
Application of Gradient-Dependent Optimal Interpolation in Fishery Analysis of Neon Flying Squid (Ommastrephes bartramii) in the Kuroshio–Oyashio Confluence Region


Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Fishery and Environment Data
2.2. Construction Algorithms
3. Results
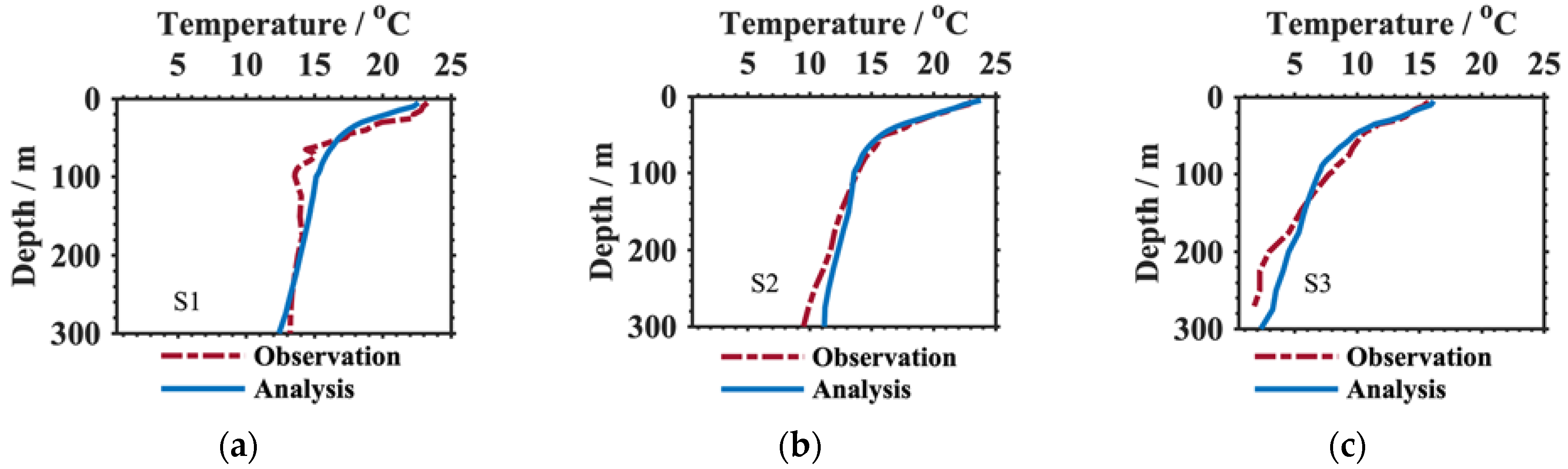
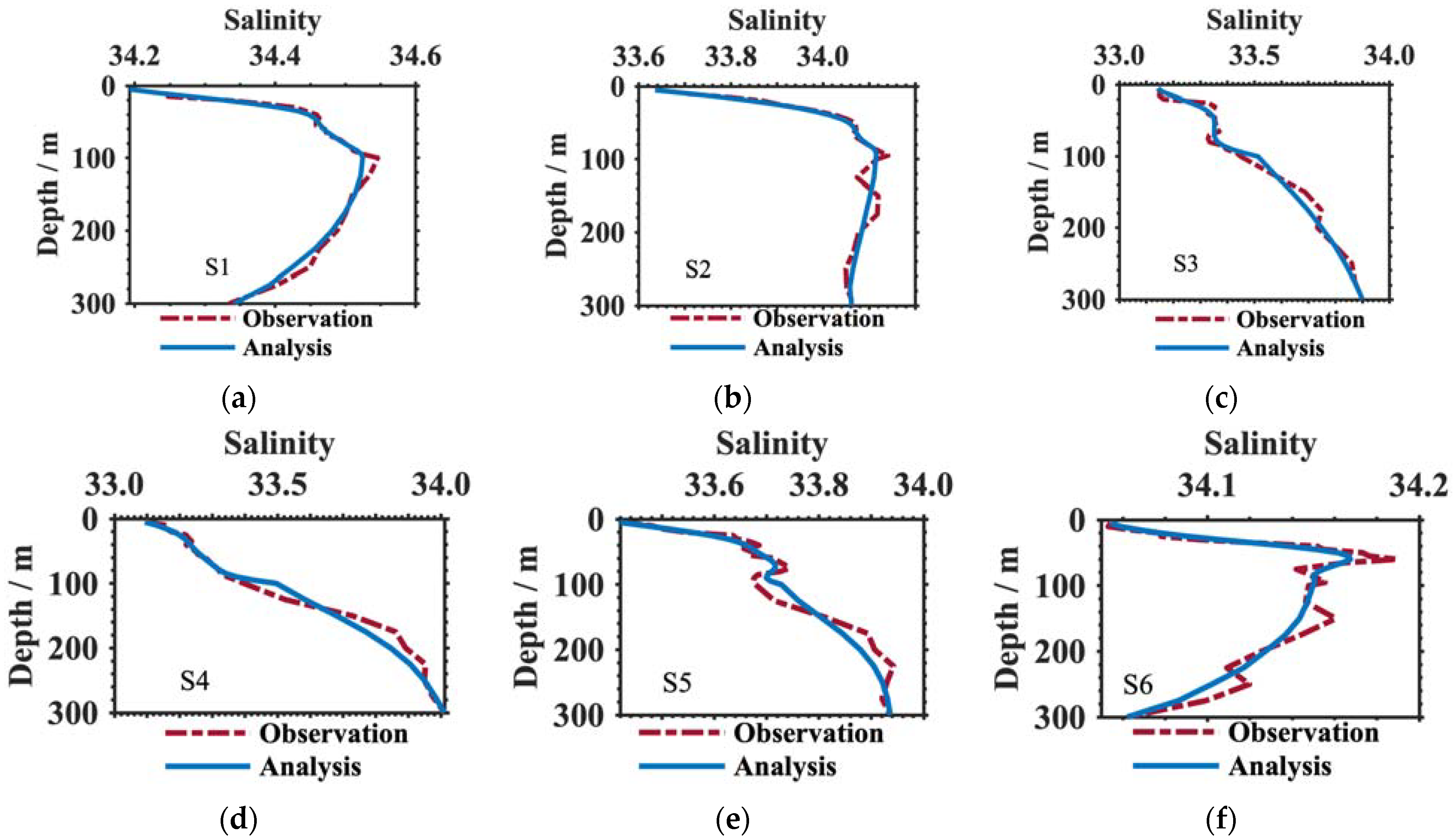
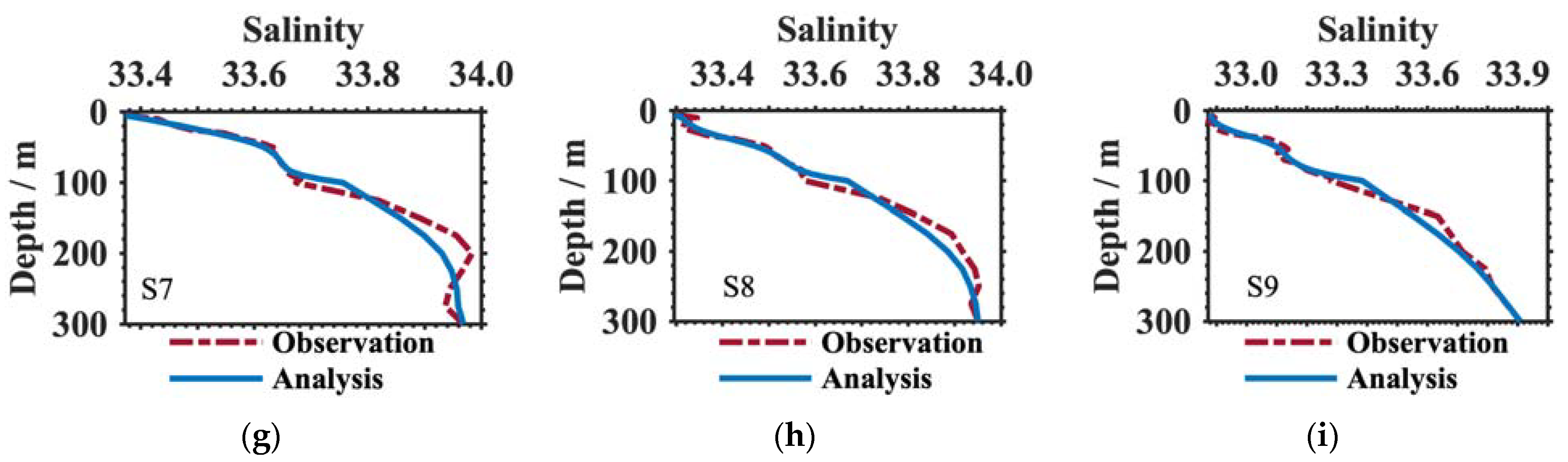
3.1. Verification of the Results
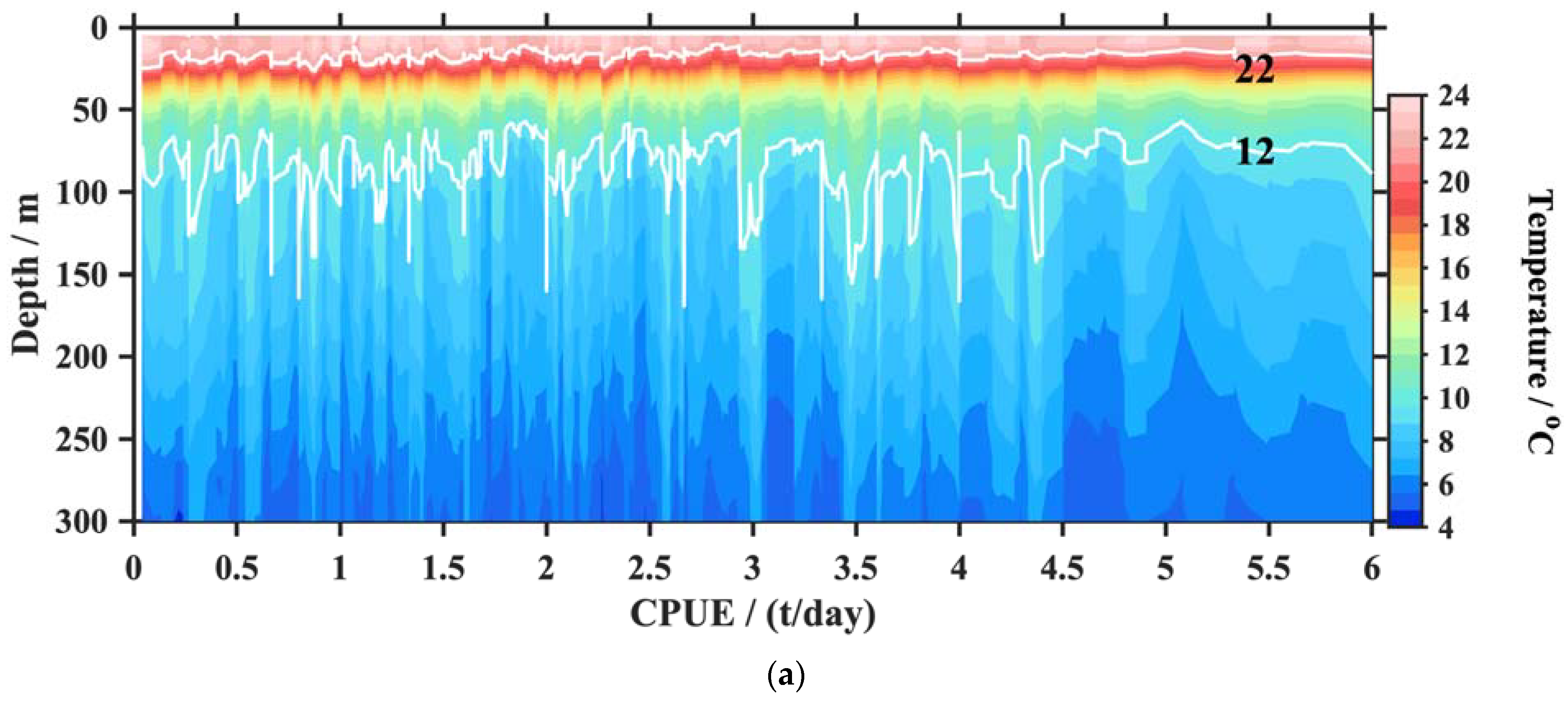
3.2. Application and Analysis
4. Discussion
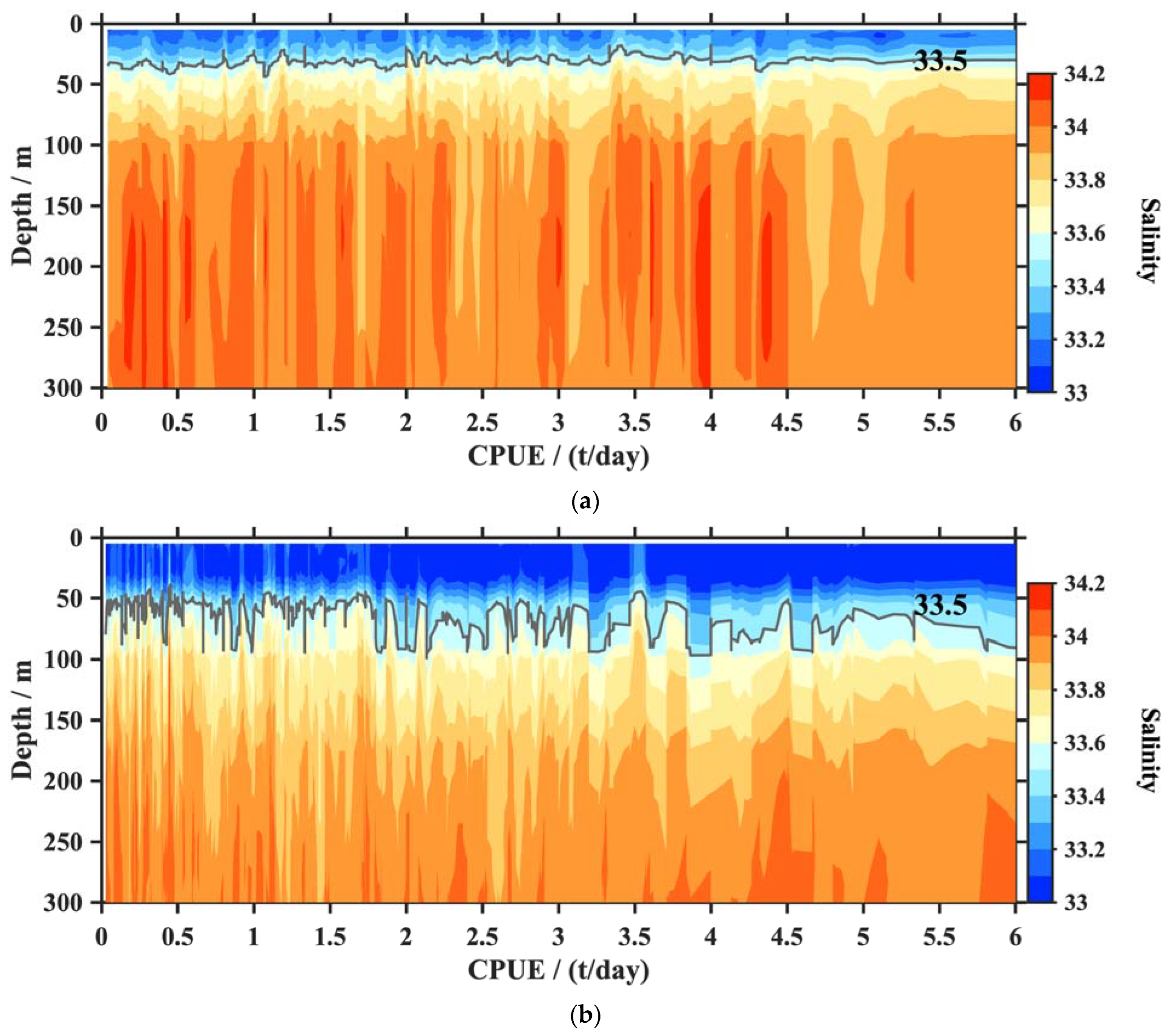
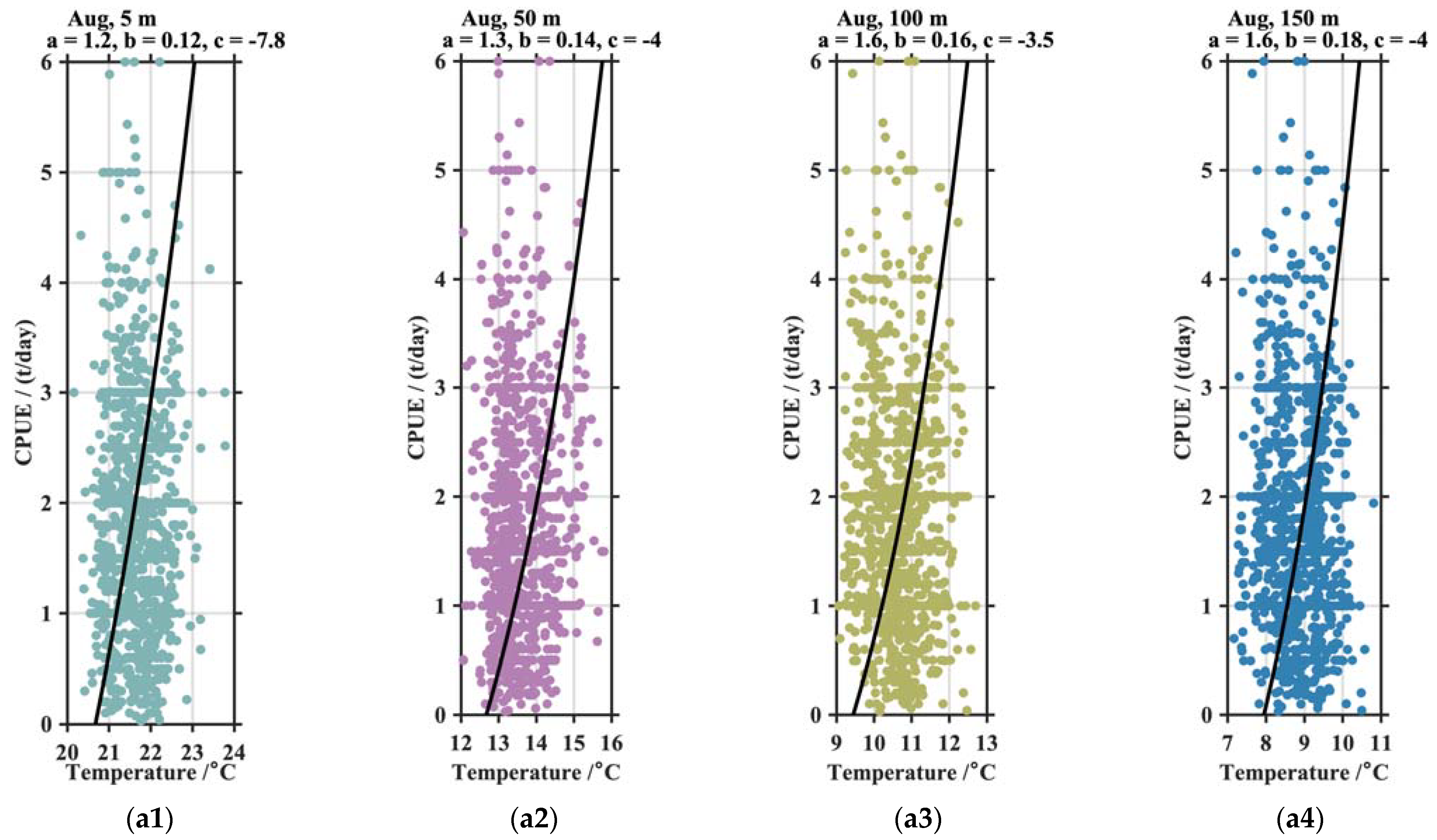
4.1. The CPUE and Vertical T/S
4.2. The CPUE and the Thermocline
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klemas, V.; Yan, X. Subsurface and deeper ocean remote sensing from satellites: An overview and new results. Prog. Oceanogr. 2014, 122, 1–9. [Google Scholar] [CrossRef]
- Langley, A.; Briand, K.; Kirby, D.S.; Murtugudde, R. Influence of oceanographic variability on recruitment of yellowfin tuna (Thunnus albacares) in the western and central Pacific Ocean. Can. J. Fish. Aquat. Sci. 2009, 66, 1462–1477. [Google Scholar] [CrossRef]
- Song, L.M.; Wu, Y.P. Standardizing CPUE of yellowfin tuna (Thunnus albacares) longline fishery in the tropical waters of the northwestern Indian Ocean using a deterministic habitat-based model. J. Oceanogr. 2011, 67, 541–550. [Google Scholar] [CrossRef]
- Yang, S.; Ma, J.; Wu, Y.; Fan, X.; Jin, S.; Chen, X. Relationship between temporal–spatial distribution of fishing grounds of bigeye tuna (Thunnus obesus) and thermocline characteristics in the Atlantic Ocean. Acta Ecol. Sin. 2015, 35, 1–9. [Google Scholar] [CrossRef]
- Liu, C. Variations of Global Ocean Salinity from Multiple Gridded Argo Products; Graduate theses and dissertations, University of South Florida: Tampa, FL, USA, 2019. [Google Scholar]
- Xu, J. Studies of Global Argo Ocean Observation; Ocean Press: Beijing, China, 2002; p. 115. [Google Scholar]
- Claustre, H.; Bishop, J.; Boss, E.; Bernard, S.; Berthon, J.F.; Coatanoan, C.; Johnson, K.; Lotiker, A.; Ulloa, O.; Perry, M.J.; et al. Bio-optical profiling floats as new observational tools for biogeochemical and ecosystem studies. In Proceedings of the OceanObs’09: Sustained Ocean Observations and Information for Society Conference, Venice, Italy, 21–25 September 2009. [Google Scholar] [CrossRef]
- Gruber, N.; Doney, S.C.; Emerson, S.R.; Gilbert, D.; Kobayashi, T.; Körtzinger, A.; Johnson, G.C.; Johnson, K.S.; Riser, S.C.; Ulloa, O. Adding oxygen to Argo: Developing a global in-situ observatory for ocean deoxygenation and biogeochemistry. In Proceedings of the OceanObs’09: Sustained Ocean Observations and Information for Society Conference, Venice, Italy, 21–25 September 2009. [Google Scholar] [CrossRef]
- Johnson, K.S. BioArgo: A global scale chemical sensor network to observe carbon, oxygen, and nitrogen cycles in the ocean. In Proceedings of the 2013 IEEE SENSORS, Baltimore, MD, USA, 3–6 November 2013; p. 1. [Google Scholar] [CrossRef]
- Johnson, K.S.; Plant, J.N.; Riser, S.C.; Gilbert, D. Air oxygen calibration of oxygen optodes on a profiling float array. J. Atmos. Ocean Technol. 2015, 32, 2160–2172. [Google Scholar] [CrossRef]
- John, R.B.; Taro, I. The red flying squid (Ommastrephes bartramii): A review of recent research and the fishery in Japan. Fish. Res. 2005, 76, 39–55. [Google Scholar] [CrossRef]
- Chen, X. Development status of world cephalopod fisheries and suggestions for squid jigging fishery in China. J. Shanghai Ocean Univ. 2019, 28, 5–14. [Google Scholar] [CrossRef]
- Kida, S.; Mitsudera, H.; Aoki, S.; Guo, X.; Ito, S.; Kobashi, F.; Komori, N.; Kubokawa, A.; Miyama, T.; Morie, R.; et al. Oceanic fronts and jets around Japan: A review. J. Oceanogr. 2015, 71, 469–497. [Google Scholar] [CrossRef]
- Nishikawa, H.; Yasuda, I.; Itoh, S. Impact of winter-to-spring environmental variability along the Kuroshio jet on the recruitment of Japanese sardine (Sardinops melanostictus). Fish. Oceanogr. 2011, 20, 570–582. [Google Scholar] [CrossRef]
- Lehodey, P.; Senina, I.; Calmettes, B.; Hampton, J.; Nicol, S. Modelling the impact of climate change on Pacific skipjack tuna population and fisheries. Clim. Chang. 2013, 119, 95–109. [Google Scholar] [CrossRef]
- Kawabata, A.; Yatsu, A.; Ueno, Y.; Suyama, S.; Kurita, Y. Spatial distribution of the Japanese common squid, Todarodespacificus, during its northward migration in the western North Pacific Ocean. Fish. Oceanogr. 2006, 15, 113–124. [Google Scholar] [CrossRef]
- Alabia, I.D.; Saitoh, S.I.; Mugo, R.; Igarashi, H.; Ishikawa, Y.; Usui, N.; Kamachi, M.; Awaji, T.; Seito, M. Seasonal potential fishing ground prediction of neon flying squid (Ommastrephes bartramii) in the western and central North Pacific. Fish. Oceanogr. 2015, 24, 190–203. [Google Scholar] [CrossRef]
- Lan, K.W.; Shimada, T.; Lee, M.A.; Su, N.J.; Chang, Y. Using remote-sensing environmental and fishery data to map potential yellowfin tuna habitats in the tropical Pacific Ocean. Remote Sens. 2017, 9, 444. [Google Scholar] [CrossRef]
- Chen, X.; Chen, Y.; Tian, S.; Liu, B.; Qian, W. An assessment of the west winter–spring cohort of neon flying squid (Ommastrephes bartramii) in the northwest Pacific Ocean. Fish. Res. 2008, 92, 221–230. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Yu, W. Variations of gravity centers of fishing ground for neon flying squid Ommastrephes bartramii in the northwest Pacific Ocean and its relation with marine environment. J. Shanghai Ocean Univ. 2020, 29, 899–909. [Google Scholar] [CrossRef]
- Tang, F.; Yang, S.; Fan, X.; Wu, Z.; Wu, Y.; Cui, X. Variation characteristics of vertical water temperature structure of neon flying squid fishery in northwestern Pacific Ocean based on Argo. J. Shanghai Ocean Univ. 2019, 28, 427–437. [Google Scholar] [CrossRef]
- Yu, W.; Chen, X.; Yi, Q.; Tian, S. A review of interaction between neon flying squid (Ommastrephes bartramii) and oceanographic variability in the north Pacific Ocean. J. Ocean Univ. China 2015, 14, 739–748. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, J.; Bao, X.; Wang, Z. An effective method for improving the accuracy of Argo objective analysis. Acta Oceanol. Sin. 2013, 32, 66–77. [Google Scholar] [CrossRef]
- Li, Z.Q.; Liu, Z.H.; Lu, S.L. Global Argo data fast receiving and post-quality-control system. IOP Conf. Ser. Earth Environ. Sci. 2020, 502, 012012. [Google Scholar] [CrossRef]
- Kalnay, E. Atmospheric Modeling, Data Assimilation and Predictability; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Murata, M.; Hayase, S. Life history and biological information on flying squid (Ommastrephes bartramii) in the north Pacific Ocean. Bull. Int. Nat. North Pac. Comm. 1993, 53, 147–182. [Google Scholar]
- Murata, M.; Nakamura, Y. Seasonal migration and diel vertical migration of the neon flying squid, Ommastrephes bartramii, in the North Pacific. In Proceedings of the International Symposium on Large Pelagic Squids, Tokyo, Japan, 18–19 July 1996; pp. 13–30. [Google Scholar]
- Chu, P.C.; Fan, C. Maximum angle method for determining mixed layer depth from seaglider data. J. Oceanogr. 2011, 67, 219–230. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | |
|---|---|---|---|---|---|---|---|---|---|
| Longitude | 162.0° E | 160.0° E | 160.0° E | 161.9° E | 162.0° E | 161.9° E | 163.9° E | 164.0° E | 164.2° E |
| Latitude | 36.0° N | 39.5° N | 44.2° N | 44.9° N | 43.0° N | 41.0° N | 43.2° N | 44.2° N | 45.1° N |
| date | 5 July 2022 | 9 July 2022 | 13 July 2022 | 14 July 2022 | 15 July 2022 | 16 July 2022 | 20 July 2022 | 21 July 2022 | 22 July 2022 |
| August | October | |||||||
|---|---|---|---|---|---|---|---|---|
| 5 m | 50 m | 150 m | 300 m | 5 m | 50 m | 150 m | 300 m | |
| <2 t | 20.6–23.8 | 12.1–15.7 | 7.6–10.8 | 4.5–7.5 | 19.8–21.7 | 11.4–17.9 | 4.0–6.8 | 4.0–5.7 |
| 2–4 t | 21.5–23.5 | 12.2–15.3 | 7.7–10.3 | 4.2–8.1 | 19.0–21.3 | 11.6–17.6 | 4.0–7.2 | 4.0–6.3 |
| 4–6 t | 22.2–23.4 | 12.4–14.6 | 7.5–9.2 | 4.9–7.3 | 18.9–20.9 | 12.1–17.0 | 4.0–5.9 | 4.0–6.1 |
| >6 t | 23.4–23.6 | 14.1–14.7 | 7.7–8.4 | 5.1–6.0 | 19.3–19.9 | 12.5–15.8 | 5.3–6.5 | 5.1–5.7 |
| August | October | |||||||
|---|---|---|---|---|---|---|---|---|
| 5 m | 50 m | 150 m | 300 m | 5 m | 50 m | 150 m | 300 m | |
| <2 t | 33.1–33.6 | 33.4–33.7 | 33.8–34.2 | 34.0–34.2 | 33.0–33.5 | 33.2–33.6 | 33.7–33.9 | 33.7–33.9 |
| 2–4 t | 33.1–33.4 | 33.6–34.0 | 33.8–34.3 | 33.2–34.2 | 33.0–33.7 | 33.4–33.8 | 33.7–33.9 | 33.7–33.9 |
| 4–6 t | 33.1–33.2 | 33.6–33.7 | 33.8–34.2 | 34.0–34. 2 | 33.1–33.2 | 33.1–33.6 | 33.6–33.7 | 34.0–34.1 |
| >6 t | 33.1–33.2 | 33.5–33.6 | 33.8–33.9 | 34.0–34.1 | 33.0–33.1 | 33.3–33.4 | 33.7–33.8 | 33.9–34.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Cui, M.; Yu, W.; Liu, B. Application of Gradient-Dependent Optimal Interpolation in Fishery Analysis of Neon Flying Squid (Ommastrephes bartramii) in the Kuroshio–Oyashio Confluence Region. Animals 2023, 13, 3425. https://doi.org/10.3390/ani13213425
Zhang C, Cui M, Yu W, Liu B. Application of Gradient-Dependent Optimal Interpolation in Fishery Analysis of Neon Flying Squid (Ommastrephes bartramii) in the Kuroshio–Oyashio Confluence Region. Animals. 2023; 13(21):3425. https://doi.org/10.3390/ani13213425
Chicago/Turabian StyleZhang, Chunling, Manman Cui, Wei Yu, and Bilin Liu. 2023. "Application of Gradient-Dependent Optimal Interpolation in Fishery Analysis of Neon Flying Squid (Ommastrephes bartramii) in the Kuroshio–Oyashio Confluence Region" Animals 13, no. 21: 3425. https://doi.org/10.3390/ani13213425
APA StyleZhang, C., Cui, M., Yu, W., & Liu, B. (2023). Application of Gradient-Dependent Optimal Interpolation in Fishery Analysis of Neon Flying Squid (Ommastrephes bartramii) in the Kuroshio–Oyashio Confluence Region. Animals, 13(21), 3425. https://doi.org/10.3390/ani13213425