

A Single-Step Genome-Wide Association Study for Semen Traits of Egyptian Buffalo Bulls

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Phenotypes
2.3. Genotyping and Quality Control
2.4. Statistical Analyses
2.5. Candidate Genes Identification
2.6. Enrichment Analyses and Functional Annotation
3. Results
3.1. Descriptive Statistics of Semen Traits in the Egyptian Buffalo Bulls
3.2. Genotype Filtering and Quality Control
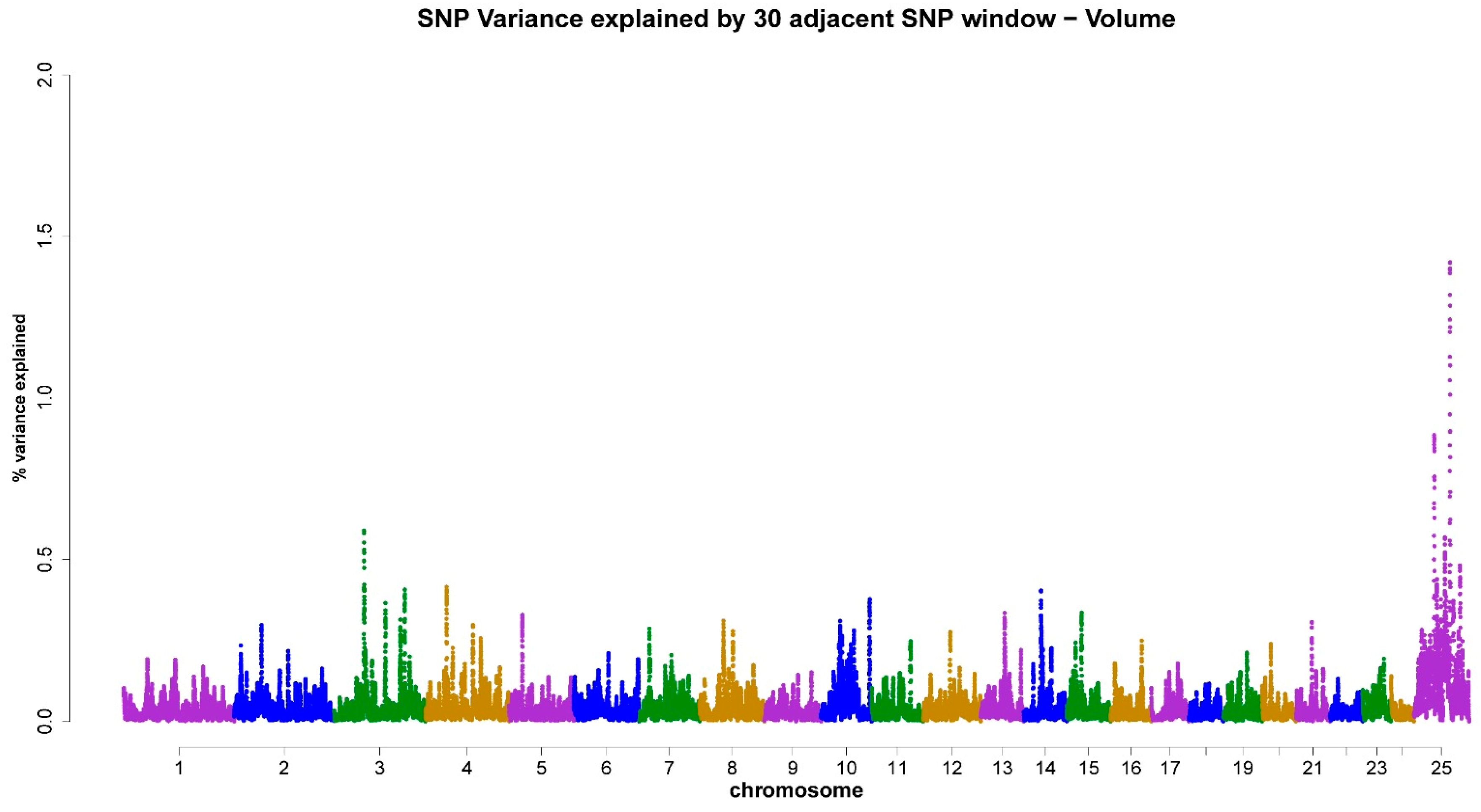
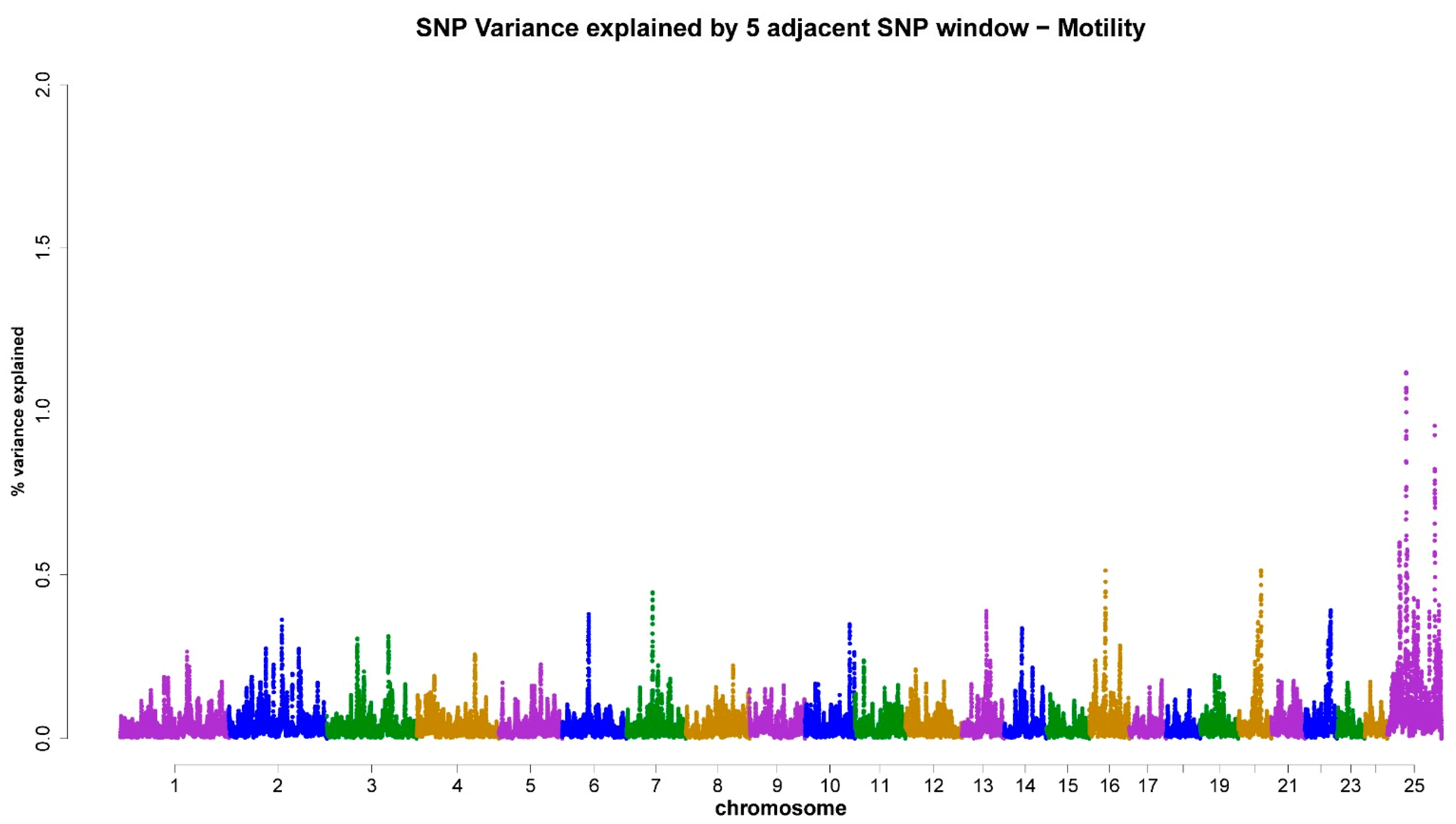
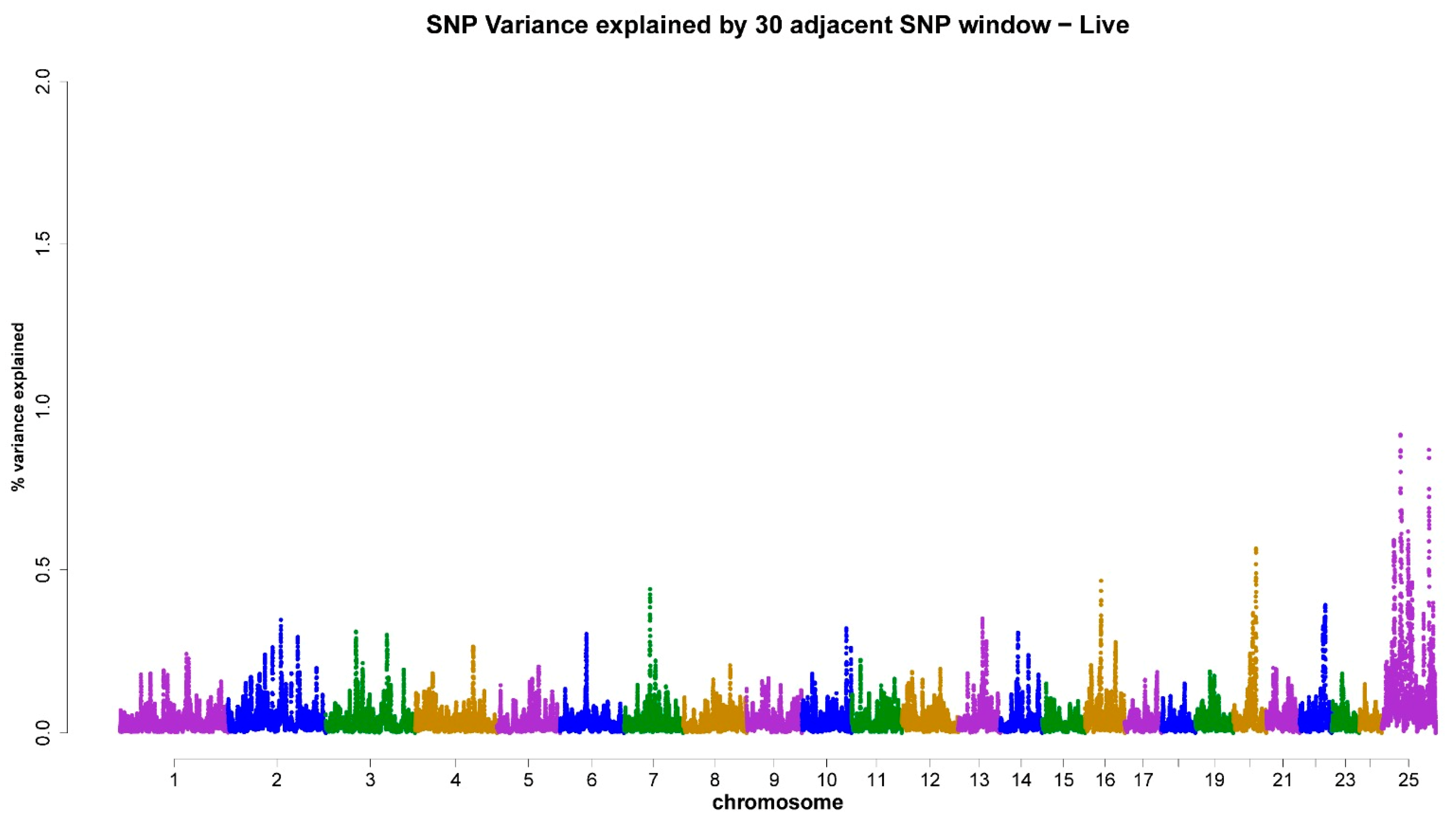
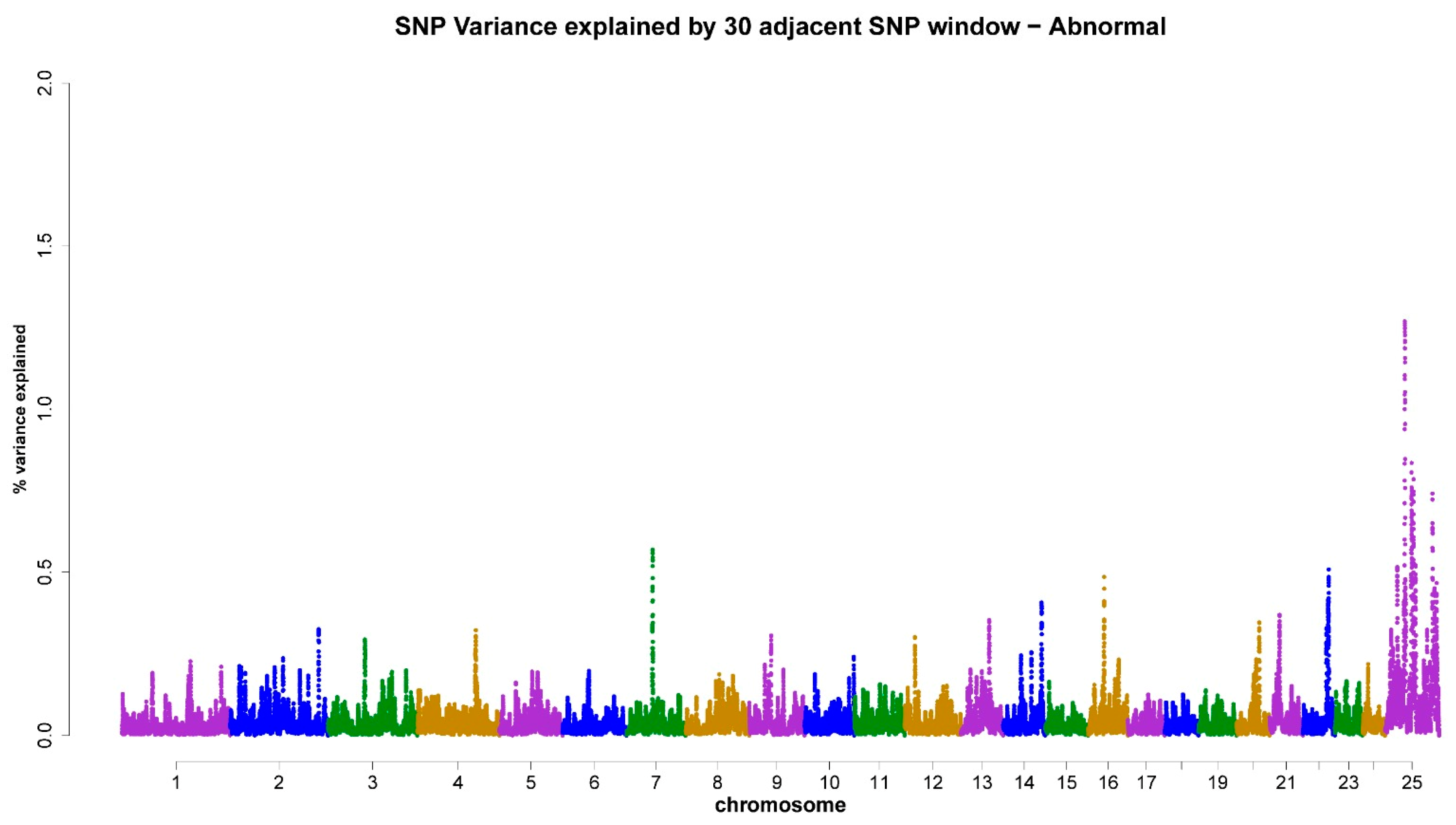
3.3. ss-GWAS for Semen Traits in the Egyptian Buffalo Bulls
3.4. Functional Annotation and Identification of Putative Candidate Genes for Semen Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Humberto, A.; Minervino, H.; Zava, M.; Vecchio, D.; Borghese, A. Bubalus Bubalis: A Short Story. Front. Vet. Sci. 2020, 7, 570413. [Google Scholar] [CrossRef]
- Vale, W.G. Effects of Environment on Buffalo Reproduction. Ital. J. Anim. Sci. 2007, 6, 130–142. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.; Awad, M.A.A.; El-Regalaty, H.; Ismael, A.; El-Assal, S.E.D.; Abou-Bakr, S. A Single-Step Genomic Evaluation for Milk Production in Egyptian Buffalo. Livest. Sci. 2020, 234. [Google Scholar] [CrossRef]
- Do, D.N.; Janss, L.L.G.; Jensen, J.; Kadarmideen, H.N. SNP Annotation-Based Whole Genomic Prediction and Selection: An Application to Feed Efficiency and Its Component Traits in Pigs. J. Anim. Sci. 2015, 93, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Fulton, J.E. Genomic Selection for Poultry Breeding. Anim. Front. 2012, 2, 30–36. [Google Scholar] [CrossRef]
- El-Halawany, N.; Abdel-Shafy, H.; Shawky, A.E.M.A.; Abdel-Latif, M.A.; Al-Tohamy, A.F.M.; Abd El-Moneim, O.M. Genome-Wide Association Study for Milk Production in Egyptian Buffalo. Livest. Sci. 2017, 198, 10–16. [Google Scholar] [CrossRef]
- Abdel-Shafy, H.; Awad, M.A.A.; El-Regalaty, H.; El-Assal, S.E.D.; Abou-Bakr, S. Prospecting Genomic Regions Associated with Milk Production Traits in Egyptian Buffalo. J. Dairy Res. 2020, 87, 389–396. [Google Scholar] [CrossRef]
- Hayes, B.J.; Bowman, P.J.; Chamberlain, A.J.; Goddard, M.E. Invited Review: Genomic Selection in Dairy Cattle: Progress and Challenges. J. Dairy Sci. 2009, 92, 433–443. [Google Scholar] [CrossRef]
- Wang, H.; Misztal, I.; Aguilar, I.; Legarra, A.; Muir, W.M. Genome-Wide Association Mapping Including Phenotypes from Relatives without Genotypes. Genet. Res. 2012, 94, 73–83. [Google Scholar] [CrossRef]
- Aguilar, I.; Misztal, I.; Johnson, D.L.; Legarra, A.; Tsuruta, S.; Lawlor, T.J. Hot Topic: A Unified Approach to Utilize Phenotypic, Full Pedigree, and Genomic Information for Genetic Evaluation of Holstein Final Score. J. Dairy Sci. 2010, 93, 743–752. [Google Scholar] [CrossRef]
- Abril-Parreño, L.; Carthy, T.R.; Keogh, K.; Štiavnická, M.; O’Meara, C.; Lonergan, P.; Kenny, D.A.; Fair, S. Genome-Wide Association Study Reveals Candidate Markers Related to Field Fertility and Semen Quality Traits in Holstein-Friesian Bulls. Animal 2023, 17, 100841. [Google Scholar] [CrossRef] [PubMed]
- Hackett, A.J.; Macpherson, J.W. A Method for Differential Staining of Bovine Spermatozoa after Extension in Sterile Milk. Can. Vet. J. 1965, 6, 117. [Google Scholar] [PubMed]
- Blom, E. Pathological Conditions in Genital Organs and Sperm as a Cause for the Rejection of Breeding Bulls for Import into and Export from Denmark (an Andrologic Retrospective, 1958–1982). Nord. Vet. Med. 1983, 35, 105–130. [Google Scholar] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. Gigascience 2015, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Luke, S.G. Evaluating Significance in Linear Mixed-Effects Models in R. Behav. Res. Methods 2017, 49, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Misztal, I.; Tsuruta, S.; Strabel, T.; Auvray, B.; Druet, T.; Lee, D.H. Blupf90 and Related Programs (Bgf90). In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002; pp. 2001–2002. [Google Scholar]
- VanRaden, P.M. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, J.; Ding, X.; Bijma, P.; de Koning, D.J.; Zhang, Q. Best Linear Unbiased Prediction of Genomic Breeding Values Using a Trait-Specific Marker-Derived Relationship Matrix. PLoS ONE 2010, 5, e12648. [Google Scholar] [CrossRef]
- Wang, H.; Misztal, I.; Aguilar, I.; Legarra, A.; Fernando, R.L.; Vitezica, Z.; Okimoto, R.; Wing, T.; Hawken, R.; Muir, W.M. Genome-Wide Association Mapping Including Phenotypes from Relatives without Genotypes in a Single-Step (SsGWAS) for 6-Week Body Weight in Broiler Chickens. Front. Genet. 2014, 5, 134. [Google Scholar] [CrossRef]
- Jiao, X.; Sherman, B.T.; Huang, D.W.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A Stateful Web Service to Facilitate Gene/Protein List Analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef]
- Salem, M.M.I.; Amin, A.M.S.; Ashour, A.F.; El Nagar, A.G. Estimation of Genetic Parameters for Semen Traits in Egyptian Buffalo Bulls. Trop. Anim. Health Prod. 2023, 55, 264. [Google Scholar] [CrossRef]
- Khattab, A.S.; Zeweil, H.S.; Ahmed, M.H.; Asmaa, Z.M.; Mourd, K.A. A Comparison of Different Selection Indexes for Some Semen Traits on Egyptian Buffalo Bulls. Middle East J. Appl. Sci. 2015, 5, 368–373. [Google Scholar]
- Khattab, A.S.; Adenaike, A.S.; Shymma, M.E.K.; El-Gendy, M.M.; Peters, S.O.; Hussein, A.M. Genetic Evaluation of Semen Traits in Friesian Bulls Raised in Egypt. Theriogenology 2022, 179, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, K.G.M.; El-Sokary, A.A.E.; El-Roos, M.E.A.A.; Ghaffar, A.D.A.; Nawito, M. Sperm Characteristics in Cryopreserved Buffalo Bull Semen and Field Fertility. Iran. J. Appl. Anim. Sci. 2013, 3, 777–783. [Google Scholar]
- Gabr, A.A.W.; El Basuini, M.F. Effect of Tonophosphan, Zinc Oxide, and Ascorbic Acid on Semen, Sexual Desire, and the Fertility Rate of Egyptian Buffalo Bulls. Ann. Agric. Sci. 2018, 63, 215–221. [Google Scholar] [CrossRef]
- Rushdi, H.E.; Saad, M.F.; Saeed, A.M. Association of Chromosomal Aberrations and Semen Quality in Egyptian Buffalo Bulls. Egypt. J. Anim. Prod. 2017, 54, 95–103. [Google Scholar] [CrossRef]
- Kumar, D.; Mehta, J.S.; Jerome, A.; Kumar, P.; Kumar, D.; Shivani, B.; Patil, C.S.; Bala, R.; Verma, N.; Nain, S.; et al. Genetic Analysis for Semen Quality Traits in Buffalo Bulls. Trop. Anim. Health Prod. 2023, 55, 313. [Google Scholar] [CrossRef]
- Qin, C.; Yin, H.; Zhang, X.; Sun, D.; Zhang, Q.; Liu, J.; Ding, X.; Zhang, Y.; Zhang, S. Genome-Wide Association Study for Semen Traits of the Bulls in Chinese Holstein. Anim. Genet. 2017, 48, 80–84. [Google Scholar] [CrossRef]
- Rezende, F.M.; Dietsch, G.O.; Peñagaricano, F. Genetic Dissection of Bull Fertility in US Jersey Dairy Cattle. Anim. Genet. 2018, 49, 393–402. [Google Scholar] [CrossRef]
- Mueller, J.L.; Skaletsky, H.; Brown, L.G.; Zaghlul, S.; Rock, S.; Graves, T.; Auger, K.; Warren, W.C.; Wilson, R.K.; Page, D.C. Independent Specialization of the Human and Mouse X Chromosomes for the Male Germ Line. Nat. Genet. 2013, 45, 1083–1087. [Google Scholar] [CrossRef]
- Liu, W.S. Mammalian Sex Chromosome Structure, Gene Content, and Function in Male Fertility. Annu. Rev. Anim. Biosci. 2019, 7, 103–124. [Google Scholar] [CrossRef]
- Fortes, M.R.S.; Porto-Neto, L.R.; Satake, N.; Nguyen, L.T.; Freitas, A.C.; Melo, T.P.; Scalez, D.C.B.; Hayes, B.; Raidan, F.S.S.; Reverter, A.; et al. X Chromosome Variants Are Associated with Male Fertility Traits in Two Bovine Populations. Genet. Sel. Evol. 2020, 52, 46. [Google Scholar] [CrossRef]
- Fortes, M.R.S.; Reverter, A.; Kelly, M.; Mcculloch, R.; Lehnert, S.A. Genome-Wide Association Study for Inhibin, Luteinizing Hormone, Insulin-like Growth Factor 1, Testicular Size and Semen Traits in Bovine Species. Andrology 2013, 1, 644–650. [Google Scholar] [CrossRef]
- Suchocki, T.; Szyda, J. Genome-Wide Association Study for Semen Production Traits in Holstein-Friesian Bulls. J. Dairy Sci. 2015, 98, 5774–5780. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, H.A.; Rezende, F.M.; Peñagaricano, F. Gene Mapping and Genomic Prediction of Bull Fertility Using Sex Chromosome Markers. J. Dairy Sci. 2020, 103, 3304–3311. [Google Scholar] [CrossRef] [PubMed]
- Hering, D.M.; Oleński, K.; Ruść, A.; Kaminski, S. Genome-Wide Association Study for Semen Volume and Total Number of Sperm in Holstein-Friesian Bulls. Anim. Reprod. Sci. 2014, 151, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Diaz, J.; Cenadelli, S.; Bornaghi, V.; Bongioni, G.; Montedoro, S.M.; Achilli, A.; Capelli, C.; Rincon, J.C.; Milanesi, M.; Passamonti, M.M.; et al. Identification of Genomic Regions Associated with Total and Progressive Sperm Motility in Italian Holstein Bulls. J. Dairy Sci. 2023, 106, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Sweett, H.; Fonseca, P.A.S.; Suárez-Vega, A.; Livernois, A.; Miglior, F.; Cánovas, A. Genome-Wide Association Study to Identify Genomic Regions and Positional Candidate Genes Associated with Male Fertility in Beef Cattle. Sci. Rep. 2020, 10, 20102. [Google Scholar] [CrossRef]
- Dickins, R.A.; Frew, I.J.; House, C.M.; O’Bryan, M.K.; Holloway, A.J.; Haviv, I.; Traficante, N.; de Kretser, D.M.; Bowtell, D.D.L. The Ubiquitin Ligase Component Siah1a Is Required for Completion of Meiosis I in Male Mice. Mol. Cell. Biol. 2002, 22, 2294–2303. [Google Scholar] [CrossRef]
- Weon, J.L.; Potts, P.R. The MAGE Protein Family and Cancer. Curr. Opin. Cell Biol. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Lee, A.K.; Potts, P.R. A Comprehensive Guide to the MAGE Family of Ubiquitin Ligases. J. Mol. Biol. 2017, 429, 1114–1142. [Google Scholar] [CrossRef]
- Park, S.K.; Yoon, J.; Wang, L.; Shibata, T.K.; Motamedchaboki, K.; Shim, K.J.; Chang, M.S.; Lee, S.H.; Tamura, N.; Hatakeyama, S.; et al. Enhancement of Mouse Sperm Motility by Trophinin-Binding Peptide. Reprod. Biol. Endocrinol. 2012, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, L.I.; Pastuszak, A.W.; Goldstein, A.T.; Giraldi, A.; Perelman, M.A. Management of Sexual Dysfunction in Men and Women: An Interdisciplinary Approach; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 9781493931002. [Google Scholar]
- Wang, R.S.; Yeh, S.; Tzeng, C.R.; Chang, C. Androgen Receptor Roles in Spermatogenesis and Fertility: Lessons from Testicular Cell-Specific Androgen Receptor Knockout Mice. Endocr. Rev. 2009, 30, 119–132. [Google Scholar] [CrossRef]
- Ito, H.; Langenhorst, T.; Ogden, R.; Inoue-Murayama, M. Androgen Receptor Gene Polymorphism in Zebra Species. Meta Gene 2015, 5, 120–123. [Google Scholar] [CrossRef]
- Lin, C.; Tholen, E.; Jennen, D.; Ponsuksili, S.; Schellander, K.; Wimmers, K. Evidence for Effects of Testis and Epididymis Expressed Genes on Sperm Quality and Boar Fertility Traits. Reprod. Domest. Anim. 2006, 41, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Sarakul, M.; Elzo, M.A.; Koonawootrittriron, S.; Suwanasopee, T.; Jattawa, D.; Laodim, T. Characterization of Biological Pathways Associated with Semen Traits in the Thai Multibreed Dairy Population. Anim. Reprod. Sci. 2018, 197, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Song, N.N.; Cheng, X.; Zhu, L.; Zhang, Q.; Zhang, L.L.; Yang, X.; Wang, H.; Kong, Q.; Shi, D.; et al. ZC4H2 Stabilizes RNF220 to Pattern Ventral Spinal Cord through Modulating Shh/Gli Signaling. J. Mol. Cell Biol. 2020, 12, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Mao, B. The Many Faces of the E3 Ubiquitin Ligase, RNF220, in Neural Development and Beyond. Dev. Growth Differ. 2022, 64, 98–105. [Google Scholar] [CrossRef]
- Fu, J.; Wang, Y.; Fok, K.L.; Yang, D.; Qiu, Y.; Chan, H.C.; Koide, S.S.; Miao, S.; Wang, L. Anti-ACTL7a Antibodies: A Cause of Infertility. Fertil. Steril. 2012, 97, 1226–1233.e8. [Google Scholar] [CrossRef]
- Fortes, M.R.S.; Reverter, A.; Hawken, R.J.; Bolormaa, S.; Lehnert, S.A. Candidate Genes Associated with Testicular Development, Sperm Quality, and Hormone Levels of Inhibin, Luteinizing Hormone, and Insulin-like Growth Factor 1 in Brahman Bulls. Biol. Reprod. 2012, 87, 58. [Google Scholar] [CrossRef]
- El-Maghrabi, M.R.; Correiat, J.J.; Heil, P.J.; Pate, T.M.; Cobb, C.E.; Pilkis, S.J. Tissue distribution, immunoreactivity, and physical properties of 6-phosphofructo-2-kinase/fructose-2, 6-bisphosphatase. Proc. Natl. Acad. Sci. USA 1986, 83, 5005–5009. [Google Scholar] [CrossRef]
- Umehara, T.; Tsujita, N.; Shimada, M.; ActiUmehara, T.; Tsujita, N.; Shimada, M. Activation of Toll-like Receptor 7/8 Encoded by the X Chromosome Alters Sperm Motility and Provides a Novel Simple Technology for Sexing Sperm. PLoS Biol. 2019, 17, e3000398. [Google Scholar] [CrossRef] [PubMed]
- Winge, S.B.; Dalgaard, M.D.; Belling, K.G.; Jensen, J.M.; Nielsen, J.E.; Aksglaede, L.; Schierup, M.H.; Brunak, S.; Skakkebæk, N.E.; Juul, A.; et al. Transcriptome Analysis of the Adult Human Klinefelter Testis and Cellularity-Matched Controls Reveals Disturbed Differentiation of Sertoli- and Leydig Cells Article. Cell Death Dis. 2018, 9, 586. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Liang, G.; Martin, G.B.; Guan, L.L. Functional Changes in mRNA Expression and Alternative Pre-mRNA Splicing Associated with the Effects of Nutrition on Apoptosis and Spermatogenesis in the Adult Testis. BMC Genomics 2017, 18, 64. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sun, P.; Hu, Y.; Shan, L.; Geng, Q.; Gong, Y.; Fan, H.; Zhang, T.; Zhou, Y. Elevated Testicular Apoptosis Is Associated with Elevated Sphingosine Driven by Gut Microbiota in Prediabetic Sheep. BMC Biol. 2022, 20, 121. [Google Scholar] [CrossRef] [PubMed]
- Özbek, M.; Hitit, M.; Kaya, A.; Jousan, F.D.; Memili, E. Sperm Functional Genome Associated With Bull Fertility. Front. Vet. Sci. 2021, 8, 610888. [Google Scholar] [CrossRef]
- Ma, D.D.; Wang, D.H.; Yang, W.X. Kinesins in Spermatogenesis. Biol. Reprod. 2017, 96, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.S.; Kehrl, J.H. Pyk2 Amplifies Epidermal Growth Factor and C-Src-Induced Stat3 Activation. J. Biol. Chem. 2004, 279, 17224–17231. [Google Scholar] [CrossRef]
- Wang, H.; Hong, X.; Kinsey, W.H. Sperm-Oocyte Signaling: The Role of IZUMO1R and CD9 in PTK2B Activation and Actin Remodeling at the Sperm Binding Site. Biol. Reprod. 2021, 104, 1292–1301. [Google Scholar] [CrossRef]
- Yang, F.; Chen, F.; Li, L.; Yan, L.; Badri, T.; Lv, C.; Yu, D.; Zhang, M.; Jang, X.; Li, J.; et al. Three Novel Players: PTK2B, SYK, and TNFRSF21 Were Identified to Be Involved in the Regulation of Bovine Mastitis Susceptibility via GWAS and Post-Transcriptional Analysis. Front. Immunol. 2019, 10, 1579. [Google Scholar] [CrossRef]
- Pastorekova, S.; Parkkila, S.; Pastorek, J.; Supuran, C.T. Carbonic Anhydrases: Current State of the Art, Therapeutic Applications and Future Prospects. J. Enzyme Inhib. Med. Chem. 2004, 19, 199–229. [Google Scholar] [CrossRef]
- de Camargo, G.M.F.; Aspilcueta-Borquis, R.R.; Fortes, M.R.S.; Porto-Neto, R.; Cardoso, D.F.; Santos, D.J.A.; Lehnert, S.A.; Reverter, A.; Moore, S.S.; Tonhati, H. Prospecting Major Genes in Dairy Buffaloes. BMC Genomics 2015, 16, 872. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.E.; Baldi, F.S.; Alexandre, P.A.; Santana, M.H.A.; Ventura, R.V.; Bueno, R.S.; Bonin, M.N.; Rezende, F.M.; Coutinho, L.L.; Eler, J.P.; et al. Genomic Regions and Genes Associated with Carcass Quality in Nelore Cattle. Genet. Mol. Res. 2019, 18, 1–15. [Google Scholar] [CrossRef]
- Ding, Q.; Joshi, P.S.; Xie, Z.H.; Xiang, M.; Gan, L. BARHL2 Transcription Factor Regulates the Ipsilateral/ Contralateral Subtype Divergence in Postmitotic DI1 Neurons of the Developing Spinal Cord. Proc. Natl. Acad. Sci. USA 2012, 109, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, X.; Wang, W. LncRNA and MRNA Sequencing of the Left Testis in Experimental Varicocele Rats Treated with Morinda Officinalis Polysaccharide. Exp. Ther. Med. 2021, 22, 1136. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, R.; Montanari, M.; Chiarella, P.; Stefanini, M.; Boitani, C. Regulatory Role of BMP2 and BMP7 in Spermatogonia and Sertoli Cell Proliferation in the Immature Mouse. Eur. J. Endocrinol. 2004, 151, 511–520. [Google Scholar] [CrossRef]
- Conklin, D.; Holderman, S.; Whitmore, T.E.; Maurer, M.; Feldhaus, A.L. Molecular Cloning, Chromosome Mapping and Characterization of UBQLN3 a Testis-Specific Gene That Contains an Ubiquitin-like Domain. Gene 2000, 249, 91–98. [Google Scholar] [CrossRef]
- Zhao, W.; Li, Z.; Ping, P.; Wang, G.; Yuan, X.; Sun, F. Outer Dense Fibers Stabilize the Axoneme to Maintain Sperm Motility. J. Cell. Mol. Med. 2018, 22, 1755–1768. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Belkacemi, A.; Ayesh, B.M.; Simone Becker, L.; Sindiani, A.M.; Fischer, U.; Hammadeh, M.; Keller, A.; Meese, E. MicroRNA-Targeting in Spermatogenesis: Over-Expressions of MicroRNA-23a/b-3p and Its Affected Targeting of the Genes ODF2 and UBQLN3 in Spermatozoa of Patients with Oligoasthenozoospermia. Andrology 2021, 9, 1137–1144. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait 1 | Mean | SD | Min | Max | CV (%) |
|---|---|---|---|---|---|
| Ejaculate volume, mL | 3.78 | 1.79 | 0.50 | 11.50 | 47.35 |
| Mass motility, % | 62.15 | 16.30 | 5.00 | 95.00 | 26.23 |
| Livability, % | 61.07 | 15.59 | 3.00 | 88.00 | 25.53 |
| Abnormality, % | 4.47 | 2.61 | 1.00 | 44.00 | 58.39 |
| Concentration 2 | 0.76 | 0.35 | 0.20 | 3.75 | 46.05 |
| Chr. | Number of SNPs | MAF | Chr. Length [Mb] | Gap Size [kb] # | |||||
|---|---|---|---|---|---|---|---|---|---|
| Initial | Filtered * | Initial | Filtered * | Initial | Filtered * | Mean | SD | Max | |
| 1 | 8533 | 5526 | 0.25 | 0.29 | 201.95 | 201.95 | 36.55 | 22.36 | 716.57 |
| 2 | 7786 | 4993 | 0.24 | 0.29 | 188.83 | 188.83 | 37.83 | 27.05 | 699.06 |
| 3 | 7192 | 4564 | 0.27 | 0.31 | 175.41 | 175.39 | 38.44 | 27.10 | 762.66 |
| 4 | 6499 | 4192 | 0.27 | 0.29 | 165.14 | 165.14 | 39.40 | 29.32 | 719.43 |
| 5 | 5027 | 3229 | 0.29 | 0.30 | 127.53 | 127.50 | 39.50 | 40.53 | 1721.08 |
| 6 | 5064 | 3264 | 0.29 | 0.29 | 120.32 | 120.24 | 36.85 | 23.83 | 477.34 |
| 7 | 4722 | 3027 | 0.29 | 0.30 | 117.10 | 117.10 | 38.70 | 23.74 | 418.96 |
| 8 | 5031 | 3244 | 0.28 | 0.29 | 119.61 | 119.50 | 36.85 | 22.03 | 512.54 |
| 9 | 4341 | 2825 | 0.27 | 0.30 | 110.10 | 109.97 | 38.94 | 28.94 | 689.09 |
| 10 | 3929 | 2539 | 0.28 | 0.29 | 104.07 | 104.07 | 41.01 | 24.94 | 414.49 |
| 11 | 4012 | 2572 | 0.27 | 0.29 | 102.20 | 102.05 | 39.69 | 27.67 | 609.86 |
| 12 | 4461 | 2880 | 0.27 | 0.28 | 106.40 | 106.37 | 36.95 | 19.29 | 180.88 |
| 13 | 3385 | 2154 | 0.28 | 0.29 | 90.40 | 90.22 | 41.90 | 45.88 | 1229.15 |
| 14 | 3342 | 2146 | 0.27 | 0.29 | 83.09 | 82.88 | 38.64 | 23.93 | 501.28 |
| 15 | 3428 | 2192 | 0.28 | 0.28 | 82.03 | 82.03 | 37.44 | 21.27 | 197.54 |
| 16 | 3185 | 2040 | 0.29 | 0.30 | 84.41 | 84.36 | 41.37 | 43.17 | 1139.89 |
| 17 | 2937 | 1873 | 0.28 | 0.29 | 73.12 | 72.68 | 38.82 | 24.00 | 398.63 |
| 18 | 2737 | 1728 | 0.29 | 0.30 | 65.76 | 65.69 | 38.04 | 32.20 | 560.14 |
| 19 | 2968 | 1940 | 0.28 | 0.30 | 71.58 | 71.54 | 36.90 | 18.94 | 235.90 |
| 20 | 2637 | 1691 | 0.30 | 0.31 | 68.47 | 68.45 | 40.50 | 28.75 | 386.39 |
| 21 | 2557 | 1703 | 0.28 | 0.29 | 60.72 | 60.58 | 35.59 | 18.07 | 231.64 |
| 22 | 2529 | 1633 | 0.28 | 0.30 | 61.97 | 61.69 | 37.80 | 20.60 | 173.44 |
| 23 | 2172 | 1410 | 0.32 | 0.34 | 51.44 | 51.18 | 36.32 | 21.94 | 388.82 |
| 24 | 1779 | 1152 | 0.30 | 0.34 | 42.13 | 42.00 | 36.49 | 22.36 | 264.13 |
| X-chromosome | 4891 | 2765 | 0.31 | 0.31 | 143.40 | 142.37 | 51.51 | 62.41 | 1427.38 |
| UnPos | 17,896 | --- | 0.25 | --- | --- | --- | --- | --- | --- |
| Overall | 123,040 | 67,282 | 0.28 | 0.30 | 2617 | 2614 | 38.88 | 28.01 | 1721.08 |
| Chromsome No. | Genomic Map Position (Mb) | Trait (Genomic Explained Variance %) | Gene ID (The Nearest Genes to the Position of the Chromosomal Window Segment) |
|---|---|---|---|
| X-chromosome | 48.19–121.68 | VOL (4.18%) | LOC102414874, LOC102401551, LOC102398394, TRO, CHRDL1, GRIA3, LOC112582129, AR, OPHN1, COL4A6 |
| X-chromosome | 29.35–127.08 | MM (4.59%) | ZC4H2, ZC3H12B, LOC102391626, SMARCA1, DMD, GNL3L, ITIH6, P2RY10, LOC102415058, RTL9, AMMECR1 |
| X-chromosome | 29.12–127.08 | LS (5.16%) | ZC4H2, ZC3H12B, LOC102391626, SMARCA1, DMD, GNL3L, ITIH6, P2RY10, LOC102415058, RTL9, AMMECR1, POU3F4, LOC102404920 |
| X-chromosome | 29.12–127.08 | AS (5.19%) | PFKFB1, TBX22, GPR174, DACH2, LOC112582031, LOC102391626, SMARCA1, POU3F4, LOC102404920, RTL9, AMMECR1, DMD |
| X-chromosome | 47.85–142.21 | CONC (4.31%) | BCAP31, P2RY10, LOC102415058, LOC102414874, LOC102401551, LOC102391626, SMARCA1, POU3F4, LOC102404920, LOC102394334, ZIC3, MAGEH1, FOXR2 |
| 3 | 58.52–137.06 | VOL (0.99%) | KIF2B, LOC102390200, PTK2B |
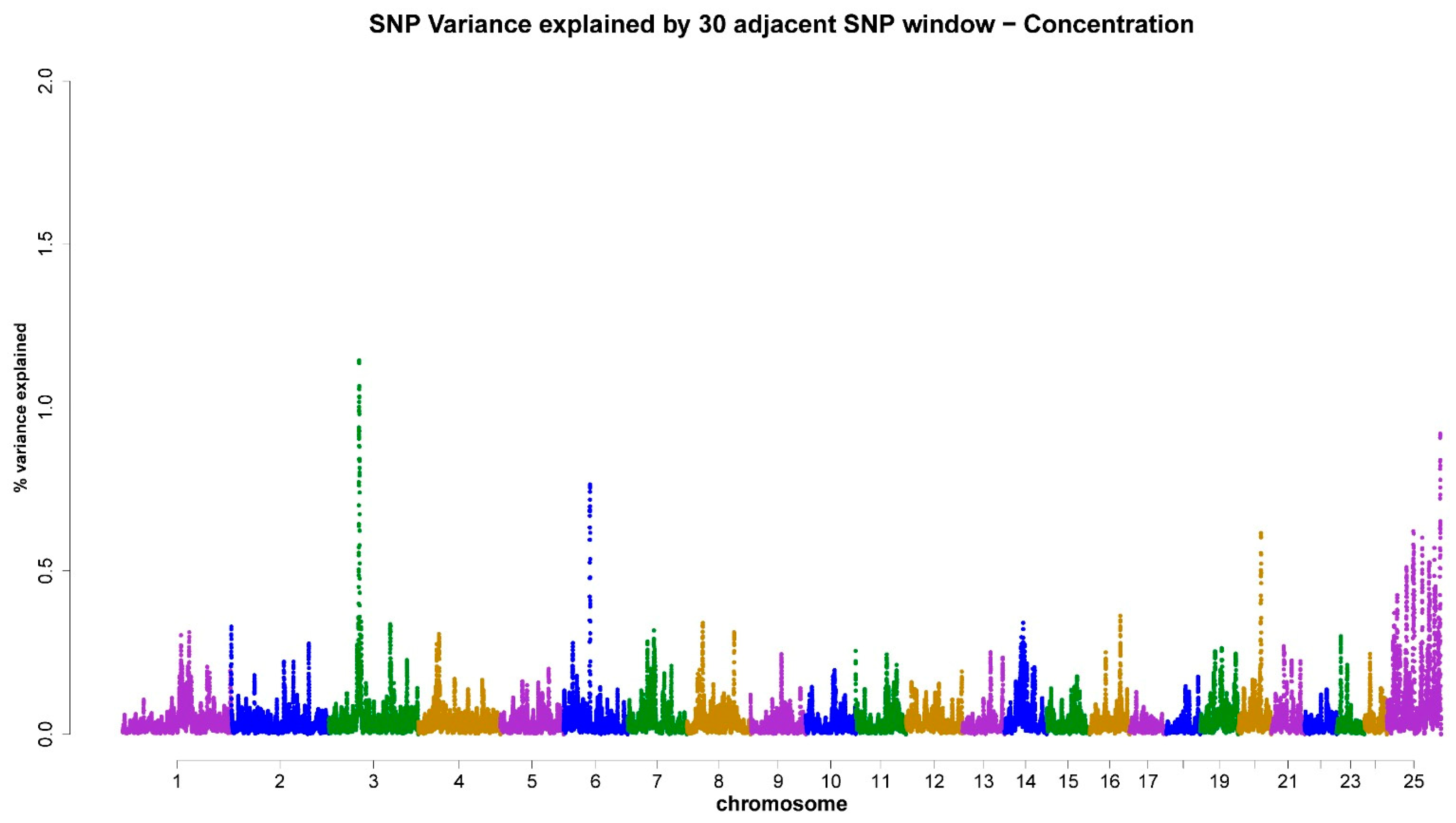
| 3 | 60.92–62.73 | CONC (1.14%) | CA10 |
| 4 | 43.42–44.85 | VOL (0.42%) | CSF2RB |
| 6 | 51.58–52.47 | CONC (0.76%) | ZNF326, BARHL2 |
| 7 | 53.23–54.25 | MM (0.45%) | KCTD8, YIPF7 |
| 7 | 53.26–54.27 | AS (0.57%) | KCTD8, YIPF7 |
| 14 | 34.03–35.39 | VOL (0.40%) | FERMT1, BMP2 |
| 16 | 33.86–36.95 | MM (0.51%) | LOC102402176, LOC102397107 |
| 16 | 33.86–36.95 | LS (0.47%) | LOC102402176, LOC102397107 |
| 16 | 33.86–36.95 | AS (0.49%) | LOC102402176, LOC102397107 |
| 20 | 46.22–47.48 | MM (0.51%) | BLM, CRTC3 |
| 20 | 46.22–47.48 | LS (0.57%) | BLM, CRTC3 |
| 20 | 46.22–47.48 | CONC (0.61%) | BLM, CRTC3 |
| 22 | 49.91–50.83 | AS (0.51%) | LOC102389067, CDH7 |
| Candidate Gene 2 | Chromosome No. | Gene Function |
|---|---|---|
| LOC102414874 | X-chromosome | E3 ubiquitin-protein ligase SIAH1-like |
| LOC102401551 | X-chromosome | Dynein light chain 1, cytoplasmic-like |
| LOC102398394 | X-chromosome | Melanoma-associated antigen D2 |
| TRO | X-chromosome | Trophinin |
| CHRDL1 | X-chromosome | Chordin like 1 |
| GRIA3 | X-chromosome | Glutamate ionotropic receptor AMPA type subunit 3 |
| LOC112582129 | X-chromosome | Cancer/testis antigen 47A-like |
| AR | X-chromosome | Androgen receptor |
| OPHN1 | X-chromosome | Oligophrenin 1 |
| COL4A6 | X-chromosome | Collagen type IV alpha 6 chain |
| ZC4H2 | X-chromosome | Zinc finger C4H2-type containing |
| ZC3H12B | X-chromosome | Zinc finger CCCH-type containing 12B |
| LOC102391626 | X-chromosome | Actin-related protein T1 |
| SMARCA1 | X-chromosome | SWI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily a, member 1 |
| DMD | X-chromosome | Dystrophin |
| GNL3L | X-chromosome | G protein nucleolar 3, like |
| ITIH6 | X-chromosome | Inter-alpha-trypsin inhibitor heavy chain family member 6 |
| P2RY10 | X-chromosome | Putative P2Y purinoceptor 10 |
| LOC102415058 | X-chromosome | P2Y receptor family member 10 |
| RTL9 | X-chromosome | Retrotransposon Gag like 9 |
| AMMECR1 | X-chromosome | AMMECR nuclear protein 1 |
| POU3F4 | X-chromosome | POU class 3 homeobox 4 |
| LOC102404920 | X-chromosome | Sodium/hydrogen exchanger 2-like |
| PFKFB1 | X-chromosome | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 1 |
| TBX22 | X-chromosome | T-box transcription factor 22 |
| GPR174 | X-chromosome | G protein-coupled receptor 174 |
| DACH2 | X-chromosome | Dachshund family transcription factor 2 |
| LOC112582031 | X-chromosome | Protein FAM209B-like |
| BCAP31 | X-chromosome | B cell receptor associated protein 31 |
| LOC102394334 | X-chromosome | Putative MAGE domain-containing protein MAGEA13P |
| ZIC3 | X-chromosome | Zic family member 3 |
| MAGEH1 | X-chromosome | MAGE family member H1 |
| FOXR2 | X-chromosome | Forkhead box R2 |
| KIF2B | 3 | Kinesin family member 2B |
| LOC102390200 | 3 | Peptidyl-prolyl cis-trans isomerase A-like |
| PTK2B | 3 | Protein tyrosine kinase 2 beta |
| CA10 | 3 | Carbonic anhydrase 10 |
| CSF2RB | 4 | Colony stimulating factor 2 receptor subunit beta |
| ZNF326 | 6 | Zinc finger protein 326 |
| BARHL2 | 6 | BarH like homeobox 2 |
| KCTD8 | 7 | Potassium channel tetramerization domain containing 8 |
| YIPF7 | 7 | Yip1 domain family member 7 |
| FERMT1 | 14 | FERM domain containing kindlin 1 |
| BMP2 | 14 | Bone morphogenetic protein 2 |
| LOC102402176 | 16 | Ubiquilin-1 or Ubiquitin-like proteins |
| LOC102397107 | 16 | Ubiquilin-3 |
| BLM | 20 | BLM RecQ-like helicase |
| CRTC3 | 20 | CREB-regulated transcription coactivator 3 |
| LOC102389067 | 22 | Serpin B8 |
| CDH7 | 22 | Cadherin 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
EL Nagar, A.G.; Salem, M.M.I.; Amin, A.M.S.; Khalil, M.H.; Ashour, A.F.; Hegazy, M.M.; Abdel-Shafy, H. A Single-Step Genome-Wide Association Study for Semen Traits of Egyptian Buffalo Bulls. Animals 2023, 13, 3758. https://doi.org/10.3390/ani13243758
EL Nagar AG, Salem MMI, Amin AMS, Khalil MH, Ashour AF, Hegazy MM, Abdel-Shafy H. A Single-Step Genome-Wide Association Study for Semen Traits of Egyptian Buffalo Bulls. Animals. 2023; 13(24):3758. https://doi.org/10.3390/ani13243758
Chicago/Turabian StyleEL Nagar, Ayman G., Mohamed M. I. Salem, Amin M. S. Amin, Maher H. Khalil, Ayman F. Ashour, Mohammed M. Hegazy, and Hamdy Abdel-Shafy. 2023. "A Single-Step Genome-Wide Association Study for Semen Traits of Egyptian Buffalo Bulls" Animals 13, no. 24: 3758. https://doi.org/10.3390/ani13243758
APA StyleEL Nagar, A. G., Salem, M. M. I., Amin, A. M. S., Khalil, M. H., Ashour, A. F., Hegazy, M. M., & Abdel-Shafy, H. (2023). A Single-Step Genome-Wide Association Study for Semen Traits of Egyptian Buffalo Bulls. Animals, 13(24), 3758. https://doi.org/10.3390/ani13243758