
Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Sample Collection
2.2. Bisulfite Sequencing Polymerase Chain Reaction
2.3. Real-Time Quantitative PCR
2.4. Plasmid Construction
2.5. Cell Culture and Transfection
2.6. Western Blot
2.7. Cell Counting Kit-8 (CCK-8) Assay to Determine Cell Proliferation
2.8. Flow Cytometry to Determine the Cell Cycle
2.9. Statistical Analysis
3. Results
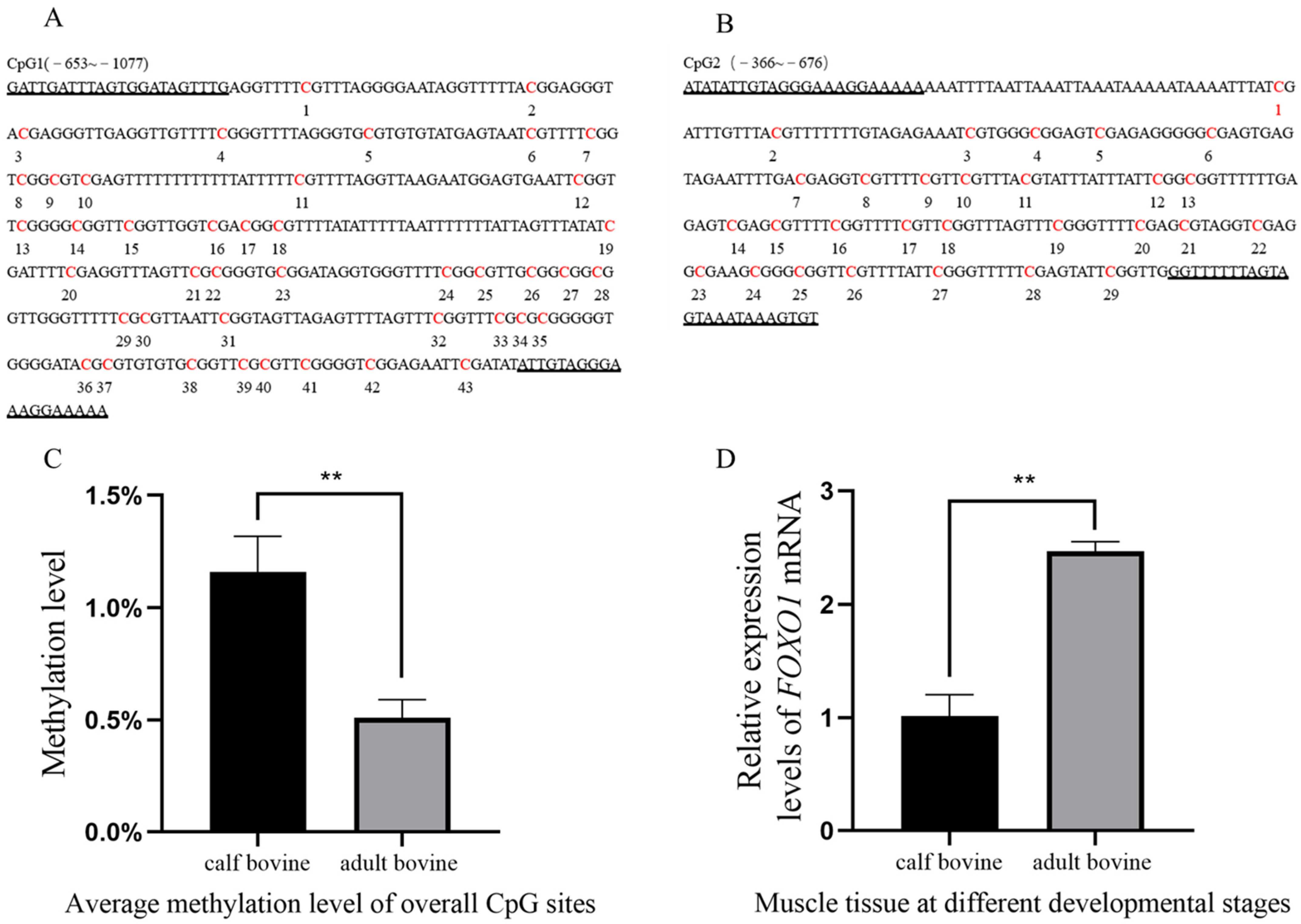
3.1. Effect of FOXO1 Promoter Methylation on Transcript Levels
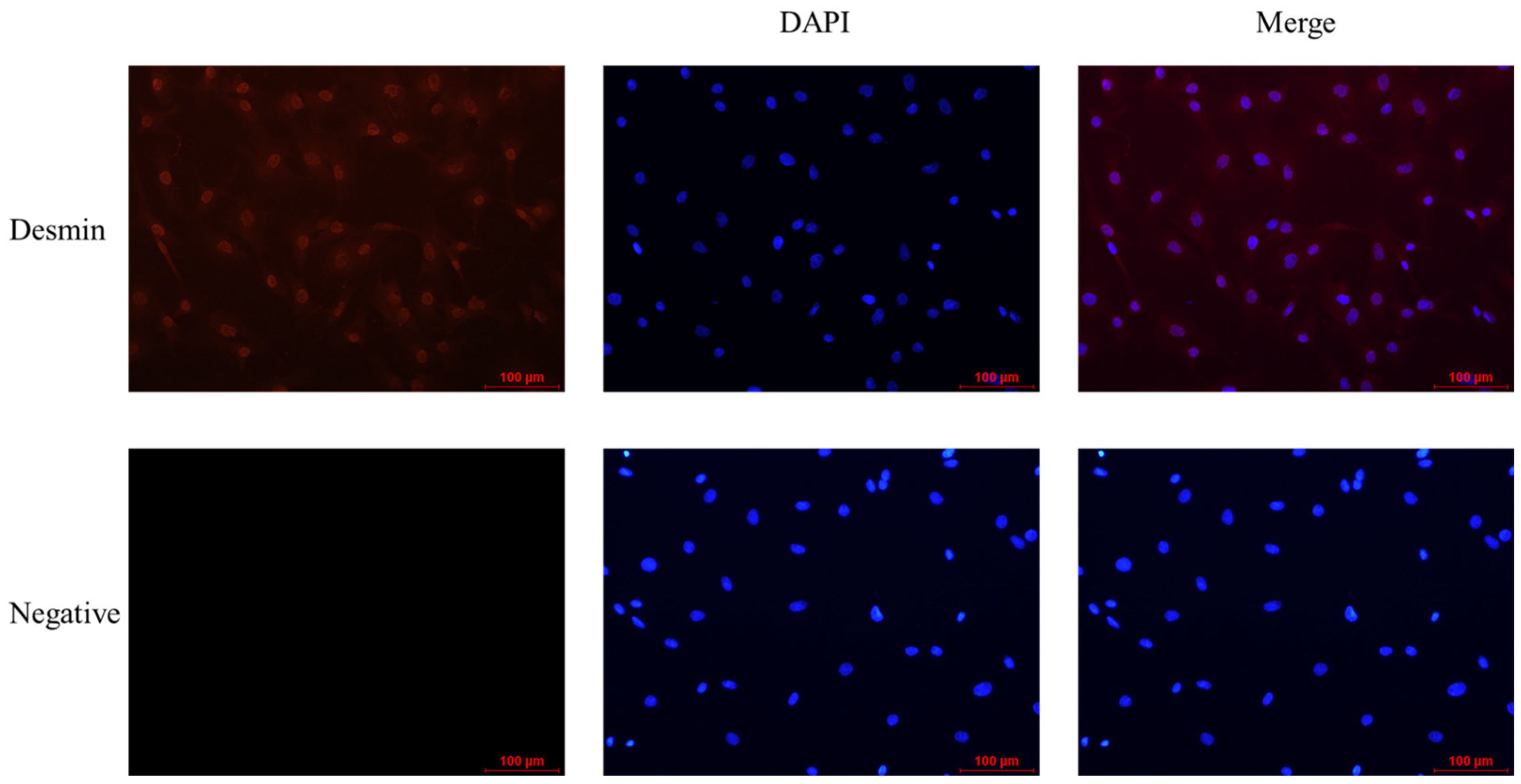
3.2. Cell Purity Assay
3.3. Assay of shRNA Interference Efficiency
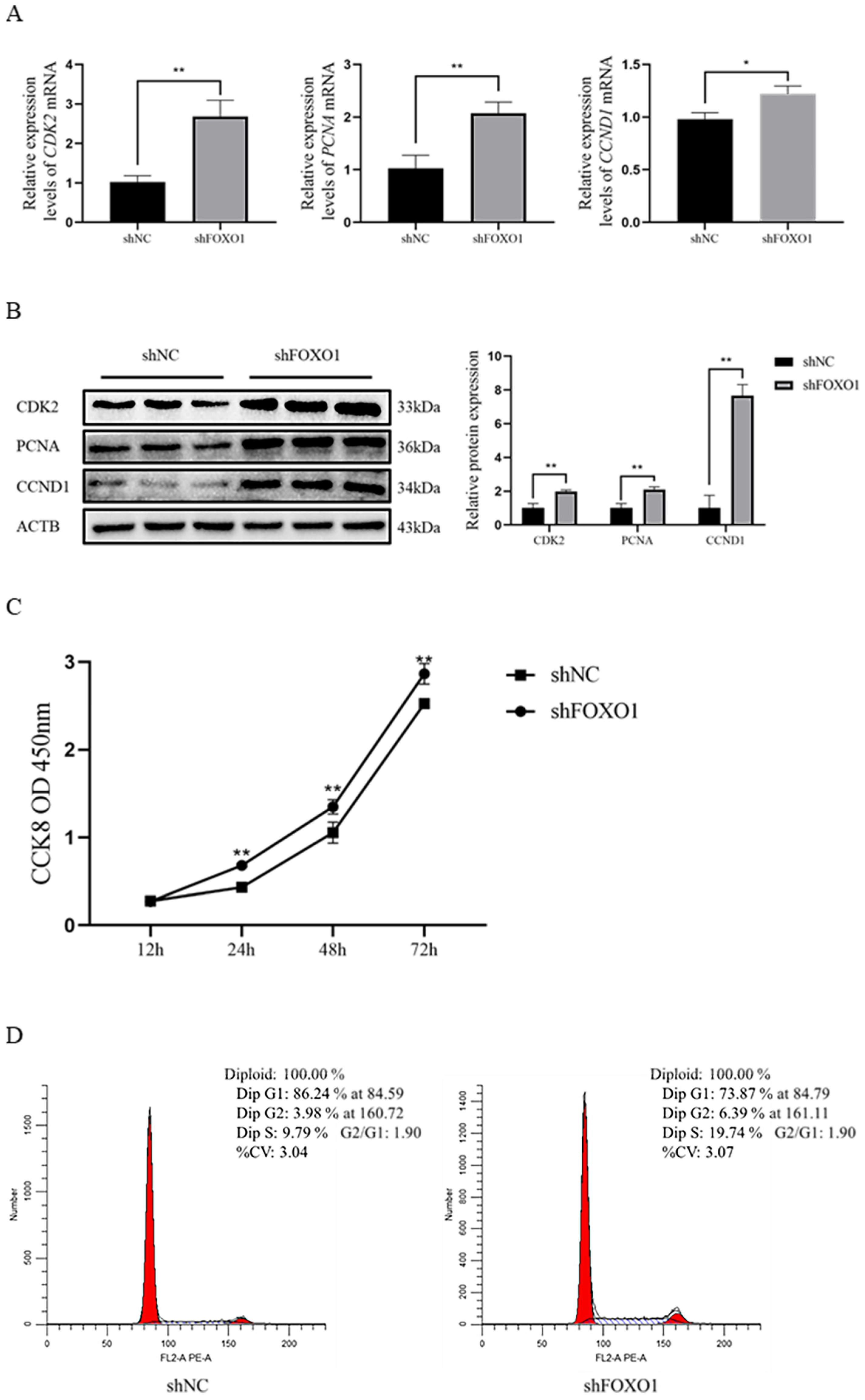
3.4. Effect of Silencing FOXO1 Gene Expression on Myoblast Proliferation
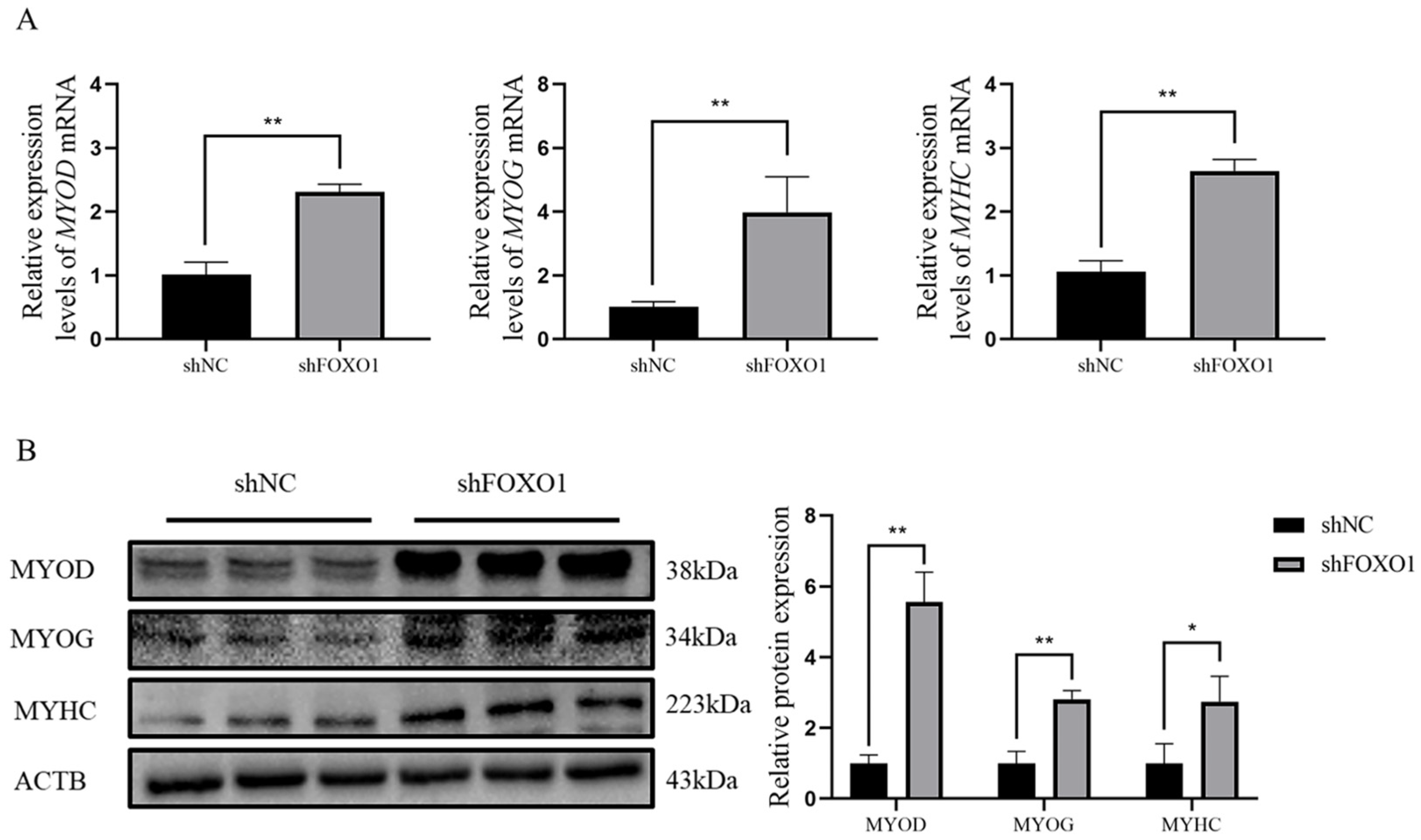
3.5. Effect of Silencing FOXO1 Gene Expression on Myoblast Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Niu, Z.; Liu, H.; Zhou, M.; Wang, H.; Liu, Y.; Li, X.; Xiong, W.; Ma, J.; Li, X.; Li, G. Knockdown of c-Myc inhibits cell proliferation by negatively regulating the Cdk/Rb/E2F pathway in nasopharyngeal carcinoma cells. Acta Biochim. Biophys. Sin. 2015, 47, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, K.; Wang, Y.; Hua, H.; Qin, S.; Yang, A.; Shao, J. Ginsenoside Rg3 Combined with Oxaliplatin Inhibits the Proliferation and Promotes Apoptosis of Hepatocellular Carcinoma Cells via Downregulating PCNA and Cyclin D1. Biol. Pharm. Bull. 2019, 42, 900–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Michel, S. The FOXO’s Advantages of Being a Family: Considerations on Function and Evolution. Cells 2020, 9, 787. [Google Scholar]
- Xing, Y.-Q.; Li, A.; Yang, Y.; Li, X.-X.; Zhang, L.-N.; Guo, H.-C. The regulation of FOXO1 and its role in disease progression. Life Sci. 2018, 193, 124–131. [Google Scholar] [CrossRef]
- Hornsveld, M.; Dansen, T.B.; Derksen, P.W.; Burgering, B.M.T. Re-evaluating the role of FOXOs in cancer. Semin. Cancer Biol. 2018, 50, 90–100. [Google Scholar] [CrossRef]
- Furuyama, T.; Kitayama, K.; Shimoda, Y.; Ogawa, M.; Sone, K.; Yoshida-Araki, K.; Hisatsune, H.; Nishikawa, S.; Nakayama, K.; Nakayama, K.; et al. Abnormal angiogenesis in Foxo1 (Fkhr)-deficient mice. J. Biol. Chem. 2004, 279, 34741–34749. [Google Scholar] [CrossRef] [Green Version]
- Hosaka, T.; Biggs, W.H.; Tieu, D.; Boyer, A.D.; Varki, N.M.; Cavenee, W.K.; Arden, K.C. Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc. Natl. Acad. Sci. USA. 2004, 101, 2975–2980. [Google Scholar] [CrossRef] [Green Version]
- Gross, D.N.; Wan, M.; Birnbaum, M.J. The role of FOXO in the regulation of metabolism. Curr. Diabetes Rep. 2009, 9, 208–214. [Google Scholar] [CrossRef]
- Chen, J.; Lu, Y.; Tian, M.; Huang, Q. Molecular mechanisms of FOXO1 in adipocyte differentiation. J. Mol. Endocrinol. 2019, 62, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Guo, X.; Jia, K.; Jing, J.; Dang, W.; Li, Y.; Qin, X.; Li, P.; Ren, Y.; Liu, W.; et al. Effects of FOXO1 on the proliferation and cell cycle-, apoptosis- and steroidogenesis-related genes expression in sheep granulosa cells. Anim. Reprod. Sci. 2020, 221, 106604. [Google Scholar] [CrossRef]
- Daniele, L.B.; Katia, A.; Maria, R.C. FoxO1 at the nexus between fat catabolism and longevity pathways. Biochim. Biophys. Acta 2014, 1841, 1555–1560. [Google Scholar]
- Ana, O.; Alejo, E.; Elena, L.; Maribel, M.; Gonzalo, G.; Marta, C.; Francisca, M.; Joaquin, P.; Sonia, M.; Eduardo, R.; et al. Pten Positively Regulates Brown Adipose Function, Energy Expenditure, and Longevity. Cell Metab. 2012, 15, 382–394. [Google Scholar]
- Wu, Y.J.; Fang, Y.H.; Chi, H.C.; Chang, L.C.; Chung, S.Y.; Huang, W.C.; Wang, X.W.; Lee, K.W.; Chen, S.L. Insulin and LiCl synergistically rescue myogenic differentiation of FoxO1 over-expressed myoblasts. PLoS ONE 2014, 9, 88450. [Google Scholar] [CrossRef] [Green Version]
- Laura, I.; Fabio, C.; Maria, R.C. Adipose Tissue and FoxO1: Bridging Physiology and Mechanisms. Cells 2020, 9, 849. [Google Scholar]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Huang, Z. FoxO1: A novel insight into its molecular mechanisms in the regulation of skeletal muscle differentiation and fiber type specification. Oncotarget 2017, 8, 10662–10674. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, K.; Huang, Y.; Lan, X.; Chen, H. Differential expression of FOXO1 during development and myoblast differentiation of Qinchuan cattle and its association analysis with growth traits. Sci. China Life Sci. 2018, 61, 826–835. [Google Scholar] [CrossRef]
- Sun, Y.; Xue, J.; Guo, W.; Li, M.; Huang, Y.; Lan, X.; Lei, C.; Zhang, C.; Chen, H. Haplotypes of bovine FoxO1 gene sequence variants and association with growth traits in Qinchuan cattle. J. Genet. 2013, 92, 8–14. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Y. Reversing DNA methylation: Mechanisms, genomics, and biological functions. Cell 2014, 156, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Kazemier, H.G.; de Groote, M.L.; Ruiters, M.H.J.; Xu, G.; Rots, M.G. Induced DNA demethylation by targeting Ten-Eleven Translocation 2 to the human ICAM-1 promoter. Nucleic Acids Res. 2014, 42, 1563–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Cao, X.K.; Peng, S.J.; Wang, X.G.; Li, Z.; Elnour, I.E.; Huang, Y.Z.; Lan, X.Y.; Chen, H. Transcriptional regulation of the bovine FGFR1 gene facilitates myoblast proliferation under hypomethylation of the promoter. J. Cell. Physiol. 2020, 235, 8667–8678. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Su, X.; Tian, Y.; Song, G.; Zan, L.; Wang, H. Effect of Actin Alpha Cardiac Muscle 1 on the Proliferation and Differentiation of Bovine Myoblasts and Preadipocytes. Animals 2021, 11, 3468. [Google Scholar] [CrossRef] [PubMed]
- Rando, T.A.; Blau, H.M. Primary mouse myoblast purification, characterization, and transplantation for cell-mediated gene therapy. J. Cell Biol. 1994, 125, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Takahashi, H.; Kitagawa, E.; Watanabe, H.; Sakurada, T.; Aso, H.; Yamaguchi, T. AMPK activation by AICAR inhibits myogenic differentiation and myostatin expression in cattle. Cell Tissue Res. 2012, 349, 615–623. [Google Scholar] [CrossRef]
- Margaret, B. Skeletal muscle formation in vertebrates. Curr. Opin. Genet. Dev. 2001, 11, 440–448. [Google Scholar]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Pipes, G.C.T.; Creemers, E.E.; Olson, E.N. The myocardin family of transcriptional coactivators: Versatile regulators of cell growth, migration, and myogenesis. Genes Dev. 2006, 20, 1545–1556. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.J.; Olson, E.N. MEF2: A central regulator of diverse developmental programs. Development 2007, 134, 4131–4140. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wang, L. Cell-Free DNA Methylation Profiling Analysis-Technologies and Bioinformatics. Cancers 2019, 11, 1741. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Hatazawa, Y.; Hirose, Y.; Ono, Y.; Kamei, Y. FOXO1 delays skeletal muscle regeneration and suppresses myoblast proliferation. Biosci. Biotechnol. Biochem. 2016, 80, 1531–1535. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef]
- Maryam, A.; Maryam, K.; Khalil, A.; Nazanin, B.; Sadra, S.T.; Mahmood, M.; Ansar, K. Multiple functions of p27 in cell cycle, apoptosis, epigenetic modification and transcriptional regulation for the control of cell growth: A double-edged sword protein. DNA Repair 2018, 69, 63–72. [Google Scholar]
- Nikolaos, Z.; Vasilios, P.; Dimitrios, D.; Konstantinos, S.; Christos, V.; Despina, N.P.; Konstantinos, K. A potential role of cyclin-dependent kinase inhibitor 1 (p21/WAF1) in the pathogenesis of endometriosis: Directions for future research. Med. Hypotheses 2019, 133, 109414. [Google Scholar]
- Li, Z.; Zhang, L.; Wang, H.; Ma, J.; Sun, J. Curcumin inhibits lung cancer progression and metastasis through induction of FOXO1. Tumour Biol. 2014, 35, 111–116. [Google Scholar] [CrossRef]
- Kamei, Y.; Miura, S.; Suzuki, M.; Kai, Y.; Mizukami, J.; Taniguchi, T.; Mochida, K.; Hata, T.; Matsuda, J.; Aburatani, H.; et al. Skeletal muscle FOXO1 (FKHR) transgenic mice have less skeletal muscle mass, down-regulated Type I (slow twitch/red muscle) fiber genes, and impaired glycemic control. J. Biol. Chem. 2004, 279, 41114–41123. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Chen, K.; Abdel, K.W.; Ward, J.L.; Yang, H.; Chabi, B.; Wrutniak-Cabello, C.; Tong, Q. Regulation of skeletal muscle oxidative capacity and muscle mass by SIRT3. PLoS ONE 2014, 9, 85636. [Google Scholar] [CrossRef]
- Hribal, M.L.; Nakae, J.; Kitamura, T.; Shutter, J.R.; Accili, D. Regulation of insulin-like growth factor-dependent myoblast differentiation by Foxo forkhead transcription factors. J. Cell Biol. 2003, 162, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.M.; Yang, Z.; Liu, C.W.; Wang, R.; Tien, P.; Dale, R.; Sun, L.Q. Effect of RNA oligonucleotide targeting Foxo-1 on muscle growth in normal and cancer cachexia mice. Cancer Gene Ther. 2007, 14, 945–952. [Google Scholar] [CrossRef]
- Bois, P.R.; Grosveld, G.C. FKHR (FOXO1a) is required for myotube fusion of primary mouse myoblasts. EMBO J. 2003, 22, 1147–1157. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | Accession Numbers | Amplified DNA Fragment (bp) |
|---|---|---|---|
| Primers used in real-time PCR | |||
| FOXO1 | F: GCAGATTTACGAGTGGATGGTC R: GCAGGGACAGATTATGACGAA | XM_025000053.1 | 107 |
| PCNA | F: ACATCAGCTCAAGTGGCGTGAAC R: GCAGCGGTAAGTGTCGAAGCC | NM_001034494.1 | 101 |
| CCND1 | F: CTGGTCCTGGTGAACAAACT R: ACAGAGGGCAACGAAGGT | NM_001046273.2 | 144 |
| CDK2 | F: CAAGTTGACGGGAGAAGTGGT R: CTTTATGAGCGGAAGAGGAAT | NM_001014934.1 | 247 |
| MYOD | F: GGCCGCTGTTTACTGTGGG R: CAGCCGCTGGTTTGGGTT | NM_001040478.2 | 162 |
| MYOG | F: TGGGCGTGTAAGGTGTGTAA R: TGCAGGCGCTCTATGTACTG | NM_001111325.1 | 197 |
| MYHC | F: GCCCACTTCTCCCTCATTCACT R: ACCCTTCTTCTTGCCACCTTTC | NM_174117.1 | 201 |
| β-actin | F: ATGATATTGCTGCGCTCGTGG R: TACGAGTCCTTCTGGCCCAT | NM_173979.3 | 151 |
| Primers used in methylation analysis | |||
| MF1-1 | F: GATTGATTTAGTGGATAGTTTG R: TTTTTCCTTTCCCTACAAT | Gene ID:506618 | 424 |
| MF1-2 | F: ATATATTGTAGGGAAAGGAAAA R: ACACTTTATTTACTACTAAAAAACC | 310 | |
| shRNA Name | Sequences |
|---|---|
| shRNA1 | 5′-3′ CAGTCTGTCCGAGATCAGTAA |
| shRNA2 | 5′-3′ AGCGGGCTGGAAGAATTCAAT |
| shRNA3 | 5′-3′ CAGGACAACAAATCGAGTTAT |
| shRNA4 | 5′-3′ CTGTGACATGGAGTCCATCAT |
| shNC | 5′-3′ GTTCTCCGAACGTGTCACGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, P.; Ruan, Y.; Liu, W.; Sun, J.; Xu, J.; Xu, H. Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts. Animals 2023, 13, 319. https://doi.org/10.3390/ani13020319
Shi P, Ruan Y, Liu W, Sun J, Xu J, Xu H. Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts. Animals. 2023; 13(2):319. https://doi.org/10.3390/ani13020319
Chicago/Turabian StyleShi, Pengfei, Yong Ruan, Wenjiao Liu, Jinkui Sun, Jiali Xu, and Houqiang Xu. 2023. "Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts" Animals 13, no. 2: 319. https://doi.org/10.3390/ani13020319
APA StyleShi, P., Ruan, Y., Liu, W., Sun, J., Xu, J., & Xu, H. (2023). Analysis of Promoter Methylation of the Bovine FOXO1 Gene and Its Effect on Proliferation and Differentiation of Myoblasts. Animals, 13(2), 319. https://doi.org/10.3390/ani13020319