
Cytotoxicity and Antimicrobial Resistance of Salmonella enterica Subspecies Isolated from Raised Reptiles in Beijing, China

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Selection and Sampling
2.3. Sample Collection and Processing
2.4. Multiplex PCR Assay
2.5. Evaluation of Cytotoxic Activity on Human Cells
2.6. Antimicrobial Resistance Evaluation
2.7. Whole Genome Sequencing (WGS)
2.8. Statistical Analysis
3. Results
3.1. Prevalence of Salmonella spp. in Screened Reptiles
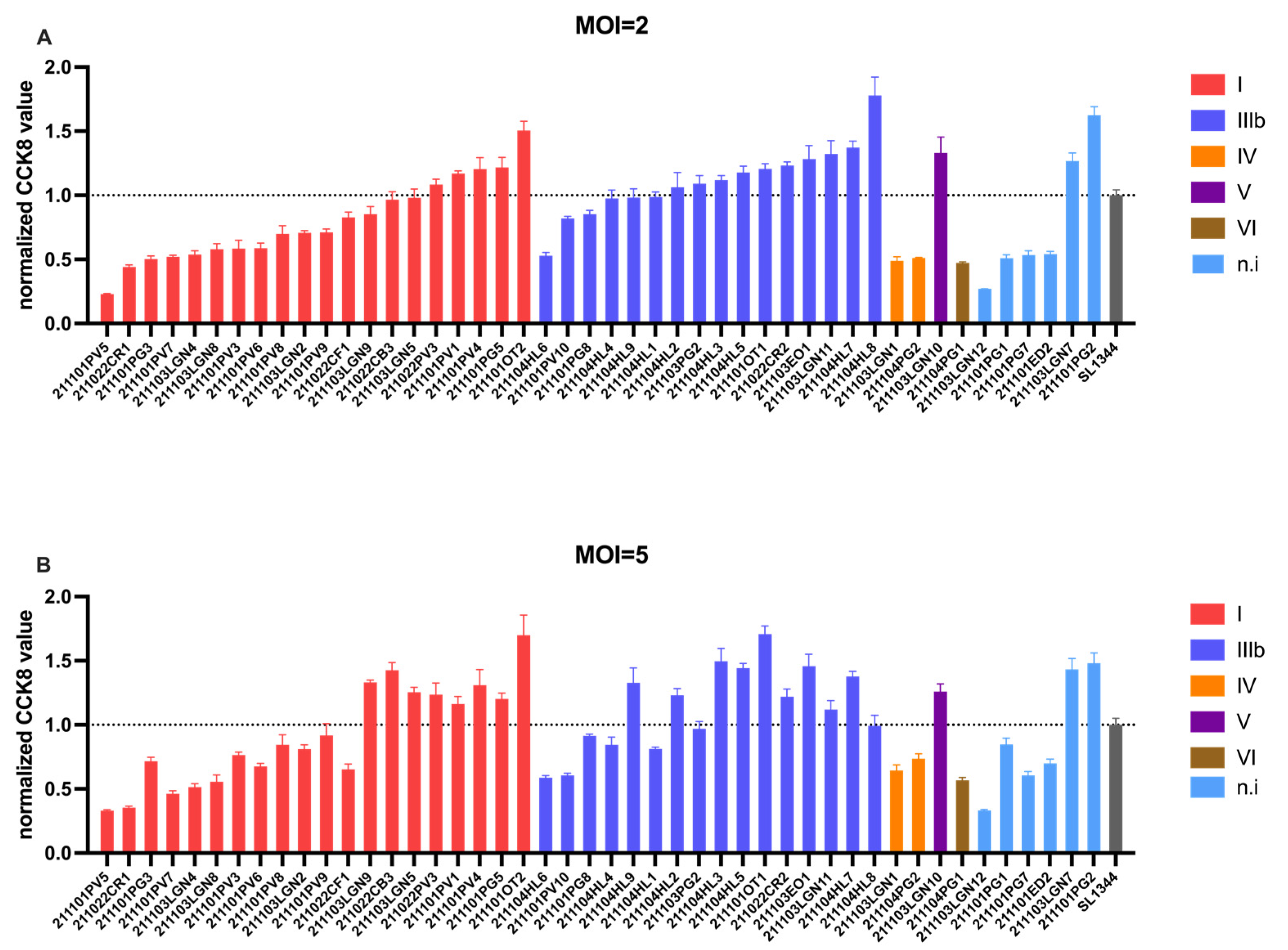
3.2. Cytotoxicity of Isolated Salmonella spp.
3.3. Serovar Information of Salmonella Isolated from Lizards via WGS
3.4. Antibiotic Resistant Profiles of Reptile Associated Salmonella Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, H.K.; Frederick, J.A.; Crump, J.A. Global burden of invasive nontyphoidal Salmonella disease 2010. Emerg. Infect Dis. 2015, 21, 941–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vazquez, B.; Franco, C.M.; Cepeaa, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef]
- Patrick, A.D.; Francois-Xavier, W. Antigenic Formulae of the Salmonella Serovars, 9th ed.; World Health Organization: Paris, France, 2007; pp. 1–166. [Google Scholar]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; Guibourdenche, M.; Pinna, E.D.; Satheesh, N.; Fields, F.I.; Weill, F. Supplement 2008–2010 (no. 48) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, G.L.; Papa, M.I. Salmonella spp. Infection—A continuous threat worldwide. Germs 2021, 11, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Greening, B.J.; Whitham, H.K.; Aldous, W.K.; Hall, N.; Garvey, A.; Mandernach, S.; Kahn, E.B.; Patrick, N.; Snow, J.; Melter, M.I.; et al. Public health response to multistate Salmonella Typhimurium outbreak associated with prepackaged chicken salad, United States, 2018. Emerg. Infect Dis. 2022, 28, 1254–1256. [Google Scholar] [CrossRef]
- Fabrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [Green Version]
- Pees, M.; Rabsch, W.; Plenz, B.; Fruth, A.; Prager, R.; Simon, S.; Schmidt, V.; Munch, S.; Braun, P. Evidence for the transmission of Salmonella from reptiles to children in Germany, July 2010 to October 2011. Eurosurveillance 2013, 18, 20634. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Glass, K.; Liu, B.; Hope, K.; Kirk, M. Salmonella infection in middle-aged and older adults: Incidence and risk factors from the 45 and up study. Foodborne Pathog. Dis. 2016, 13, 689–694. [Google Scholar] [CrossRef]
- Ebani, V.V. Domestic reptiles as source of zoonotic bacteria: A mini review. Asian Pac. J. Trop. Med. 2017, 10, 723–728. [Google Scholar] [CrossRef]
- Marin, C.; Lorenzo-Rebenaque, L.; Laso, O.; Villora-Gonzalez, J.; Vega, S. Pet reptiles: A potential source of transmission of multidrug-resistant salmonella. Front. Vet. Sci. 2020, 7, 613718. [Google Scholar] [CrossRef]
- Zhang, J.; Kuang, D.; Wang, F.; Meng, J.; Jin, H.; Yang, X.; Liao, M.; Klena, J.D.; Wu, S.; Zhang, Y.; et al. Turtles as a possible reservoir of nontyphoidal Salmonella in Shanghai, China. Foodborne Pathog. Dis. 2016, 13, 428–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, S.; Tauxe, R.V.; Behravesh, C.B. Turtle-associated salmonellosis, United States, 2006–2014. Emerg. Infect Dis. 2016, 22, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C. Overview: Foodborne Pathogens in Wildlife Populations; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–30. [Google Scholar]
- Harris, J.R.; Bergmire-Sweat, D.; Schlegel, J.H.; Winpisinger, K.A.; Klos, R.F.; Perry, C.; Tauxe, R.V.; Sotir, M.J. Multistate outbreak of Salmonella infections associated with small turtle exposure, 2007–2008. Pediatrics 2009, 124, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Marus, J.R.; Magee, M.J.; Manikonda, K.; Nichols, M.C. Outbreaks of Salmonella enterica infections linked to animal contact: Demographic and outbreak characteristics and comparison to foodborne outbreaks-United States, 2009–2014. Zoonoses Public Health 2019, 66, 370–376. [Google Scholar] [CrossRef]
- Schroter, M.; Roggentin, P.; Hofmann, J.; Speicher, L.; Laufs, R.; Mack, D. Pet snakes as a reservoir for Salmonella enterica subsp. diarizonae (Serogroup IIIb): A prospective study. Appl. Environ. Microbiol. 2004, 70, 613–615. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Wang, F.; Shi, H.; Zhou, P.; Ge, Y.; Hua, L.; Liu, W. Highly pathogenic Salmonella Pomona was first isolated from the exotic red-eared slider (Trachemys scripta elegans) in the wild in China: Implications for public health. Sci. Total Environ. 2014, 468–469, 28–30. [Google Scholar] [CrossRef]
- Bush, E.R.; Baker, S.E.; Macdonald, D.W. Global trade in exotic pets 2006–2012. Conserv. Biol. 2014, 28, 663–676. [Google Scholar] [CrossRef]
- Diaz, M.A.; Cooper, R.K.; Cloeckaert, A.; Siebeling, R.J. Plasmid-mediated high-level gentamicin resistance among enteric bacteria isolated from pet turtles in Louisiana. Appl. Environ. Microb. 2006, 72, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Cristina, G.; Maria, F.; Vittorio, F.; Giuseppe, L.; Aurora, A.; Caterina, M. Antibiotic resistance in Salmonella Isolated from Tegus (Tupinambis spp.). J. Exot. Pet Med. 2012, 21, 328–331. [Google Scholar]
- Walters, M.S.; Simmons, L.; Anderson, T.C.; DeMent, J.; Zile, K.V.; Matthias, L.P.; Etheridge, S.; Baker, R.; Healan, C.; Bagby, R.; et al. Outbreaks of salmonellosis from small turtles. Pediatrics 2016, 137, e20151735. [Google Scholar] [CrossRef] [Green Version]
- Bjelland, A.M.; Sandvik, L.M.; Skarstein, M.M.; Svendal, L.; Debenham, J.J. Prevalence of Salmonella serovars isolated from reptiles in Norwegian zoos. Acta Vet. Scand. 2020, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wikström, V.O.; Fernström, L.L.; Melin, L.; Boqvist, S. Salmonella isolated from individual reptiles and environmental samples from terraria in private households in Sweden. Acta Vet. Scand. 2014, 56, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, K.H.; Blitchington, R.B.; Greene, R.C. Amplification of bacterial 16S ribosomal DNA with polymerase chain reaction. J. Clin. Microbiol. 1990, 28, 1942–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Iwata, T.; Shimizu, M.; Taniguchi, T.; Nakadai, A.; Hirota, Y.; Hayashidani, H. A novel multiplex PCR assay for Salmonella subspecies identification. J. Appl. Microbiol. 2009, 107, 805–811. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of changes to the clinical and laboratory standards institute performance standards for antimicrobial susceptibility testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e00213-21. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling Genomes and Mini-metagenomes from Highly Chimeric Reads. In Proceedings of the Annual International Conference on Research in Computational Molecular Biology, Berlin, Germany, 7 April 2013; pp. 158–170. [Google Scholar]
- McWhorter, A.; Owens, J.; Valcanis, M.; Olds, L.; Myers, C.; Smith, I.; Trott, D.; McLelland, D. In vitro invasiveness and antimicrobial resistance of Salmonella enterica subspecies isolated from wild and captive reptiles. Zoonoses Public Health 2021, 68, 402–412. [Google Scholar] [CrossRef]
- Geue, L.; Loschner, U. Salmonella enterica in reptiles of German and Austrian origin. Vet. Microbiol. 2002, 84, 79–91. [Google Scholar] [CrossRef]
- Nakadai, A.; Kuroki, T.; Kato, Y.; Suzuki, R.; Yamai, S.; Yaginuma, C.; Shiotani, R.; Yamanouchi, A.; Hayashidani, H. Prevalence of Salmonella spp. in pet reptiles in Japan. J. Vet. Med. Sci. 2005, 67, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.P.; Varriale, L.; Borrelli, L.; Pace, A.; Latronico, M.; Menna, L.F.; Fioretti, A.; Dipineto, L. Salmonella serotypes isolated in geckos kept in seven collections in southern Italy. J. Small Anim. Pract. 2018, 59, 294–297. [Google Scholar] [CrossRef]
- Kikillus, K.H.; Gartrell, B.D.; Motion, E. Prevalence of Salmonella spp., and serovars isolated from captive exotic reptiles in New Zealand. New Zealand Vet. J. 2011, 59, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Lukac, M.; Pedersen, K.; Prukner-Radovcic, E. Prevalence of Salmonella in captive reptiles from Croatia. J. Zoo Wildl. Med. 2015, 46, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Kai, W.; Jinlong, R.; Hongman, C.; Che, J. The updated checklists of amphibians and reptiles of China. Biodiv. Sci. 2020, 28, 189–218. [Google Scholar] [CrossRef]
- Scheelings, T.F.; Lightfoot, D.; Holz, P. Prevalence of Salmonella in Australian reptiles. J. Wildlife Dis. 2011, 47, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hydeskov, H.B.; Guardabassi, L.; Aalbaek, B.; Olsen, K.; Nielsen, S.S.; Bertelsen, M.F. Salmonella prevalence among reptiles in a zoo education setting. Zoonoses Public Health 2013, 60, 291–295. [Google Scholar] [CrossRef]
- Corrente, M.; Sangiorgio, G.; Grandolfo, E.; Bodnar, L.; Catella, C.; Trotta, A.; Martella, V.; Buonavoglia, D. Risk for zoonotic Salmonella transmission from pet reptiles: A survey on knowledge, attitudes and practices of reptile-owners related to reptile husbandry. Prev. Vet. Med. 2017, 146, 73–78. [Google Scholar] [CrossRef]
- Galán, J.E.; Ginocchio, C.; Costeas, P. Molecular and functional characterization of the Salmonella invasion gene invA: Homology of InvA to members of a new protein family. J. Bacteriol. 1992, 174, 4338–4349. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.M.; Lin, W.; Van, K.T.; Phan, L.; Tran, N.N.; Farmer, D. Rapid detection of Salmonella in foods using real-time PCR. J. Food Prot. 2008, 71, 2436–2441. [Google Scholar] [CrossRef]
- Kadry, M.; Nader, S.M.; Dorgham, S.M.; Kandil, M.M. Molecular diversity of the invA gene obtained from human and egg samples. Vet. World 2019, 12, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Yanestria, S.M.; Rahmaniar, R.P.; Wibisono, F.J.; Effendi, M.H. Detection of invA gene of Salmonella from milkfish (Chanos chanos) at Sidoarjo wet fish market, Indonesia, using polymerase chain reaction technique. Vet. World 2019, 12, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Wray, C.; Sojka, W.J. Experimental Salmonella typhimurium infection in calves. Res. Vet. Sci. 1978, 25, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, W.R.; Mcneil, E.; Taylor, T.J. Avian salmonellosis: Types of Salmonella Isolated and their relation to public health. Am. J. Epidemiol. 1944, 3, 264–278. [Google Scholar] [CrossRef]
- Franco, A.; Hendriksen, R.S.; Lorenzetti, S.; Onorati, R.; Gentile, G.; Dell‘Omo, G.; Aarestrup, F.M.; Battisti, A. Characterization of Salmonella occurring at high prevalence in a population of the land iguana Conolophus subcristatus in Galapagos Islands, Ecuador. PLoS ONE 2011, 6, 0023147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasmans, F.; Martel, A.; Boyen, F.; Vandekerchove, D.; Wybo, I.; Immerseel, F.V.; Heyndrickx, M.; Collard, J.M.; Ducatelle, R.; Haesebrouk, F. Characterization of Salmonella isolates from captive lizards. Vet. Microbiol. 2005, 110, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Romero, S.B.; Kvapil, P.; Cizek, A.; Knotek, Z. The incidence and antimicrobial susceptibility of Salmonella species isolated from captive reptiles at Ljubljana Zoo. Slov. Vet. Res. 2016, 1, 39–43. [Google Scholar]
- Wang, X.; Wang, H.; Li, T.; Liu, F.; Cheng, Y.; Guo, X.; Wen, G.; Luo, Q.; Shao, H.; Pan, Z.; et al. Characterization of Salmonella spp. isolated from chickens in central China. BMC Vet. Res. 2020, 16, 299. [Google Scholar] [CrossRef]
- Li, Y.; Yang, X.; Zhang, H.; Jia, H.; Liu, X.; Yu, B.; Zeng, Y.; Zhang, Y.; Pei, X.; Yang, D. Prevalence and antimicrobial susceptibility of Salmonella in the commercial eggs in China. Int. J. Food Microbiol. 2020, 325, 108623. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Q.; Huang, Y.L.; Zhou, H.W.; Zhang, R.; Zhu, K. Persistent carbapenem-resistant Klebsiella pneumoniae: A Trojan horse. Lancet Infect. Dis. 2018, 18, 22–23. [Google Scholar] [CrossRef] [Green Version]
- Vitale, A.; Pessi, G.; Urfer, M.; Locher, H.H.; Zerbe, K.; Obrecht, D.; Robinson, J.A.; Eberl, L. Identification of Genes Required for Resistance to Peptidomimetic Antibiotics by Transposon Sequencing. Front. Microbiol. 2020, 11, 1681–1694. [Google Scholar] [CrossRef]


{kind=link}
| Type | Species | Common Name | No. Positive/No. Tested (%) |
|---|---|---|---|
| Turtle | Cuora amboinensis | Amboina Box Turtle | 0/1 |
| Cuora aurocapitata | Yellow-headed Box Turtle | 0/1 | |
| Cuora flavomarginata | Snake-eating Turtle | 1/19 | |
| Cuora galbinifrons | Indochinese Box Turtle | 0/1 | |
| Cuora mccordi | McCord’s Box Turtle | 0/2 | |
| Cuora mouhotii | Keeled Box Turtle | 0/1 | |
| Cuora pani | Pan’s Box Turtle | 0/1 | |
| Cuora trifasciata | Chinese Three-striped Box Turtle | 0/2 | |
| Cuora yunnanensis | Yunnan Box Turtle | 0/1 | |
| Mauremys japonica | Japanese Pond Turtle | 0/1 | |
| Mauremys reevesii | Reeves’ Turtle | 3/20 | |
| Mauremys sinensis | Chinese Striped-necked Turtle | 0/1 | |
| Trachemys scripta elegans | Red-eared Slider | 0/1 | |
| Total | 4/52 (7.69) | ||
| Snake | Elaphe dione | Steppes Ratsnakes | 1/5 |
| Elaphe taeniura | Beauty Snake | 2/5 | |
| Heterodon nasicus | Hognose Snake | 9/15 | |
| Gonyosoma frenatum | Khasi Hills Trinket snake | 0/1 | |
| Pantherophis guttatus | Corn Snake | 9/33 | |
| Pantherophis obsoletus | Rat Snake | 1/3 | |
| Lampropeltis getula | Kingsnake | 10/41 | |
| Lycodon rufozonatus | Red-banded Snake | 0/1 | |
| Total | 32/104 (30.77) | ||
| Lizard | Pogona vitticeps | Bearded Dragon | 10/19 |
| Rhacodactylus leachianus | New Caledonia Giant Gecko | 0/2 | |
| Total | 10/21 (47.62) | ||
| Total | 46/177 (25.99) |
| Multiplex PCR Result | Subspecies | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain ID | Collecton Sites | Reptile Source | fljb | mdcA | gatD | stn | STM4057 | invA | |
| 1101ED2 | A (Individual breeder) | Elaphe dione | + | n.i | |||||
| 1101OT1 | A (Individual breeder) | Elaphe taeniura | + | + | + | IIIb | |||
| 1101OT2 | A (Individual breeder) | Elaphe taeniura | + | + | + | I | |||
| 1101PG1 | A (Individual breeder) | Pantherophis guttatus | + | n.i | |||||
| 1101PG2 | A (Individual breeder) | Pantherophis guttatus | + | n.i | |||||
| 1101PG3 | A (Individual breeder) | Pantherophis guttatus | + | + | + | + | I | ||
| 1101PG5 | A (Individual breeder) | Pantherophis guttatus | + | + | + | I | |||
| 1101PG7 | A (Individual breeder) | Pantherophis guttatus | + | n.i | |||||
| 1101PG8 | A (Individual breeder) | Pantherophis guttatus | + | + | + | IIIb | |||
| 1103EO1 | B (Individual breeder) | Pantherophis obsoletus | + | + | + | IIIb | |||
| 1103LGN1 | B (Individual breeder) | Lampropeltis getula | + | IV | |||||
| 1103LGN10 | B (Individual breeder) | Lampropeltis getula | + | + | V | ||||
| 1103LGN11 | B (Individual breeder) | Lampropeltis getula | + | + | + | IIIb | |||
| 1103LGN12 | B (Individual breeder) | Lampropeltis getula | + | n.i | |||||
| 1103LGN2 | B (Individual breeder) | Lampropeltis getula | + | + | + | + | + | I | |
| 1103LGN4 | B (Individual breeder) | Lampropeltis getula | + | + | + | I | |||
| 1103LGN5 | B (Individual breeder) | Lampropeltis getula | + | + | + | I | |||
| 1103LGN7 | B (Individual breeder) | Lampropeltis getula | + | + | n.i | ||||
| 1103LGN8 | B (Individual breeder) | Lampropeltis getula | + | + | + | I | |||
| 1103LGN9 | B (Individual breeder) | Lampropeltis getula | + | + | + | + | + | I | |
| 1103PG2 | B (Individual breeder) | Pantherophis guttatus | + | + | + | + | IIIb | ||
| 1104HL1 | C (Individual breeder) | Heterodon nasicus | + | + | + | IIIb | |||
| 1104HL2 | C (Individual breeder) | Heterodon nasicus | + | + | + | + | IIIb | ||
| 1104HL3 | C (Individual breeder) | Heterodon nasicus | + | + | + | + | IIIb | ||
| 1104HL4 | C (Individual breeder) | Heterodon nasicus | + | + | + | IIIb | |||
| 1104HL5 | C (Individual breeder) | Heterodon nasicus | + | + | + | + | IIIb | ||
| 1104HL6 | C (Individual breeder) | Heterodon nasicus | + | + | + | IIIb | |||
| 1104HL7 | C (Individual breeder) | Heterodon nasicus | + | + | + | + | IIIb | ||
| 1104HL8 | C (Individual breeder) | Heterodon nasicus | + | + | + | + | IIIb | ||
| 1104HL9 | C (Individual breeder) | Heterodon nasicus | + | + | + | IIIb | |||
| 1104PG1 | C (Individual breeder) | Pantherophis guttatus | + | + | + | + | VI | ||
| 1104PG2 | C (Individual breeder) | Pantherophis guttatus | + | + | IV | ||||
| 1022PV3 | D (Individual breeder) | Pogona vitticeps | + | + | + | I | |||
| 1101PV1 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1101PV10 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | IIIb | ||
| 1101PV3 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | I | ||
| 1101PV4 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1101PV5 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1101PV6 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1101PV7 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1101PV8 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1101PV9 | D (Individual breeder) | Pogona vitticeps | + | + | + | + | + | I | |
| 1022CB3 | E (Commercial farm) | Mauremys reevesii | + | + | + | + | + | I | |
| 1022CF1 | E (Commercial farm) | Cuora flavomarginata | + | + | + | + | + | I | |
| 1022CR1 | E (Commercial farm) | Mauremys reevesii | + | + | + | + | + | I | |
| 1022CR2 | E (Commercial farm) | Mauremys reevesii | + | + | + | IIIb | |||
| Strain ID | Reptile Source | Subspecies | Serovar Type |
|---|---|---|---|
| 1022PV3 | Pogona vitticeps | I | ago |
| 1101PV1 | Pogona vitticeps | I | pomona |
| 1101PV10 | Pogona vitticeps | IIIb | IIIb 57:c:z |
| 1101PV3 | Pogona vitticeps | I | ago |
| 1101PV4 | Pogona vitticeps | I | pomona |
| 1101PV5 | Pogona vitticeps | I | carmel |
| 1101PV6 | Pogona vitticeps | I | carmel |
| 1101PV7 | Pogona vitticeps | I | carmel |
| 1101PV8 | Pogona vitticeps | I | carmel |
| 1101PV9 | Pogona vitticeps | I | carmel |
| Salmonella from Snakes (%) | Salmonella from Lizards (%) | Salmonella from Turtles (%) | Total (%) | |
|---|---|---|---|---|
| Beta-lactam antibiotic | ||||
| Ampicillin | 25.00 | 30.00 | 0 | 25.53 |
| Meropenem | 0 | 0 | 0 | 2.13 |
| Amoxicillin-clavulanic acid | 25.00 | 50.00 | 0 | 29.79 |
| Cephalosporin Antibiotic | ||||
| Ceftiofur | 12.50 | 10.00 | 0 | 14.89 |
| Cefazolin | 50.00 | 40.00 | 0 | 44.68 |
| Aminoglycoside antibiotic | ||||
| Gentamicin | 96.88 | 70.00 | 75.00 | 89.36 |
| Streptomycin | 9.38 | 10.00 | 25.00 | 12.77 |
| Amikacin sulfate | 0 | 0 | 0 | 2.13 |
| Kanamycin | 40.63 | 60.00 | 75.00 | 51.06 |
| Quinolone antibiotics | ||||
| Ciprofloxacin | 50 | 40.00 | 50.00 | 51.06 |
| Enrofloxacin | 18.75 | 40.00 | 25.00 | 25.53 |
| Nalidixic acid | 28.13 | 30.00 | 0 | 27.66 |
| Chloramphenicol antibiotics | ||||
| Chloramphenicol | 3.13 | 10.00 | 0 | 6.38 |
| Florfenicol | 15.63 | 40.00 | 0 | 23.40 |
| Tetracycline antibiotics | ||||
| Tetracycline | 90.63 | 100.00 | 75.00 | 91.49 |
| Polypeptide antibiotics | ||||
| Polymyxin B sulfate | 25.00 | 20.00 | 50.00 | 27.66 |
| Nitrofuran antibiotics | ||||
| Macrodantin | 100 | 90.00 | 75.00 | 97.87 |
| Sulfonamide antibiotics | ||||
| Bactrim | 15.63 | 40.00 | 75.00 | 27.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, D.; He, X.; Chi, Y.; Zhang, Z.; Shuai, J.; Wang, H.; Li, Q.; Du, M. Cytotoxicity and Antimicrobial Resistance of Salmonella enterica Subspecies Isolated from Raised Reptiles in Beijing, China. Animals 2023, 13, 315. https://doi.org/10.3390/ani13020315
Song D, He X, Chi Y, Zhang Z, Shuai J, Wang H, Li Q, Du M. Cytotoxicity and Antimicrobial Resistance of Salmonella enterica Subspecies Isolated from Raised Reptiles in Beijing, China. Animals. 2023; 13(2):315. https://doi.org/10.3390/ani13020315
Chicago/Turabian StyleSong, Dingka, Xuebai He, Yiming Chi, Zhao Zhang, Jing Shuai, Hui Wang, Qiuming Li, and Mengze Du. 2023. "Cytotoxicity and Antimicrobial Resistance of Salmonella enterica Subspecies Isolated from Raised Reptiles in Beijing, China" Animals 13, no. 2: 315. https://doi.org/10.3390/ani13020315
APA StyleSong, D., He, X., Chi, Y., Zhang, Z., Shuai, J., Wang, H., Li, Q., & Du, M. (2023). Cytotoxicity and Antimicrobial Resistance of Salmonella enterica Subspecies Isolated from Raised Reptiles in Beijing, China. Animals, 13(2), 315. https://doi.org/10.3390/ani13020315