
The Effect of Parity on the Quality of Colostrum of Holstein Dairy Cows in the Organic Production System

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Study Animals
2.2. Data Collection
2.3. Chemical Analyses of Colostrum
2.4. Statistical Analyses
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fox, P.F.; Mc Sweeney, P.L.H. Advanced Dairy Chemistry, 3rd ed.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2003; Volume 1 Proteins. [Google Scholar]
- Kehoe, S.I.; Jayarao, B.M.; Heinrichs, A.J. A survey of bovine colostrum composition and colostrum management practices on Pennsylvania dairy farms. J. Dairy Sci. 2007, 90, 4108–4116. [Google Scholar]
- Puppel, K.; Gołębiewski, M.; Slósarz, J.; Grodkowski, G.; Solarczyk, P.; Kostusiak, P.; Grodkowska, K.; Balcerak, M.; Sakowski, T. Interaction between the level of immunoglobulins and number of somatic cells as a factor shaping the immunomodulating properties of colostrum. Sci. Rep. 2021, 11, 15686. [Google Scholar] [CrossRef]
- Puppel, K.; Gołębiewski, M.; Grodkowski, G.; Slósarz, J.; Kunowska-Slósarz, M.; Solarczyk, P.; Łukasiewicz, M.; Balcerak, M.; Przysucha, T. Composition and Factors Affecting Quality of Bovine Colostrum: A Review. Animals 2019, 9, 1070. [Google Scholar] [CrossRef]
- Buczinski, S.; Vandeweerd, J.M. Diagnostic accuracy of refraktometry for assessing bovine colostrum quality: A systematic review and meta-analysis. J. Dairy Sci. 2016, 99, 7381–7394. [Google Scholar]
- Puppel, K.; Gołębiewski, M.; Grodkowski, G.; Solarczyk, P.; Kostusiak, P.; Klopčič, M.; Sakowski, T. Use of somatic cell count as an indicator of colostrum quality. PLoS ONE 2020, 15, e0237615. [Google Scholar] [CrossRef]
- Lombard, J.; Urie, N.; Garry, F.; Godden, S.; Quigley, J.; Earleywine, T.; McGuirk, S.; Moore, D.; Branan, M.; Chamorro, M.; et al. Consensus recommendations on calf- and herd-level passive immunity in dairy calves in the United States. J. Dairy Sci. 2020, 103, 7611–7624. [Google Scholar]
- Szulc, T.; Zachwieja, A. Siara-eliksir życia osesków. Zesz. Nauk. Akad. Rol. We Wrocławiu Monogr. 1998, 13, 1–98. [Google Scholar]
- Ahmann, J.; Steinhoff-Wagner, J.; Büscher, W. Determining Immunoglobulin Content of Bovine Colostrum and Factors Affecting the Outcome: A Review. Animals 2021, 11, 3587. [Google Scholar] [CrossRef]
- Żychlińska-Buczek, J.; Bauer, E.; Kania-Gierdziewicz, J.; Wrońska, A. The main causes of calf mortality in dairy farms in Poland. J. Agric. Sci. Technol. 2015, 5, 363–369. [Google Scholar]
- Skrzypek, R. Znaczenie odporności siarowej u bydła mlecznego oraz czynniki wpływające na jej status. Biul. Inf. Zootech. 2002, 40, 77–91. [Google Scholar]
- Skrzypczak, W.; Stefaniak, T.; Zabielski, R. Fizjologia Noworodka z Elementami Patofizjologii; Powszechne Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 2011. [Google Scholar]
- Cabral, R.G.; Chapman, C.E.; Aragona, K.M.; Clark, E.; Lunak, M.; Erickson, P.S. Predicting colostrum quality from performance in the previous lactation and environmental changes. J. Dairy Sci. 2016, 99, 4048–4055. [Google Scholar]
- IBM Corp. Released 2022: IBM SPSS Statistics for Windows, Version 22.0; IBM Corp.: Armonk, NY, USA, 2022. [Google Scholar]
- Quigley, J.D., III.; Martin, K.; Dowlen, H.; Wallis, L.; Lamar, K. Immunoglobulin concentration, specific gravity, and nitrogen fractions of colostrum from Jersey cattle. J. Dairy Sci. 1994, 77, 264–269. [Google Scholar]
- Król, J.; Litwińczuk, Z.; Brodziak, A.; Karasińska, A. Zawartość immunoglobulin G w mleku w zależności od rasy i wieku krów oraz fazy laktacji. Med. Wet. 2014, 70, 237–241. [Google Scholar]
- Coleman, L.; Hickson, R.; Amoore, J.; Laven, R.; Back, P. Colostral immunoglobulin G as a predictor for serum immunoglobulin G concentration in dairy calves. In Proceedings of the New Zealand Society of Animal Production; New Zealand Society of Animal Production: Auckland, New Zealand, 2015. [Google Scholar]
- Gulliksen, S.; Lie, K.; Sølverød, L.; Østerås, O. Risk factors associated with colostrum quality in Norwegian dairy cows. J. Dairy Sci. 2008, 91, 704–712. [Google Scholar]
- Dunn, A.; Ashfield, A.; Earley, B.; Welsh, M.; Gordon, A.; Morrison, S. Evaluation of factors associated with immunoglobulin G, fat, protein, and lactose concentrations in bovine colostrum and colostrum management practices in grassland-based dairy systems in Northern Ireland. J. Dairy Sci. 2017, 100, 2068–2079. [Google Scholar]
- Genc, M.; Coban, O. Effect of Some Environmental Factors on Colostrum Quality and Passive Immunity in Brown Swiss and Holstein Cattle. Ph.D. Thesis, Atatürk University, Erzurum, Turkey, 2015. [Google Scholar]
- Abdullahoğlu, E.; Duru, S.; Özlüer, A.; Filya, İ. Factors affecting colostrum quality and calf passive transfer levels in Holstein cattle. Anim. Sci. Pap. Rep. 2019, 37, 29–39. [Google Scholar]
- Nowak, W.; Mikuła, R.; Kasprowicz-Potocka, M.; Ignatowicz, M.; Zachwieja, A.; Paczyńska, K.; Pecka, E. Effect of cow nutrition in the far-off period on colostrum quality and immune response of calves. J. Vet. Res. 2012, 56, 241–246. [Google Scholar]
- Mann, S.; Yepes, F.L.; Overton, T.; Lock, A.; Lamb, S.; Wakshlag, J.; Nydam, D. Effect of dry period dietary energy level in dairy cattle on volume, concentrations of immunoglobulin G, insulin, and fatty acid composition of colostrum. J. Dairy Sci. 2016, 99, 1515–1526. [Google Scholar]
- Bar, E.; Tiris, I.; Sarbu, D.; Iridon, C.; Ochea, I.; Bratu, I. Full characterization of bovine colostrum, raw material for dietary supplements. His beneficial effect on the human immune system. AUCFT 2010, 14, 33–40. [Google Scholar]
- Król, J.; Litwińczuk, Z.; Brodziak, A.; Barłowska, J. Lactoferrin, lysozyme and immunoglobulin G content in milk of four breeds of cows managed under intensive production system. Pol. J. Vet. Sci. 2010, 13, 357–361. [Google Scholar]
- Soufleri, A.; Banos, G.; Panousis, N.; Fletouris, D.; Arsenos, G.; Kougioumtzis, A.; Valergakis, G.E. Evaluation of Factors Affecting Colostrum Quality and Quantity in Holstein Dairy Cattle. Animals 2021, 11, 2005. [Google Scholar]
- Zachwieja, A. Wpływ wieku krów na jakość siary i poziom białek w surowicy krwi ich cieląt. Med. Wet. 1991, 47, 270–271. [Google Scholar]
- Pawlak, H. Dlaczego siara jest tak ważna dla cielęcia? Porad. Gospod. 1995, 10, 14–15. [Google Scholar]
- Lach, Z. Pierwszy łyk życia. Hod. Chów Bydła 2009, 9, 18–19. [Google Scholar]
- Wasowska, E.; Puppel, K. Changes in the content of immunostimulating components of colostrum obtained from dairy cows at different levels of production. J. Sci. Food Agric. 2018, 98, 5062–5068. [Google Scholar]
- Morrill, K.M.; Conrad, E.; Lago, A.; Campbell, J.; Quigley, J.; Tyler, H. Nationwide evaluation of quality and composition of colostrum on dairy farms in the United States. J. Dairy Sci. 2012, 95, 3997–4005. [Google Scholar]
- Quinn, E.M.; O’Callaghan, T.F.; Tobin, J.T.; Murphy, J.P.; Sugrue, K.; Slattery, H. Changes to the oligosaccharide profile of bovine milk at the onset of lactation. Dairy 2020, 1, 284–296. [Google Scholar]
- Zarei, S.; Ghorbani, G.R.; Khorvash, M.; Martin, O.B.; Mahdavi, A.H.; Riasi, A. The impact of season, parity, and volume of colostrum on Holstein dairy cows colostrum composition. Agric. Sci. 2017, 8, 572–581. [Google Scholar]
- Kessler, E.C.; Bruckmaier, R.M.; Gross, J.J. Colostrum composition and immunoglobulin G content in dairy and dual-purpose cattle breeds. J. Anim. Sci. 2020, 98, skaa237. [Google Scholar] [CrossRef]
- Aydogdu, U.; Guzelbektes, H. Effect of colostrum composition on passive calf immunity in primiparous and multiparous dairy cows. Vet. Medic. 2018, 63, 1–11. [Google Scholar]
- Jolazadeh, A.R.; Mohammadabadi, T.; Dehghan-Banadaky, M.; Chaji, M.; Garcia, M. Effect of supplementing calcium salts of n-3 and n-6 fatty acid to pregnant nonlactating cows on colostrum composition, milk yield, and reproductive performance of dairy cows. Anim. Feed Sci. Technol. 2019, 247, 127–140. [Google Scholar]
- Garcia, M.; Greco, L.F.; Favoreto, M.G.; Marsola, R.S.; Martins, L.T.; Bisinotto, R.S. Effect of supplementing fat to pregnant nonlactating cows on colostral fatty acid profile and passive immunity of the newborn calf. J. Dairy Sci. 2014, 97, 392–405. [Google Scholar]
- O'Callaghan, T.F.; O'Donovan, M.; Murphy, J.P.; Sugrue, K.; Mannion, D.; McCarthy, W.P. Evolution of the bovine milk fatty acid profile–From colostrum to milk five days post parturition. Int. Dairy J. 2020, 104, 104655. [Google Scholar] [CrossRef]
- Demkowicz, M. Bioaktywne preparat z siary zachowujące parametry jej wartości biologicznej oraz ich przyswajalność u jagniąt i cieląt. Ph.D. Thesis, Uniwersytet Przyrodniczy we Wrocławiu, Wrocław, Poland, 2012. [Google Scholar]
- Guliński, P.; Niedziałek, G.; Salamończyk, E.; Górski, T. Zawartość immunoglobulin w siarze krów w zależności od wybranych czynników genetycznych i środowiskowych. Med. Wet. 2006, 62, 339–342. [Google Scholar]
- Debier, C.; Pottier, J.; Goffe, C.; Larondelle, Y. Present knowledge and unexpected behaviors of vitamins A and E in colostrum and milk. Livest. Prod. Sci. 2005, 98, 135–147. [Google Scholar]
- Kowalski, Z.M. Siara to nie tylko immunoglobuliny. Hoduj Głową Bydło 2010, 4, 14–16. [Google Scholar]
- Foley, J.A.; Otterby, D.E. Availability, storage, treatment, composition, and feeding value of surplus colostrum: A review. J. Dairy Sci. 1978, 61, 1033–1060. [Google Scholar]
- Hammon, H.M.; Zanker, I.A.; Blum, J.W. Delayed colostrum feeding affects IGF-I and insulin plasma concentrations in neonatal calves. J. Dairy Sci. 2000, 83, 85–92. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Cows | |
|---|---|---|
| Close-Up | Fresh | |
| Feed intake and energy balance | ||
| Conserved forages (kg DM/d) | 11.5 | 12.5 |
| Concentrate (kg DM/d) | 1.9 | 4.5 |
| Total (kg DM/d) | 13.4 | 17.0 |
| NEL intake (MJ NEL/d) | 74.3 | 84.9 |
| EB (MJ NEL/d) | 18.7 | 30.1 |
| nXP intake (g/d) | 2232 | 2658 |
| BW (kg) | 675 | 659 |
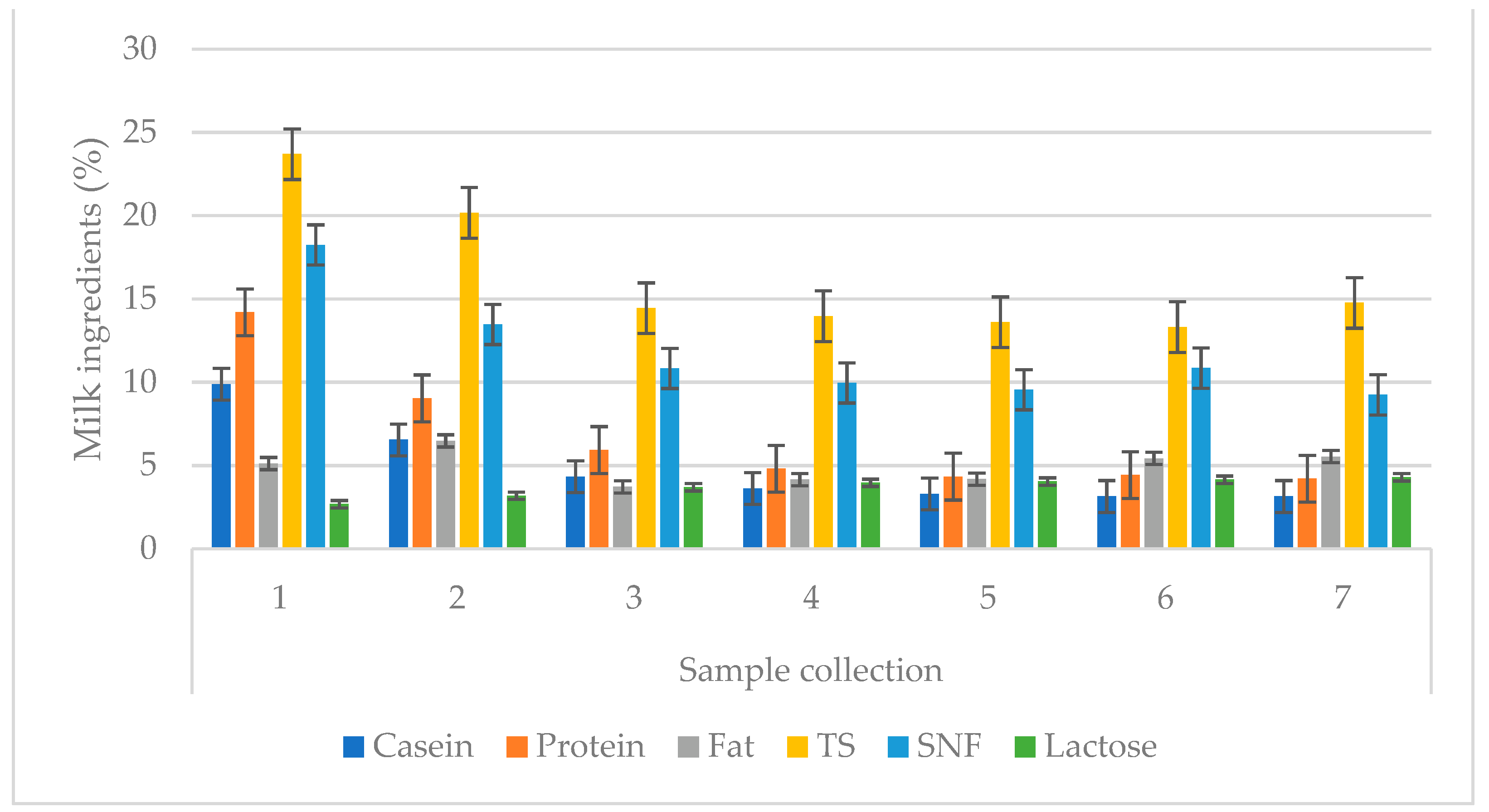
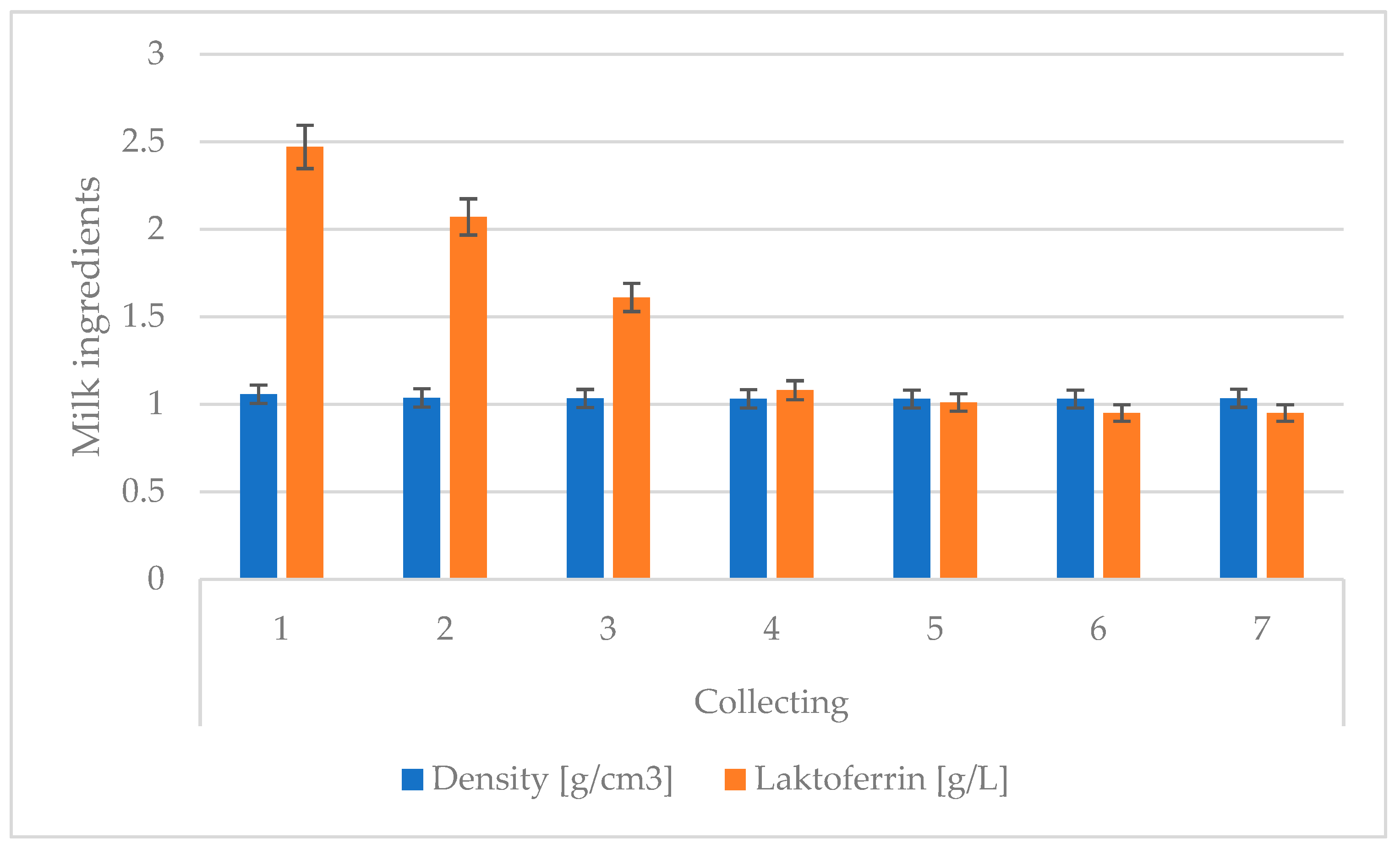
| Sample Collection | Lactation | Ingredient | |
|---|---|---|---|
| Density [g/cm3] | Lactoferrin [g/L] | ||
| 1 | 1 | 1.037 ABC | 1.69 ABC |
| 2 | 1.056 ADE | 3.32 ADE | |
| 4 | 1.049 BDf | 2.05 BDF | |
| 5 | 1.051 CEf | 3.20 CEF | |
| 2 | 1 | 1.029 ABC | 0.92 ABC |
| 2 | 1.039 ADE | 3.03 ADE | |
| 4 | 1.040 BDF | 2.68 BDF | |
| 5 | 1.036 CEF | 1.25 CEF | |
| 3 | 1 | 1.031 A | 0.66 ABC |
| 2 | 1.042 ABC | 2.40 ADE | |
| 4 | 1.031 B | 2.06 BDF | |
| 5 | 1.031 C | 1.03 CEF | |
| 4 | 1 | 1.032 a | 0.62 ABC |
| 2 | 1.036 abc | 2.10 ADE | |
| 4 | 1.032 b | 0.69 BDF | |
| 5 | 1.031 c | 0.79 CEF | |
| 5 | 1 | 1.031 a | 0.65 AB |
| 2 | 1.035 abc | 1.90 ACD | |
| 4 | 1.030 b | 0.63 CE | |
| 5 | 1.029 c | 0.72 BDE | |
| 6 | 1 | 1.031 ab | 0.58 AB |
| 2 | 1.028 ac | 1.83 ACD | |
| 4 | 1.029 b | 0.58 CE | |
| 5 | 1.030 C | 0.65 BDE | |
| 7 | 1 | 1.023 ABC | 0.56 AbC |
| 2 | 1.031 A | 1.90 ADE | |
| 4 | 1.032 B | 0.52 BDF | |
| 5 | 1.031 C | 0.73 CEF | |
| Sample Collection | Lactation | Whey Protein | ||
|---|---|---|---|---|
| α-Lactalbumin [g/L] | β-Lactoglobulin [g/L] | Immunoglobulin G [g/L] | ||
| 1 | 1 | 1.24 ABC | 5.27 ABC | 26.50 ABC |
| 2 | 3.29 Ade | 6.19 AD | 79.43 ADE | |
| 4 | 3.71 Bd | 6.20 BE | 38.14 BDF | |
| 5 | 3.52 Ce | 8.83 CDE | 69.32 CEF | |
| 2 | 1 | 0.94 ABC | 2.37 ABC | 14.37 ABC |
| 2 | 2.85 ADE | 4.94 A | 19.51 ADE | |
| 4 | 1.79 BDF | 4.92 B | 22.55 BDF | |
| 5 | 3.12 CEF | 4.58 C | 8.64 CEF | |
| 3 | 1 | 0.75 ABC | 3.08 ABC | 6.86 ABC |
| 2 | 2.30 Ade | 3.97 A | 11.70 AD | |
| 4 | 1.94 BdF | 3.92 B | 11.83 BE | |
| 5 | 2.57 CeF | 3.78 C | 3.95 CDE | |
| 4 | 1 | 0.69 ABC | 2.80 AB | 5.65 ABC |
| 2 | 2.14 ACd | 3.63 AC | 6.30 ADE | |
| 4 | 0.77 CE | 3.19 BD | 8.84 BDF | |
| 5 | 1.92 BdE | 2.83 CD | 4.50 CEF | |
| 5 | 1 | 0.75 AB | 3.17 A | 5.14 ABC |
| 2 | 1.92 ACD | 3.24 B | 3.81 ADE | |
| 4 | 0.72 CE | 3.01 C | 7.46 BDF | |
| 5 | 1.79 BDE | 2.63 ABC | 4.47 CEF | |
| 6 | 1 | 0.66 AB | 2.82 abC | 4.68 ABC |
| 2 | 1.77 ACD | 3.05 adE | 16.89 ADE | |
| 4 | 0.72 CE | 2.73 bd | 5.79 BDF | |
| 5 | 1.63 BDE | 2.40 CE | 3.81 CEF | |
| 7 | 1 | 0.63 AB | 2.72 Abc | 11.12 ABC |
| 2 | 2.00 ACd | 3.37 ADE | 1.97 ADE | |
| 4 | 0.59 CE | 2.56 bDf | 5.63 BDF | |
| 5 | 1.82 BdE | 2.67 cEf | 3.57 CEF | |
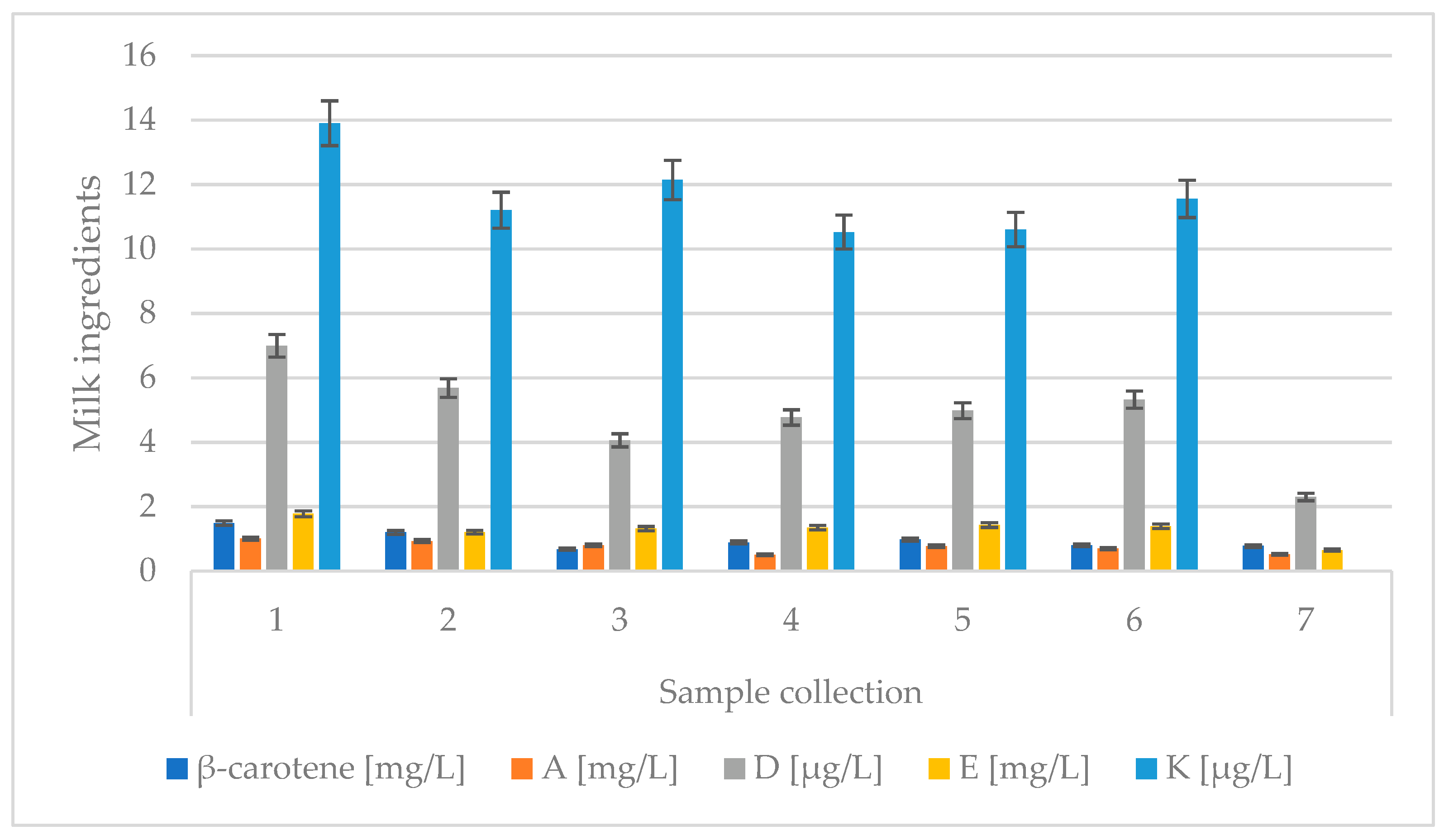
| Sample Collection | Lactation | Vitamin | ||||
|---|---|---|---|---|---|---|
| β-Carotene [mg/L] | A [mg/L] | D [µg/L] | E [mg/L] | K [µg/L] | ||
| 1 | 1 | 1.19 ABC | 0.74 ABC | 5.90 ABC | 1.62 Abc | 14.78 AB |
| 2 | 1.71 ADE | 1.33 ADF | 6.52 ADE | 2.77 ADE | 14.18 CD | |
| 4 | 1.34 BDF | 1.00 BDg | 8.04 BDF | 1.14 bD | 12.87 ACF | |
| 5 | 1.99 CEF | 0.94 CFg | 7.99 CEF | 1.17 cE | 13.56 BDF | |
| 2 | 1 | 0.97 abC | 0.54 ABC | 5.48 ABC | 1.07 a | 11.04 ABC |
| 2 | 1.19 aD | 1.34 ADF | 3.41 ADE | 1.55 abc | 9.51 AD | |
| 4 | 1.16 bE | 0.90 BD | 5.68 BDF | 1.05 b | 13.67 BDE | |
| 5 | 1.80 CDE | 0.93 CF | 10.37 CEF | 1.15 c | 9.97 CE | |
| 3 | 1 | 0.61 ABC | 0.47 ABC | 3.02 ABC | 1.08 A | 7.21 ABC |
| 2 | 1.43 ADE | 1.34 ADE | 8.02 ADE | 2.30 ABC | 11.98 ADE | |
| 4 | 0.13 BDF | 0.58 BDF | 1.30 BDF | 0.83 B | 16.75 BDF | |
| 5 | 0.40 CEF | 0.90 CEF | 3.75 CEF | 0.80 C | 13.09 CEF | |
| 4 | 1 | 0.60 BCb | 0.49 ABC | 4.45 ABC | 0.68 A | 5.98 ABC |
| 2 | 0.72 aBeF | 0.66 ADE | 1.74 ADE | 0.62 BC | 15.66 ADE | |
| 4 | 0.82 CeG | 0.39 BDF | 8.89 BDF | 1.29 ABD | 6.78 BDF | |
| 5 | 1.67 DFG | 0.50 CEF | 3.30 CEF | 2.29 CD | 16.83 CEF | |
| 5 | 1 | 0.43 ABC | 0.58 ABC | 0.99 ABC | 0.73 ABc | 11.84 AB |
| 2 | 0.90 ADE | 0.82 ADE | 2.31 ADE | 1.04 ADE | 11.97 CD | |
| 4 | 1.51 BDF | 0.97 BDF | 12.58 BDF | 2.77 BDF | 9.88 ACE | |
| 5 | 1.20 CEF | 0.70 CEF | 3.09 CEF | 0.94 cEF | 6.87 BDE | |
| 6 | 1 | 0.35 ABC | 0.47 AbC | 1.25 ABC | 0.71 ABc | 9.18 ABC |
| 2 | 1.27 ADE | 0.87 ADE | 9.14 ADE | 2.10 ADE | 14.71 ADE | |
| 4 | 0.99 BDF | 0.51 bDF | 6.47 BDF | 1.68 BDF | 10.95 BDF | |
| 5 | 0.42 CEF | 1.15 CEF | 3.50 CEF | 0.77 cEF | 11.19 CEF | |
| 7 | 1 | 0.58 AB | 0.42 ABC | 2.04 ABC | 0.94 aBC | 5.84 ABC |
| 2 | 0.88 ACD | 0.74 ADE | 1.98 ADE | 0.68 ade | 11.16 ADE | |
| 4 | 0.57 CE | 0.39 BDF | 1.74 BD | 0.48 Bd | 10.13 BDF | |
| 5 | 1.16 BDE | 0.53 CEF | 4.68 CE | 0.40 Ce | 6.86 CEF | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grodkowska, K.; Gołębiewski, M.; Slósarz, J.; Grodkowski, G.; Kostusiak, P.; Sakowski, T.; Klopčič, M.; Puppel, K. The Effect of Parity on the Quality of Colostrum of Holstein Dairy Cows in the Organic Production System. Animals 2023, 13, 540. https://doi.org/10.3390/ani13030540
Grodkowska K, Gołębiewski M, Slósarz J, Grodkowski G, Kostusiak P, Sakowski T, Klopčič M, Puppel K. The Effect of Parity on the Quality of Colostrum of Holstein Dairy Cows in the Organic Production System. Animals. 2023; 13(3):540. https://doi.org/10.3390/ani13030540
Chicago/Turabian StyleGrodkowska, Kinga, Marcin Gołębiewski, Jan Slósarz, Grzegorz Grodkowski, Piotr Kostusiak, Tomasz Sakowski, Marija Klopčič, and Kamila Puppel. 2023. "The Effect of Parity on the Quality of Colostrum of Holstein Dairy Cows in the Organic Production System" Animals 13, no. 3: 540. https://doi.org/10.3390/ani13030540
APA StyleGrodkowska, K., Gołębiewski, M., Slósarz, J., Grodkowski, G., Kostusiak, P., Sakowski, T., Klopčič, M., & Puppel, K. (2023). The Effect of Parity on the Quality of Colostrum of Holstein Dairy Cows in the Organic Production System. Animals, 13(3), 540. https://doi.org/10.3390/ani13030540