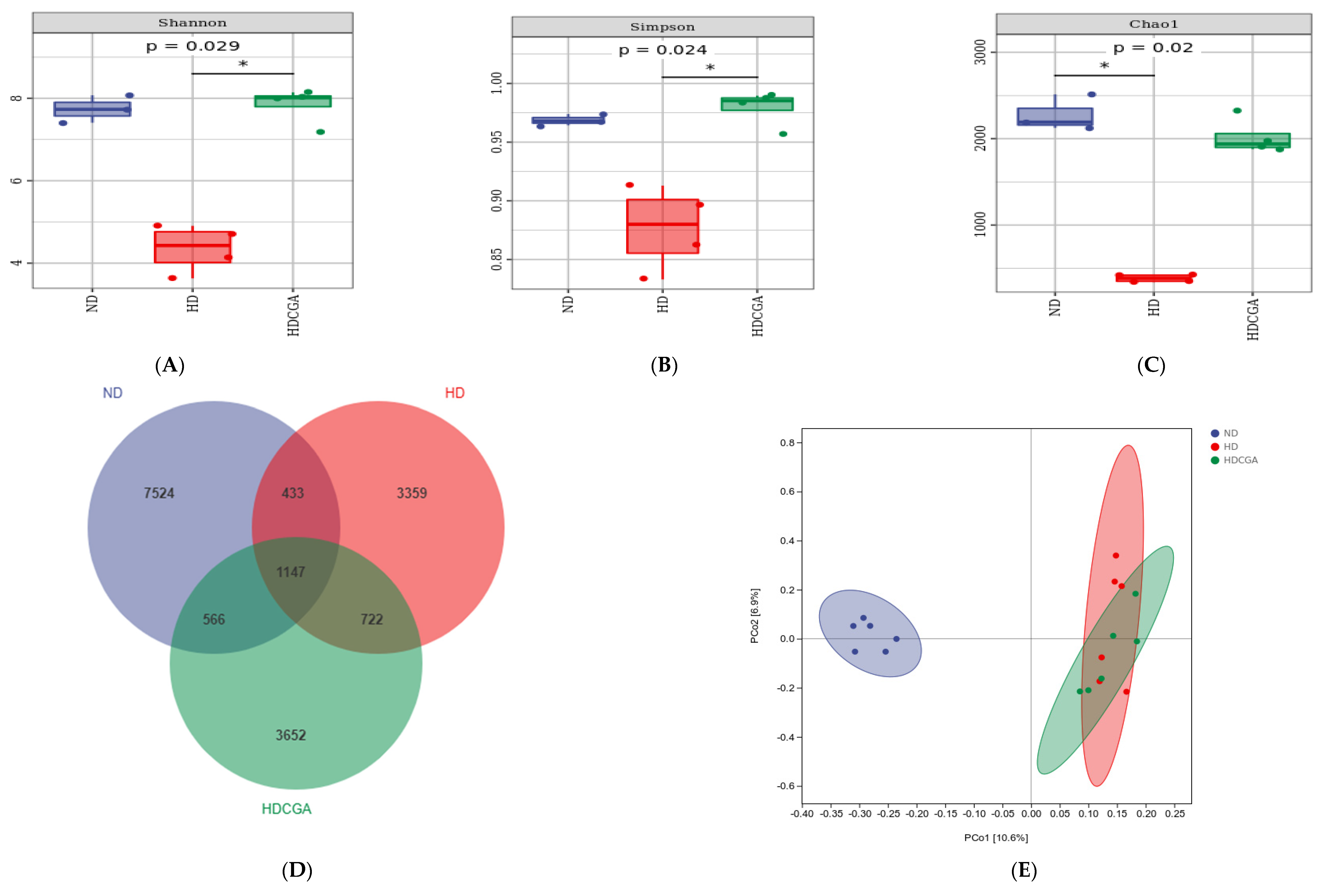
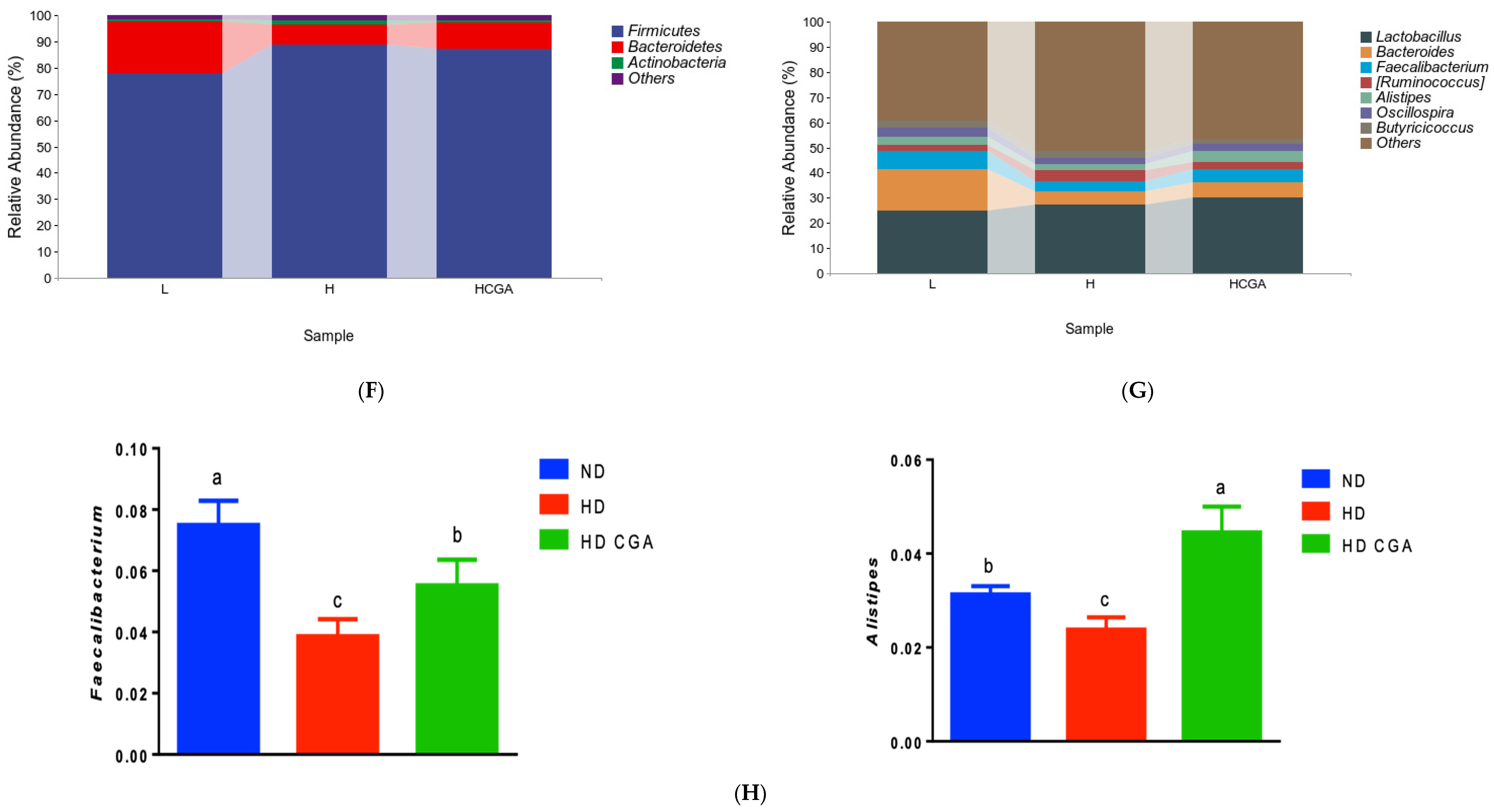
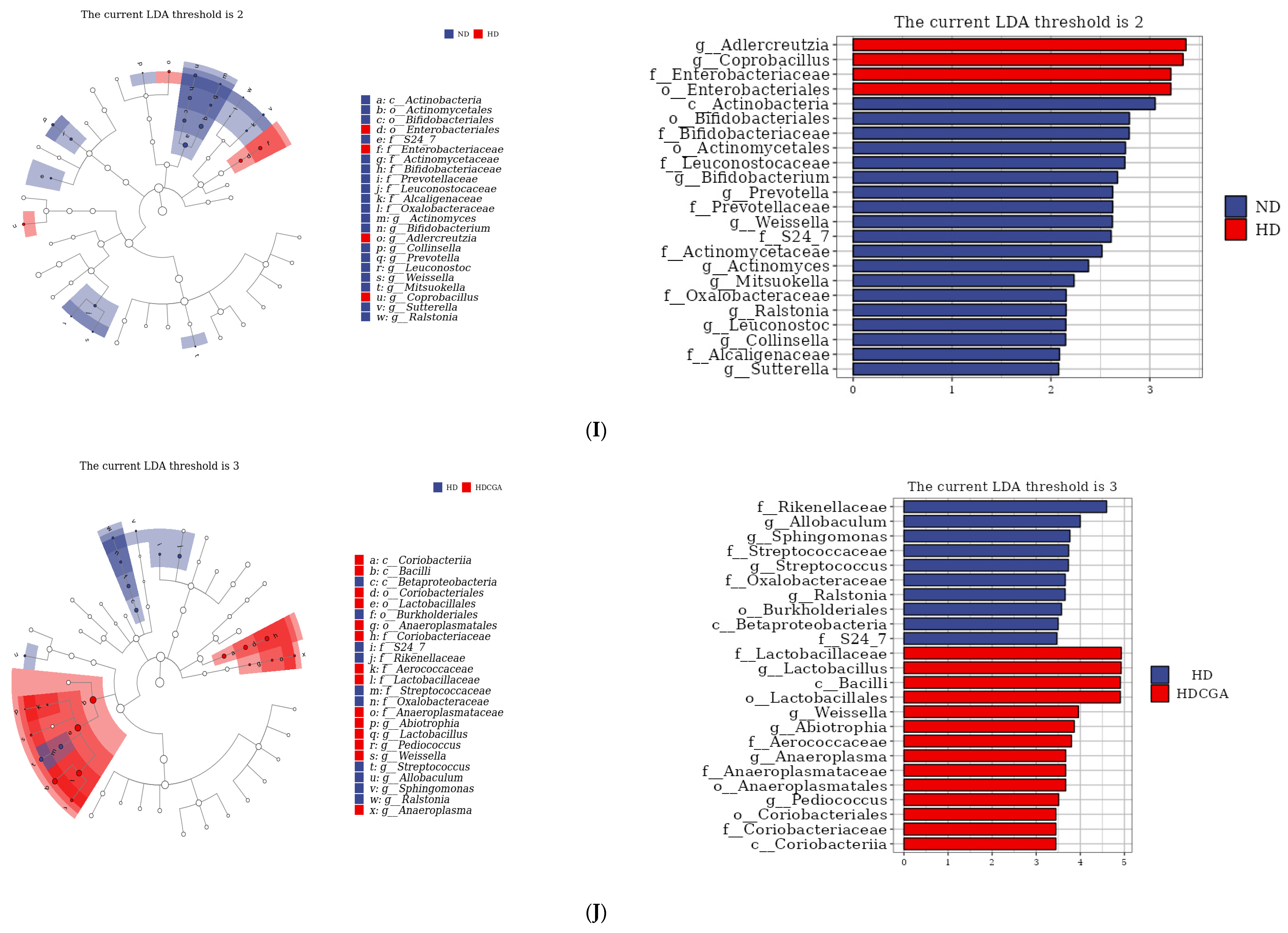
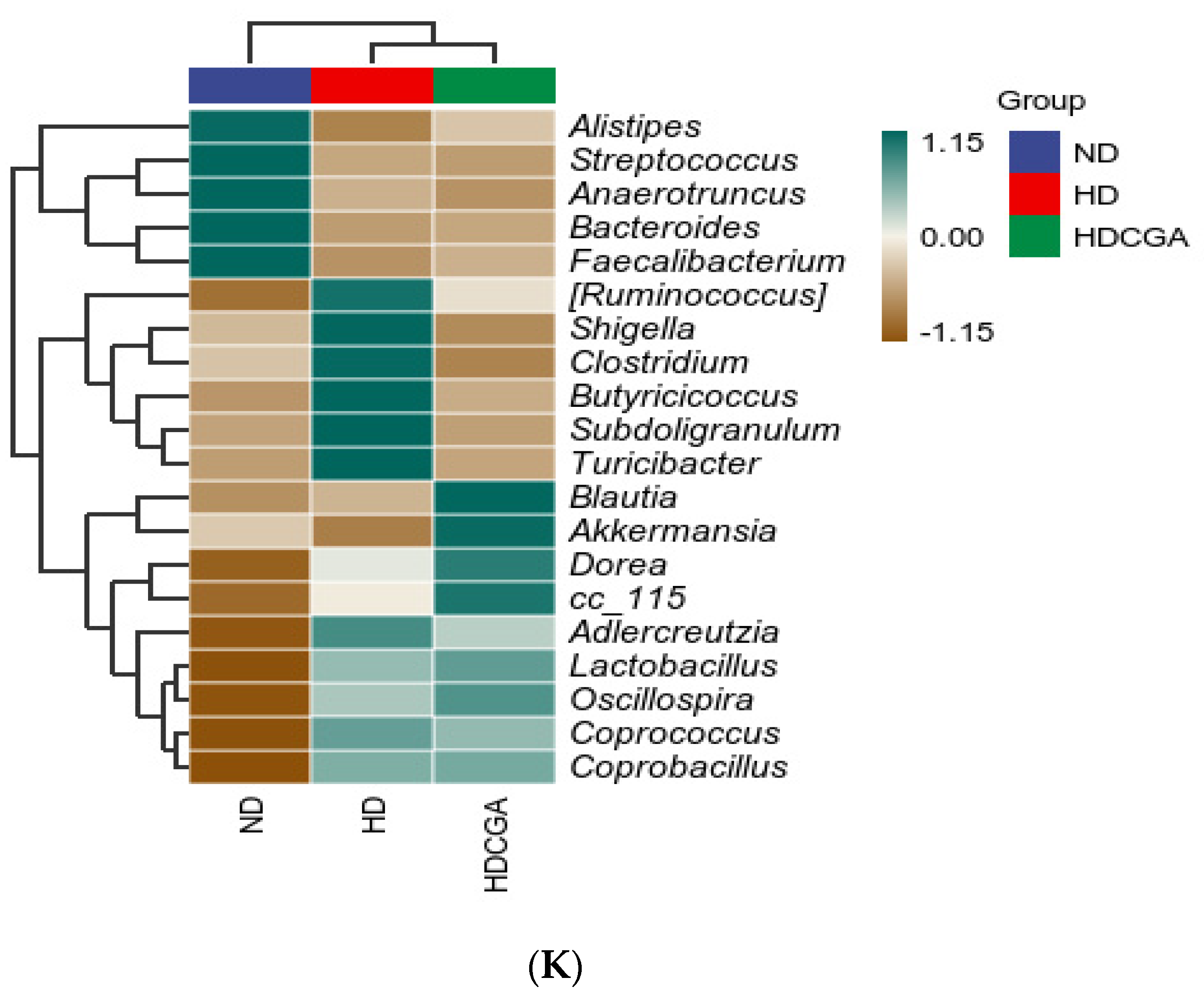
3.5. Effect of CGA on the Composition and Diversity of Cecal Microbial Flora of HD Broilers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Simitzis, P.E.; Kalogeraki, E.; Goliomytis, M.; Charismiadou, M.A.; Triantaphyllopoulos, K.; Ayoutanti, A.; Niforou, K.; Hager-Theodorides, A.L.; Deligeorgis, S.G. Impact of stocking density on broiler growth performance, meat characteristics, behavioural components and indicators of physiological and oxidative stress. Br. Poult. Sci. 2012, 53, 721–730. [Google Scholar] [CrossRef]
- Wang, B.; Min, Z.; Yuan, J.; Zhang, B.; Guo, Y. Effects of dietary tryptophan and stocking density on the performance, meat quality, and metabolic status of broilers. J. Anim. Sci. Biotechnol. 2014, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Son, J.; Kim, H.J.; Hong, E.C.; Kang, H.K. Effects of Stocking Density on Growth Performance, Antioxidant Status, and Meat Quality of Finisher Broiler Chickens under High Temperature. Antioxidants 2022, 11, 871. [Google Scholar] [CrossRef]
- Sugiharto, S. Dietary strategies to alleviate high-stocking-density-induced stress in broiler chickens—A comprehensive review. Arch. Anim. Breed. 2022, 65, 21–36. [Google Scholar] [CrossRef]
- Buijs, S.; Keeling, L.; Rettenbacher, S.; Van Poucke, E.; Tuyttens, F.A. Stocking density effects on broiler welfare: Identifying sensitive ranges for different indicators. Poult. Sci. 2009, 88, 1536–1543. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wei, F.; Xu, B.; Sun, Q.; Deng, W.; Ma, H.; Bai, J.; Li, S. Effect of stocking density and alpha-lipoic acid on the growth performance, physiological and oxidative stress and immune response of broilers. Asian-Australas. J. Anim. Sci. 2019, 32, 1914–1922. [Google Scholar] [CrossRef] [Green Version]
- Nasr, M.; Alkhedaide, A.Q.; Ramadan, A.; Hafez, A.; Hussein, M.A. Potential impact of stocking density on growth, carcass traits, indicators of biochemical and oxidative stress and meat quality of different broiler breeds. Poult. Sci. 2021, 100, 101442. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Kim, G.B.; Cha, C.J. Spatial heterogeneity and stability of bacterial community in the gastrointestinal tracts of broiler chickens. Poult. Sci. 2014, 93, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Zhang, T.; Yan, L.; Qiu, L.; Yin, H.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; et al. Dietary 25-hydroxyvitamin D improves intestinal health and microbiota of laying hens under high stocking density. Poult. Sci. 2021, 100, 101132. [Google Scholar] [CrossRef]
- Wang, W.; Xia, T.; Yu, X. Wogonin suppresses inflammatory response and maintains intestinal barrier function via TLR4-MyD88-TAK1-mediated NF-κB pathway in vitro. Inflamm. Res. 2015, 64, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Guardia, S.; Konsak, B.; Combes, S.; Levenez, F.; Cauquil, L.; Guillot, J.F.; Moreau-Vauzelle, C.; Lessire, M.; Juin, H.; Gabriel, I. Effects of stocking density on the growth performance and digestive microbiota of broiler chickens. Poult. Sci. 2011, 90, 1878–1889. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Piao, X.S.; Zhang, Q.; Li, P.; Yi, J.Q.; Liu, J.D.; Li, Q.Y.; Wang, G.Q. The effects of Forsythia suspensa extract and berberine on growth performance, immunity, antioxidant activities, and intestinal microbiota in broilers under high stocking density. Poult. Sci. 2013, 92, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.Q.; Dong, Y.Y.; Qin, X.; Yuan, J.M.; Han, M.M.; Zhang, K.K.; Shi, S.R.; Song, X.Y.; Zhang, J.Z.; Li, J.H. Dietary supplementation of methionine mitigates oxidative stress in broilers under high stocking density. Poult. Sci. 2021, 100, 101231. [Google Scholar] [CrossRef]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of Chlorogenic Acids in Controlling Oxidative and Inflammatory Stress Conditions. Nutrients 2015, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vukelić, I.; Detel, D.; Pučar, L.B.; Potočnjak, I.; Buljević, S.; Domitrović, R. Chlorogenic acid ameliorates experimental colitis in mice by suppressing signaling pathways involved in inflammatory response and apoptosis. Food Chem. Toxicol. 2018, 121, 140–150. [Google Scholar] [CrossRef]
- Vizzier, T.Y.; Christensen, K.D.; Mench, J.A.; Rumley, E.R.; Daugherty, C.; Feinberg, B.; Parker, M.; Siegel, P.; Scanes, C.G. Symposium: Animal welfare challenges for today and tomorrow. Poult. Sci. 2016, 95, 2198–2207. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L. Natural antioxidants as food and feed additives to promote health benefits and quality of meat products: A review. Meat Sci. 2016, 120, 107–117. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Nutrient Requirement of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Bai, D.; Liu, K.; He, X.; Tan, H.; Liu, Y.; Li, Y.; Zhang, Y.; Zhen, W.; Zhang, C.; Ma, Y. Effect of Dietary Chlorogenic Acid on Growth Performance, Antioxidant Function, and Immune Response of Broiler Breeders under Immune Stress and Stocking Density Stress. Vet. Sci. 2022, 9, 582. [Google Scholar] [CrossRef]
- Zhen, W.R.; Shao, Y.J.; Wu, Y.Y.; Li, L.H.; Pham, V.H.; Abbas, W.; Guo, Y.; Wang, Z. Dietary yeast beta-glucan supplementation improves eggshell color and fertile eggs hatchability as well as enhances immune functions in breeder laying hens. Int. J. Biol. Macromol. 2020, 159, 607–621. [Google Scholar] [CrossRef]
- Zhen, W.; Liu, Y.; Shao, Y.; Ma, Y.; Wu, Y.; Guo, F.; Abbas, W.; Guo, Y.; Wang, Z. Yeast β-Glucan Altered Intestinal Microbiome and Metabolome in Older Hens. Front. Microbiol. 2021, 12, 766878. [Google Scholar] [CrossRef]
- Goo, D.; Kim, J.H.; Park, G.H.; Delos, R.J.; Kil, D.Y. Effect of stocking density and dietary tryptophan on growth performance and intestinal barrier function in broiler chickens. Poult. Sci. 2019, 98, 4504–4508. [Google Scholar] [CrossRef]
- Dai, D.; Qi, G.; Wang, J.; Zhang, H.; Qiu, K.; Han, Y.; Wu, Y.; Wu, S. Dietary organic acids ameliorate high stocking density stress-induced intestinal inflammation through the restoration of intestinal microbiota in broilers. J. Anim. Sci. Biotechnol. 2022, 13, 124. [Google Scholar] [CrossRef]
- Estevez, I. Density allowances for broilers: Where to set the limits? Poult. Sci. 2007, 86, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yu, Z.; Hou, J.; Deng, Y.; Zhou, Z.; Zhao, Z.; Cui, J. Toxicity and oxidative stress induced by T-2 toxin and HT-2 toxin in broilers and broiler hepatocytes. Food Chem. Toxicol. 2016, 87, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, H.; Zhao, N.; Yang, X.; Du, E.; Huang, S.; Guo, W.; Zhang, W.; Wei, J. Effect of chlorogenic acid on intestinal inflammation, antioxidant status, and microbial community of young hens challenged with acute heat stress. Anim. Sci. J. 2021, 92, e13619. [Google Scholar] [CrossRef]
- Mishra, B.; Jha, R. Oxidative Stress in the Poultry Gut: Potential Challenges and Interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhao, Q.; Ci, X.; Chen, S.; Xie, Z.; Li, H.; Zhang, H.; Chen, F.; Xie, Q. Evaluation of the efficacy of chlorogenic acid in reducing small intestine injury, oxidative stress, and inflammation in chickens challenged with Clostridium perfringens type A. Poult. Sci. 2020, 99, 6606–6618. [Google Scholar] [CrossRef]
- Goo, D.; Kim, J.H.; Choi, H.S.; Park, G.H.; Han, G.P.; Kil, D.Y. Effect of stocking density and sex on growth performance, meat quality, and intestinal barrier function in broiler chickens. Poult. Sci. 2019, 98, 1153–1160. [Google Scholar] [CrossRef]
- Yu, D.G.; Namgung, N.; Kim, J.H.; Won, S.Y.; Choi, W.J.; Kil, D.Y. Effects of stocking density and dietary vitamin C on performance, meat quality, intestinal permeability, and stress indicators in broiler chickens. J. Anim. Sci. Technol. 2021, 63, 815–826. [Google Scholar] [CrossRef]
- Salvo, R.E.; Alonso, C.C.; Pardo, C.C.; Casado, B.M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Gilani, S.; Howarth, G.S.; Nattrass, G.; Kitessa, S.M.; Barekatain, R.; Forder, R.; Tran, C.D.; Hughes, R.J. Gene expression and morphological changes in the intestinal mucosa associated with increased permeability induced by short-term fasting in chickens. J. Anim. Physiol. Anim. Nutr. 2018, 102, e653–e661. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H. Cytokines and prevention of infectious diseases in poultry: A review. Avian Pathol. 2000, 29, 395–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cengiz, Ö.; Köksal, B.H.; Tatlı, O.; Sevim, Ö.; Ahsan, U.; Üner, A.G.; Ulutaş, P.A.; Beyaz, D.; Büyükyörük, S.; Yakan, A.; et al. Effect of dietary probiotic and high stocking density on the performance, carcass yield, gut microflora, and stress indicators of broilers. Poult. Sci. 2015, 94, 2395–2403. [Google Scholar] [CrossRef]
- Xin, X.; Han, M.; Wu, Y.; Dong, Y.; Miao, Z.; Zhang, J.; Song, X.; Jia, R.; Su, Y.; Liu, C.; et al. Dietary Supplemental Chromium Yeast Improved the Antioxidant Capacity, Immunity and Liver Health in Broilers under High Stocking Density. Animals 2022, 12, 2216. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Niu, J.; Jing, C.; Jiao, N.; Huang, L.; Jiang, S.; Yan, L.; Yang, W.; Li, Y. Supplementation with paraformic acid in the diet improved intestinal development through modulating intestinal inflammation and microbiota in broiler chickens. Front. Microbiol. 2022, 13, 975056. [Google Scholar] [CrossRef]
- Song, Z.; Cheng, K.; Zhang, L.; Wang, T. Dietary supplementation of enzymatically treated Artemisia annua could alleviate the intestinal inflammatory response in heat-stressed broilers. J. Therm. Biol. 2017, 69, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhao, L.; Xu, Z.J.; De Marco, M.; Briens, M.; Yan, X.H.; Sun, L.H. Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response. Antioxidants 2021, 10, 1542. [Google Scholar] [CrossRef]
- Wan, F.; Zhong, R.; Wang, M.; Zhou, Y.; Chen, Y.; Yi, B.; Hou, F.; Liu, L.; Zhao, Y.; Chen, L.; et al. Caffeic Acid Supplement Alleviates Colonic Inflammation and Oxidative Stress Potentially through Improved Gut Microbiota Community in Mice. Front. Microbiol. 2021, 12, 784211. [Google Scholar] [CrossRef] [PubMed]
- Dozier, W.R.; Thaxton, J.P.; Purswell, J.L.; Olanrewaju, H.A.; Branton, S.L.; Roush, W.B. Stocking density effects on male broilers grown to 1.8 kilograms of body weight. Poult. Sci. 2006, 85, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S.; Donnelly, C.A.; Jones, T.A. Chicken welfare is influenced more by housing conditions than by stocking density. Nature 2004, 427, 342–344. [Google Scholar] [CrossRef]
- Ouyang, W.; O’Garra, A. IL-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhang, B.; Zhao, X.; Lin, Y.; Wang, J.; Wang, X.; Hu, N.; Wang, S. Chlorogenic acid supplementation ameliorates hyperuricemia, relieves renal inflammation, and modulates intestinal homeostasis. Food Funct. 2021, 12, 5637–5649. [Google Scholar] [CrossRef]
- Sekelja, M.; Rud, I.; Knutsen, S.H.; Denstadli, V.; Westereng, B.; Næs, T.; Rudi, K. Abrupt temporal fluctuations in the chicken fecal microbiota are explained by its gastrointestinal origin. Appl. Environ. Microbiol. 2012, 78, 2941–2948. [Google Scholar] [CrossRef] [Green Version]
- Kridtayopas, C.; Rakangtong, C.; Bunchasak, C.; Loongyai, W. Effect of prebiotic and synbiotic supplementation in diet on growth performance, small intestinal morphology, stress, and bacterial population under high stocking density condition of broiler chickens. Poult. Sci. 2019, 98, 4595–4605. [Google Scholar] [CrossRef]
- Chen, J.; Yu, B.; Chen, D.; Zheng, P.; Luo, Y.; Huang, Z.; Luo, J.; Mao, X.; Yu, J.; He, J. Changes of porcine gut microbiota in response to dietary chlorogenic acid supplementation. Appl. Microbiol. Biotechnol. 2019, 103, 8157–8168. [Google Scholar] [CrossRef]
- Danzeisen, J.L.; Kim, H.B.; Isaacson, R.E.; Tu, Z.J.; Johnson, T.J. Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE 2011, 6, e27949. [Google Scholar] [CrossRef]
- Xi, Y.; Li, H.; Yu, M.; Li, X.; Li, Y.; Hui, B.; Zeng, X.; Wang, J.; Li, J. Protective effects of chlorogenic acid on trimethyltin chloride-induced neurobehavioral dysfunctions in mice relying on the gut microbiota. Food Funct. 2022, 13, 1535–1550. [Google Scholar] [CrossRef]
- Teixeira, C.G.; Fusieger, A.; Milião, G.L.; Martins, E.; Drider, D.; Nero, L.A.; de Carvalho, A.F. Weissella: An Emerging Bacterium with Promising Health Benefits. Probiotics Antimicrob. Proteins 2021, 13, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, L.; Chen, R.; Zhao, Y.; Ren, D.; Yang, X. Chlorogenic acid inhibits trimethylamine-N-oxide formation and remodels intestinal microbiota to alleviate liver dysfunction in high L-carnitine feeding mice. Food Funct. 2021, 12, 10500–10511. [Google Scholar] [CrossRef]
- Xu, J.; Liang, R.; Zhang, W.; Tian, K.; Li, J.; Chen, X.; Yu, T.; Chen, Q. Faecalibacterium prausnitzii-derived microbial anti-inflammatory molecule regulates intestinal integrity in diabetes mellitus mice via modulating tight junction protein expression. J. Diabetes 2020, 12, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria with Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and disease markers correlate with gut microbiome composition across thousands of people. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef] [PubMed]
- Henke, M.T.; Brown, E.M.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Capsular polysaccharide correlates with immune response to the human gut microbe Ruminococcus gnavus. Proc. Natl. Acad. Sci. USA 2021, 118, e2007595118. [Google Scholar] [CrossRef]
- Nisa, I.; Qasim, M.; Yasin, N.; Ullah, R.; Ali, A. Shigella flexneri: An emerging pathogen. Folia Microbiol. 2020, 65, 275–291. [Google Scholar] [CrossRef]
- Shad, A.A.; Shad, W.A. Shigella sonnei: Virulence and antibiotic resistance. Arch. Microbiol. 2021, 203, 45–58. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | Starter (1–21 d) | Grower (21–42 d) |
|---|---|---|
| Corn | 527.9 | 577.8 |
| Soybean meal | 368.9 | 300.0 |
| Zea gluten meal | 0 | 24.3 |
| Soybean oil | 40.0 | 40.0 |
| Sodium chloride | 3.0 | 3.0 |
| Choline chloride | 3.0 | 2.6 |
| Vitamin premix 2 | 0.3 | 0.3 |
| Trace element premix 1 | 2.0 | 2.0 |
| Stone powder | 12.2 | 11.7 |
| Dicalcium phosphate | 19.1 | 16.2 |
| DL-Methionine | 2.7 | 1.1 |
| L-Lysine | 0.4 | 0.45 |
| Wheat bran | 20.0 | 20.0 |
| Total | 1000 | 1000 |
| Metabolic energy (MJ/kg) | 12.4 | 13.0 |
| Crude protein | 211.8 | 198.4 |
| Lysine | 11.4 | 10.5 |
| Methionine | 4.9 | 4.8 |
| Calcium | 10.2 | 8.5 |
| Available P | 4.5 | 4.2 |
| Total P | 6.9 | 6.3 |
| Threonine | 7.7 | 2.2 |
| Analyzed content | ||
| Calcium | 10.2 | 8.5 |
| Total P | 6.8 | 6.2 |
| Calcium: Total P 3 | 1.50 | 1.37 |
| Genes 1 | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Length | TM 2 °C | Accession No |
|---|---|---|---|---|---|
| OCLN | ACGGCAGCACCTACCTCAA | GGGCGAAGAAGCAGATGAG | 123 | 51.7 | XM_025144247.2 |
| CLDN1 | CATACTCCTGGGTCTGGTTGGT | GACAGCCA TCCGCA TCTTCT | 100 | 51.3 | NM_001013611.2 |
| CLDN2 | CCTACATTGGTTCAAGCATCGTGA | GATGTCGGGAGGCAGGTTGA | 131 | 50.3 | NM_001277622.1 |
| ZO-1 | CTTCAGGTGTTTCTCTTCCTCCTC | CTGTGGTTTCA TGGCTGGATC | 144 | 51.5 | XM_021098886.1 |
| TNF-α | GAGCGTTGACTTGGCTGTC | AAGCAACAACCAGCTA TGCAC | 176 | 55.4 | NM_214022.1 |
| IL-1β | ACTGGGCA TCAAGGGCTA | GGTAGAAGA TGAAGCGGGTC | 154 | 55.6 | NM_214005.1 |
| IL-6 | GCTGCGCTTCTACACAGA | TCCCGTTCTCA TCCA TCTTCTC | 203 | 55.4 | NM_204628.1 |
| IL-10 | AGAAATCCCTCCTCGCCAAT | AAATAGCGAACGGCCCTCA | 121 | 51.2 | NM_001004414.2 |
| GAPDH | TGCTGCCCAGAACATCATCC | ACGGCAGGTCAGGTCAACAA | 142 | 50–60 | NM_204305 |
| Parameter | Days | Dietary Treatment 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| ND | ND + CGA | HD | HD + CGA | ||||
| MDA, (nmol/mL) | 21 | 3.56 b | 3.52 b | 5.19 a | 4.04 b | 0.22 | 0.01 |
| 28 | 4.49 b | 4.57 b | 5.88 a | 4.73 b | 0.20 | 0.03 | |
| 35 | 4.96 a | 3.67 b | 5.59 a | 3.41 b | 0.25 | <0.01 | |
| 42 | 4.33 | 4.76 | 5.54 | 3.91 | 0.27 | 0.16 | |
| SOD, (U/mL) | 21 | 148.58 | 148.13 | 146.91 | 147.37 | 3.55 | 0.14 |
| 28 | 162.02 a | 154.63 ab | 146.70 b | 152.83 ab | 2.33 | 0.04 | |
| 35 | 150.43 ab | 158.70 a | 147.56 b | 156.89 a | 2.22 | 0.03 | |
| 42 | 156.96 a | 147.24 a | 126.91 b | 145.82 a | 3.92 | 0.03 | |
| GSH-Px, (U/mL) | 21 | 975.78 a | 954.36 a | 674.99 b | 632.35 b | 9.44 | 0.03 |
| 28 | 911.09 b | 936.76 ab | 825.41 c | 984.84 a | 14.75 | <0.01 | |
| 35 | 764.49 b | 992.03 a | 717.75 b | 823.66 a | 20.92 | 0.02 | |
| 42 | 910.00 ab | 990.15 a | 606.69 b | 992.31 a | 15.92 | <0.01 | |
| CAT, (U/mL) | 21 | 5.25 | 5.83 | 5.27 | 5.88 | 0.16 | 0.33 |
| 28 | 3.83 a | 4.29 a | 2.62 b | 1.19 b | 0.08 | <0.01 | |
| 35 | 4.70 | 5.04 | 3.45 | 1.64 | 0.15 | 0.58 | |
| 42 | 5.49 b | 6.64 a | 5.44 b | 6.90 a | 0.30 | <0.01 | |
| Parameter | Days | Dietary Treatment 1 | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|
| ND | ND + CGA | HD | HD + CGA | ||||
| VH (µm) | 21 | 1034.03 a | 1023.68 a | 852.55 b | 946.17 ab | 21.43 | <0.01 |
| 28 | 1179.68 b | 1290.67 a | 976.53 c | 1122.33 b | 25.38 | <0.01 | |
| 35 | 1335.00 ab | 1407.33 a | 1293.00 b | 1366.50 ab | 14.11 | 0.02 | |
| 42 | 1477.33 | 1450.83 | 1435.50 | 1441.50 | 27.07 | 0.39 | |
| CD (µm) | 21 | 177.83 b | 171.77 b | 188.10 a | 182.37 a | 11.10 | 0.05 |
| 28 | 179.36 a | 173.28 b | 181.00 a | 175.50 ab | 23.13 | 0.04 | |
| 35 | 171.73 | 172.61 | 168.65 | 167.80 | 19.13 | 0.38 | |
| 42 | 167.83 b | 167.77 b | 174.10 a | 173.37 a | 20.11 | 0.05 | |
| VH/CD | 21 | 5.73 a | 5.67 a | 4.56 b | 5.07 b | 0.13 | <0.01 |
| 28 | 6.74 b | 7.71 a | 5.74 c | 6.39 b | 0.18 | <0.01 | |
| 35 | 7.78 | 8.16 | 7.67 | 8.15 | 0.09 | 0.13 | |
| 42 | 8.81 a | 8.65 a | 8.24 b | 8.31 ab | 0.08 | 0.02 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhang, Y.; Bai, D.; Li, Y.; He, X.; Ito, K.; Liu, K.; Tan, H.; Zhen, W.; Zhang, B.; et al. Dietary Supplementation with Chlorogenic Acid Enhances Antioxidant Capacity, Which Promotes Growth, Jejunum Barrier Function, and Cecum Microbiota in Broilers under High Stocking Density Stress. Animals 2023, 13, 303. https://doi.org/10.3390/ani13020303
Liu Y, Zhang Y, Bai D, Li Y, He X, Ito K, Liu K, Tan H, Zhen W, Zhang B, et al. Dietary Supplementation with Chlorogenic Acid Enhances Antioxidant Capacity, Which Promotes Growth, Jejunum Barrier Function, and Cecum Microbiota in Broilers under High Stocking Density Stress. Animals. 2023; 13(2):303. https://doi.org/10.3390/ani13020303
Chicago/Turabian StyleLiu, Yanhao, Yi Zhang, Dongying Bai, Yuqian Li, Xianglong He, Koichi Ito, Kexin Liu, Haiqiu Tan, Wenrui Zhen, Bingkun Zhang, and et al. 2023. "Dietary Supplementation with Chlorogenic Acid Enhances Antioxidant Capacity, Which Promotes Growth, Jejunum Barrier Function, and Cecum Microbiota in Broilers under High Stocking Density Stress" Animals 13, no. 2: 303. https://doi.org/10.3390/ani13020303
APA StyleLiu, Y., Zhang, Y., Bai, D., Li, Y., He, X., Ito, K., Liu, K., Tan, H., Zhen, W., Zhang, B., & Ma, Y. (2023). Dietary Supplementation with Chlorogenic Acid Enhances Antioxidant Capacity, Which Promotes Growth, Jejunum Barrier Function, and Cecum Microbiota in Broilers under High Stocking Density Stress. Animals, 13(2), 303. https://doi.org/10.3390/ani13020303