
Evaluating Different Factors That Affect the Nesting Patterns of European and Algerian Hedgehogs in Urban and Suburban Environments

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
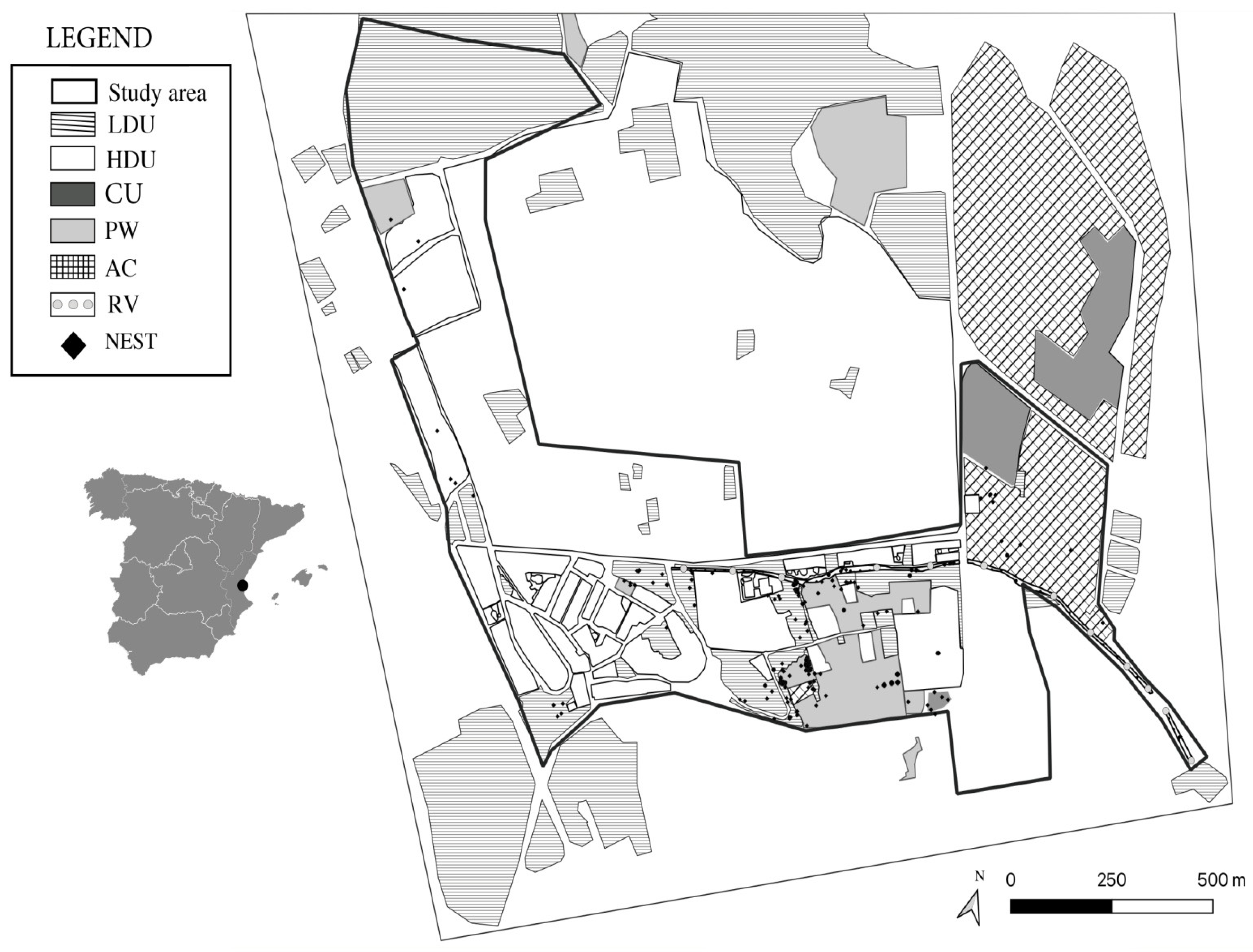
2.1. Study Area
2.2. Fieldwork
2.3. Habitat Characterization
2.4. Data Analysis
3. Results
3.1. Macrohabitats
3.2. Subhabitats
4. Discussion
4.1. General
4.2. Macrohabitats
4.3. Subhabitats
4.4. Nest Sharing and Parasites
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, G.; Fang, C.; Li, Y. Global impacts of future urban expansion on terrestrial vertebrate diversity. Nat. Commun. 2022, 13, 1628. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Lin, Y.; An, W.; Gan, M.; Shahtahmassebi, A.; Ye, Z.; Huang, L.; Zhu, C.; Huang, L.; Zhang, J.; Wang, K. Spatial Grain Effects of Urban Green Space Cover Maps on Assessing Habitat Fragmentation and Connectivity. Land 2021, 10, 1065. [Google Scholar] [CrossRef]
- Bateman, P.W.; Fleming, P.A. Big city life: Carnivores in urban environments. J. Zool. 2012, 287, 1–23. [Google Scholar] [CrossRef]
- Baker, P.J.; Ansell, R.J.; Dodds, P.A.; Webber, C.E.; Harris, S. Factors affecting the distribution of small mammals in an urban area. Mamm. Rev. 2003, 33, 95–100. [Google Scholar] [CrossRef]
- Gomes, V.; Ribeiro, R.; Carretero, M.A. Effects of urban habitat fragmentation on common small mammals: Species versus communities. Biodivers. Conserv. 2011, 20, 3577–3590. [Google Scholar] [CrossRef]
- Johnson, A.M.; Karels, T.J. Partitioning the effects of habitat fragmentation on rodent species richness in an urban landscape. Urban. Ecosyst. 2016, 19, 547–560. [Google Scholar] [CrossRef]
- Gago, H.; Ruiz-Fons, F.; Drechsler, R.M.; Alambiaga, I.; Monrós, J.S. Patterns of adult tick parasitization of coexisting European (Erinaceus europaeus) and Algerian (Atelerix algirus) hedgehog populations in eastern Iberia. Ticks Tick Borne Dis. 2022, 13, 102048. [Google Scholar] [CrossRef]
- Gago, H.; Drechsler, R.M.; Monrós, J.S. Algerian and European hedgehogs cohabiting in periurban environments: Spatial behaviour and habitat use. Eur. J. Wildl. Res. 2023, 69, 19. [Google Scholar] [CrossRef]
- Nores, C. Erinaceus europaeus Linnaeus 1758. In Atlas y Libro Rojo de los Mamíferos Terrestres de España; Palomo, L.J., Gisbert, J., Blanco, J.C., Eds.; Dirección General para la Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007; pp. 80–523. [Google Scholar]
- Morris, P. Winter Nests of the Hedgehog (Erinaceus europaeus L.). Oecologia 1973, 11, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Reeve, N.; Morris, P. Construction and use of summer nests by the hedgehog (Erinaceus europaeus). Mammalia 1985, 49, 187–194. [Google Scholar] [CrossRef]
- Bearman-Brown, L.E.; Philip, J.B.; Dawn, S.; Antonio, U.; Luke, E.; Richard, W.Y. Over-Winter Survival and Nest Site Selection of the West-European Hedgehog (Erinaceus europaeus) in Arable Dominated Landscapes. Animals 2020, 10, 1449. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.B. Experimental removal of introduced hedgehogs improves wader nest success in the Western Isles, Scotland. J. Appl. Ecol. 2001, 38, 802–812. [Google Scholar] [CrossRef]
- Haigh, A.; O’Riordan, R.M.; Butler, F. Nesting behaviour and seasonal body mass changes in a rural Irish population of the Western hedgehog (Erinaceus europaeus). Acta Theriol. 2012, 57, 321–331. [Google Scholar] [CrossRef]
- Rautio, A.; Valtonen, A.; Auttila, M.; Kunnasranta, M. Nesting Patterns of European Hedgehogs (Erinaceus Europaeus) under Northern Conditions. Acta Theriol. 2014, 59, 173–181. [Google Scholar] [CrossRef]
- Huijser, M.P. Human impact on populations of hedgehogs Erinaceus europaeus through traffic and changes in the landscape: A review. Lutra 1999, 42, 39–56. [Google Scholar]
- Riber, A.B. Habitat use and behaviour of European hedgehog Erinaceus europaeus in a Danish rural area. Acta Theriol. 2006, 51, 363–371. [Google Scholar] [CrossRef]
- Jensen, A.B. Overwintering of European hedgehogs Erinaceus europaeus in a Danish rural area. Acta Theriol. 2004, 49, 145–155. [Google Scholar] [CrossRef]
- Boitani, L.; Reggiani, G. Movements and activity patters of hedgehogs (Erinaceus europaeus) in Mediterranean coastal habitats. Z. Säugetierkd 1984, 49, 193–206. [Google Scholar]
- García, S.; Puig, X.; Peris, A. Actividad y uso del hábitat por parte del Erizo Europeo (Erinaceus europaeus Linnaeus, 1758) en el Parque Natural de la Serralada de Marina (Barcelona, Cataluña). Galemys 2009, 21, 13–23. [Google Scholar]
- Cahill, S.; Llimona, F.; Tenés, A.; Carles, S.; Cabañeros, L. Radioseguimiento post recuperación de erizos europeos (Erinaceus europaeus Linnaeus, 1758) en el Parque Natural de la Sierra de Collserola (Barcelona). Galemys 2011, 23, 2011. [Google Scholar] [CrossRef]
- Reeve, N. A Field Study of the Hedgehog (Erinaceus europaeus) with Particular Reference to Movements and Behaviour. Ph.D. Thesis, University of London, London, UK, 1981. [Google Scholar]
- García, S.; Puig-Montserrat, X. Algerian Hedgehog (Atelerix algirus Lereboullet, 1842) Habitat Selection at the Northern Limit of Its Range. Galemys 2014, 26, 49–56. [Google Scholar] [CrossRef]
- Pérez-Cueva, A. Atlas Climátic de la Comunitat Valenciana; Direcció General d’Urnanisme i Ordenació del Territori: Generalitat Valenciana, Spain, 1994. [Google Scholar]
- Rautio, A.; Valtonen, A.; Kunnasranta, M. The Effects of Sex and Season on Home Range in European Hedgehogs at the Northern Edge of the Species Range. Ann. Zool. Fenn. 2013, 50, 107–123. [Google Scholar] [CrossRef]
- Hof, A.; Bright, P. The value of green-spaces in built-up areas for western hedgehogs. Lutra 2009, 52, 69–82. [Google Scholar]
- Bunnell, T. The assessment of British Hedgehog (Erinaceus europaeus) casualties on arrival and determination of optimum release weights using a new index. J. Wildl. Rehabil. 2002, 25, 11–21. [Google Scholar]
- White, G.C.; Garrott, R.A. Analysis of Wildlife Radio-Tracking Data; Academic Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Young, R.P.; Davison, J.; Trewby, I.D.; Wilson, G.J.; Delahay, R.J.; Doncaster, C.P. Abundance of hedgehogs (Erinaceus europaeus) in relation to the density and distribution of badgers (Meles meles). J. Zool. Lond. 2006, 269, 349–356. [Google Scholar] [CrossRef]
- Manly, B.F.J.; McDonald, L.L.; Thomas, D.L.; McDonald, T.L.; Erickson, W.P. Resource Selection by Animals. Statistical Design and Analysis for Field Studies; Kluwer Academic Publishers: Philip Drive Norwell, MA, USA , 2002. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R, Statistics for Biology and Health; Springer Science: Berlin, Germany, 2009. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2023. Available online: http://qgis.osgeo.org/ (accessed on 4 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 15 January 2022).
- Pettett, C.E.; Moorhouse, T.P.; Johnson, P.J.; Macdonald, D.W. Factors affecting hedgehog (Erinaceus europaeus) attraction to rural villages in arable landscapes. Eur. J. Wildl. Res. 2017, 63, 54. [Google Scholar] [CrossRef]
- Rasmussen, S.; Dabelsteen, T.; Berg, T.; Jones, O. The ecology of suburban juvenile European hedgehogs (Erinaceus europaeus) in Denmark. Ecol. Evol. 2019, 9, 13174–13187. [Google Scholar] [CrossRef]
- Gazzard, A.; Baker, P.J. What makes a house a home? Nest box use by West European hedgehogs (Erinaceus europaeus) is influenced by nest box placement, resource provisioning and site-based factors. PeerJ 2022, 10, e13662. [Google Scholar] [CrossRef]
- Rast, W.; Barthel, L.M.F.; Berger, A. Music Festival Makes Hedgehogs Move: How Individuals Cope Behaviorally in Response to Human-Induced Stressors. Animals 2019, 9, 455. [Google Scholar] [CrossRef]
- Morris, P.; Meakin, K.; Sharafi, S. The Behaviour and Survival of Rehabilitated Hedgehogs (Erinaceus Europaeus). Anim. Welf. 1993, 2, 53–66. [Google Scholar] [CrossRef]
- Reeve, N. The survival and welfare of hedgehogs (Erinaceus europaeus) after release back into the wild. Anim. Welf. 1998, 7, 189–202. [Google Scholar] [CrossRef]
- Burel, F. Hedgerows and Their Role in Agricultural Landscapes. Crit. Rev. Plant. Sci. 1996, 15, 169–190. [Google Scholar] [CrossRef]
- Hinsley, S.A.; Bellamy, P.E. The influence of hedge structure, management and landscape context on the value of hedgerows to birds: A review. Environ. Manag. 2000, 60, 33–49. [Google Scholar] [CrossRef]
- Burel, F.; Butet, A.; Delettre, Y.R.; Millàn de la Peña, N. Differential response of selected taxa to landscape context and agricultural intensification. Landsc. Urban Plan. 2004, 67, 195–204. [Google Scholar] [CrossRef]
- Haigh, A. The Ecology of the European Hedgehog (Erinaceus europaeus) in Rural Ireland. Ph.D. Thesis, University College Cork, Cork, Ireland, 2011. [Google Scholar]
- Rym, S.; Mehdi, B.; Walid, D.; Khadidjawissal, A.; Charafeddine, S.; Feriel, S.; Slim, B. Environmental and Habitat Preferences of the Algerian Hedgehog Atelerix algirus (Lereboullet, 1842) in El Kala National Park (North-East Algeria). J. Biores. Manag. 2022, 9, 165–174. [Google Scholar]
- Rondinini, C.; Doncaster, P. Roads as barriers to movement for hedgehogs. Funct. Ecol. 2002, 16, 504–509. [Google Scholar] [CrossRef]
- Berger, A.; Barthel, L.M.F.; Rast, W.; Hofer, H.; Gras, P. Urban Hedgehog Behavioural Responses to Temporary Habitat Disturbance versus Permanent Fragmentation. Animals 2020, 10, 2109. [Google Scholar] [CrossRef]
- Bergers, P.J.; Nieuwenhuizen, W. Viability of hedgehog populations in central Netherlands. Lutra 1999, 42, 65–76. [Google Scholar]
- Molina-López, R.A.; Adelantado, C.; Arosemena, E.L.; Obón, E.; Darwich, L.; Calvo, M.A. Integument mycobiota of wild European hedgehogs (Erinaceus europaeus) from Catalonia, Spain. Int. Sch. Res. Not. 2012, 2012, 659754. [Google Scholar] [CrossRef] [PubMed]
- Molina-López, R.A.; Vidal, A.; Obón, E.; Martín, M.; Darwich, L. Multidrug-resistant Salmonella enterica serovar Typhimurium monophasic variant 4,12:i:- isolated from asymptomatic wildlife in a Catalonian wildlife rehabilitation center, Spain. J. Wildl. Dis. 2015, 51, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Garcês, A.; Poeta, P.; Soeiro, V.; Lóio, S.; Cardoso-Gomes, A.; Torres, C.; Pires, I. Pyometra caused by Staphylococcus lentus in a wild European hedgehog (Erinaceus europaeus). J. Wildl. Dis. 2019, 55, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.A.; Zarazaga, M.; Ben Sallem, R.; Jouini, A.; Ben Slama, K.; Torres, C. Antibiotic resistance in Escherichia coli in husbandry animals: The African perspective. Lett. Appl. Microbiol. 2017, 64, 318–334. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, R.; Edouard-Fournier, P.; Santos-Silva, M.; Amaro, F.; Bacellar, F.; Raoult, D. Molecular detection of Rickettsia felis, Rickettsia typhi and two genotypes closely related to Bartonella elizabethae. Am. J. Trop. Med. Hyg. 2006, 75, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Collares-Pereira, M.; Korver, H.; Cao Thi, B.V.; Santos-Reis, M.; Bellenger, E.; Baranton, G.; Terpstra, W.J. Analysis of Leptospira isolates from mainland Portugal and the Azores islands. FEMS Microbiol. Lett. 2000, 185, 181–187. [Google Scholar] [CrossRef]
- Riley, P.Y.; Chomel, B.B. Hedgehog Zoonoses. Emerg. Infect. Dis. 2005, 11, 1–5. [Google Scholar] [CrossRef]
- De la Fuente, J.; Kocan, M.; Estrada, A.; Venzal, M.; Soonenshine, D. Ticks as vectors of pathogens that cause disease in humans and animals. Front Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef]
- Ruiz-Fons, F.; Gilbert, L. The role of deer as vehicles to move ticks, Ixodes ricinus, between contrasting habitats. Int. J. Parasitol. 2010, 40, 1013–1020. [Google Scholar] [CrossRef]

{kind=link}
| Species | Origin | |||
|---|---|---|---|---|
| European Hedgehog | Algerian Hedgehog | Indigenous | Translocated | |
| N | 14 | 17 | 18 | 13 |
| L | 6.92 ± 0.48 | 7.21 ± 0.66 | 6.67 ± 0.27 | 7.2 ± 0.39 |
| Range | 6–8 | 6–8 | 6–8 | 6–8 |
| Weight (g) | 769.82 ± 223.34 | 545.07 ± 82.51 | 643.61 ± 155.71 | 702.54 ± 263.95 |
| Indigenous | Translocated | Df|n | Z | P | European | Algerian | Df|n | Z | P | |
|---|---|---|---|---|---|---|---|---|---|---|
| PW | 57.9 ± 32.5 | 51.5 ± 42.5 | 1|28 | 0.97 | 0.34 | 54.6 ± 38.4 | 56.3 ± 24.7 | 1|28 | 0.70 | 0.48 |
| HDU | 4.6 ± 13.3 | 11.8 ± 26.4 | 1|28 | 0.67 | 0.5 | 7.6 ± 22.7 | 7.3 ± 15.4 | 1|28 | −1.34 | 0.19 |
| LDU | 25.8 ± 27.8 | 11.1 ± 17.9 | 1|28 | −2.89 | <0.01 | 18.8 ± 23.7 | 21.5 ± 27.6 | 1|28 | 0.69 | 0.49 |
| CU | 1.2 ± 5.2 | 4.8 ± 16.5 | 1|28 | 0.34 | 0.73 | 3.4 ± 13.9 | 1.7 ± 6.2 | 1|28 | −0.16 | 0.86 |
| AC | 6.4 ± 10.7 | 14.9 ± 20.4 | 1|28 | 1.11 | 0.27 | 8.7 ± 12.6 | 11.3 ± 19.3 | 1|28 | −1.58 | 0.12 |
| RV | 3.4 ± 9.2 | 6.0 ± 14.2 | 1|28 | 0.13 | 0.89 | 7.1 ± 14.3 | 1.0 ± 3.5 | 1|28 | 1.22 | 0.23 |
| <700 g | >700 g | Df|n | Z | P | Warm | Cold | Df|n | Z | P | |
| PW | 63.9 ± 35.9 | 40.5 ± 33.3 | 1|28 | 1.75 | 0.09 | 54.7 ± 32.2 | 56.1 ± 42.4 | 1|28 | 1.40 | 0.17 |
| HDU | 3.0 ± 28.4 | 15.1 ± 28.4 | 1|28 | −1.66 | 0.1 | 10.5 ± 23.5 | 3.4 ± 13.3 | 1|28 | 0.08 | 0.93 |
| LDU | 20.2 ± 25.0 | 19.5 ± 25.0 | 1|28 | 0.01 | 0.98 | 15.5 ± 15.9 | 25.7 ± 33.4 | 1|28 | −2.37 | 0.02 |
| CU | 1.2 ± 17.2 | 5.2 ± 17.2 | 1|28 | −0.27 | 0.78 | 3.4 ± 13.9 | 1.7 ± 6.2 | 1|28 | 0.10 | 0.91 |
| AC | 8.9 ± 15.8 | 11.4 ± 15.8 | 1|28 | −1.83 | 0.07 | 12.6 ± 19.2 | 6.1 ± 8.4 | 1|28 | 0.38 | 0.70 |
| RV | 2.2 ± 16.0 | 8.3 ± 16.0 | 1|28 | 0.3 | 0.76 | 3.3 ± 10.6 | 6.0 ± 12.4 | 1|28 | 0.18 | 0.85 |
| Group | Macrohabitat | P | Subhabitat | P |
|---|---|---|---|---|
| Warm | PW–AC | <0.001 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| PW–RV | <0.001 | HED–LEA | <0.001 | |
| PW- LDU | <0.001 | HED–ROO | <0.001 | |
| Cold | PW–AC | <0.001 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| PW–LDU | 0.001 | HED–LEA | <0.001 | |
| PW–RV | <0.001 | HED–ROO | <0.001 | |
| PW–HDU | <0.001 | - | - | |
| LDU–CU | 0.001 | - | - | |
| >700 g | PW–AC | <0.001 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| PW–RV | <0.001 | HED–LEA | <0.001 | |
| PW–HDU | <0.001 | HED–ROO | <0.001 | |
| PW–LDU | 0.006 | - | - | |
| <700 g | PW–AC | <0.001 | LEA–WALL | 0.049 |
| PW–CU | <0.001 | HED–WALL | <0.001 | |
| PW–LDU | <0.001 | HED–RUB | <0.001 | |
| PW–RV | <0.001 | HED–LEA | 0.038 | |
| PW–HDU | <0.001 | HED–ROO | <0.001 | |
| LDU–CU | <0.001 | - | ||
| Native | PW–AC | <0.001 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| PW–RV | <0.001 | HED–LEA | <0.001 | |
| PW–HDU | <0.001 | HED–ROO | <0.001 | |
| PW–LDU | <0.001 | - | - | |
| LDU–AC | 0.021 | - | - | |
| LDU–CU | <0.001 | - | - | |
| LDU–RV | 0.001 | - | - | |
| LDU -HDU | 0.001 | - | - | |
| Released | PW–AC | 0.007 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| PW–RV | <0.001 | HED–LEA | <0.001 | |
| PW–HDU | <0.001 | HED–ROO | <0.001 | |
| PW–LDU | 0.001 | - | - | |
| European | PW–AC | <0.001 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| RV–PW | <0.001 | HED–LEA | <0.001 | |
| PW–HDU | <0.001 | HED–ROO | <0.001 | |
| PW–LDU | <0.001 | - | - | |
| Algerian | PW–AC | <0.001 | HED–WALL | <0.001 |
| PW–CU | <0.001 | HED–RUB | <0.001 | |
| RV–PW | <0.001 | HED–ROO | <0.001 | |
| PW–HDU | <0.001 | LEA–WALL | 0.035 | |
| PW–LDU | <0.001 |
| Indigenous | Translocated | Df|n | Z | P | European | Algerian | Df|n | Z | P | |
|---|---|---|---|---|---|---|---|---|---|---|
| HED | 52.1 ± 23.4 | 58.9 ± 22.8 | 1|28 | 0.11 | 0.91 | 62.9 ± 22.9 | 44.2 ± 19.0 | 1|28 | 6.42 | <0.01 |
| LEA | 20.7 ± 19.6 | 17.8 ± 19.8 | 1|28 | −0.10 | 0.91 | 14.1 ± 18.7 | 26.7 ± 18.5 | 1|28 | −7.43 | <0.01 |
| ROO | 9.3 ± 12.2 | 5.8 ± 14.9 | 1|28 | −1.47 | 0.15 | 5.3 ± 13.2 | 11.2 ± 13.0 | 1|28 | −4.81 | <0.01 |
| RUB | 9.7 ± 18.7 | 5.2 ± 12.1 | 1|28 | 0.74 | 0.46 | 3.7 ± 10.4 | 13.4 ± 21.0 | 1|28 | −7.97 | <0.01 |
| WALL | 7.1 ± 13.9 | 9.1 ± 19.7 | 1|28 | 0.46 | 0.64 | 8.4 ± 18.0 | 7.2 ± 14.2 | 1|28 | −5.23 | 0.54 |
| <700 g | >700 g | Df|n | Z | P | Warm | Cold | Df|n | Z | P | |
| HED | 49.8 ± 22.6 | 63.3 ± 22.1 | 1|28 | 2.02 | 0.09 | 56.2 ± 23.9 | 53.0 ± 22.6 | 1|28 | 0.34 | 0.73 |
| LEA | 24.8 ± 19.5 | 10.5 ± 16.1 | 1|28 | 0.83 | 0.41 | 20.2 ± 19.4 | 18.7 ± 20.2 | 1|28 | −0.04 | 0.96 |
| ROO | 8.7 ± 14.4 | 6.5 ± 11.4 | 1|28 | −0.24 | 0.80 | 4.2 ± 9.5 | 12.7 ± 16.0 | 1|28 | −1.46 | 0.16 |
| RUB | 9.8 ± 18.9 | 4.6 ± 10.5 | 1|28 | 0.70 | 0.48 | 8.8 ± 18.9 | 6.6 ± 12.7 | 1|28 | 0.99 | 0.33 |
| WALL | 4.9 ± 12.1 | 12.9 ± 21.3 | 1|28 | −0.88 | 0.38 | 8.6 ± 18.4 | 6.9 ± 13.4 | 1|28 | 0.25 | 0.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gago, H.; Drechsler, R.M.; Monrós, J.S. Evaluating Different Factors That Affect the Nesting Patterns of European and Algerian Hedgehogs in Urban and Suburban Environments. Animals 2023, 13, 3775. https://doi.org/10.3390/ani13243775
Gago H, Drechsler RM, Monrós JS. Evaluating Different Factors That Affect the Nesting Patterns of European and Algerian Hedgehogs in Urban and Suburban Environments. Animals. 2023; 13(24):3775. https://doi.org/10.3390/ani13243775
Chicago/Turabian StyleGago, Héctor, Robby M. Drechsler, and Juan S. Monrós. 2023. "Evaluating Different Factors That Affect the Nesting Patterns of European and Algerian Hedgehogs in Urban and Suburban Environments" Animals 13, no. 24: 3775. https://doi.org/10.3390/ani13243775