
Sow Nutrition, Uterine Contractions, and Placental Blood Flow during the Peri-Partum Period and Short-Term Effects on Offspring: A Review

Abstract
:Simple Summary
Abstract
1. Introduction
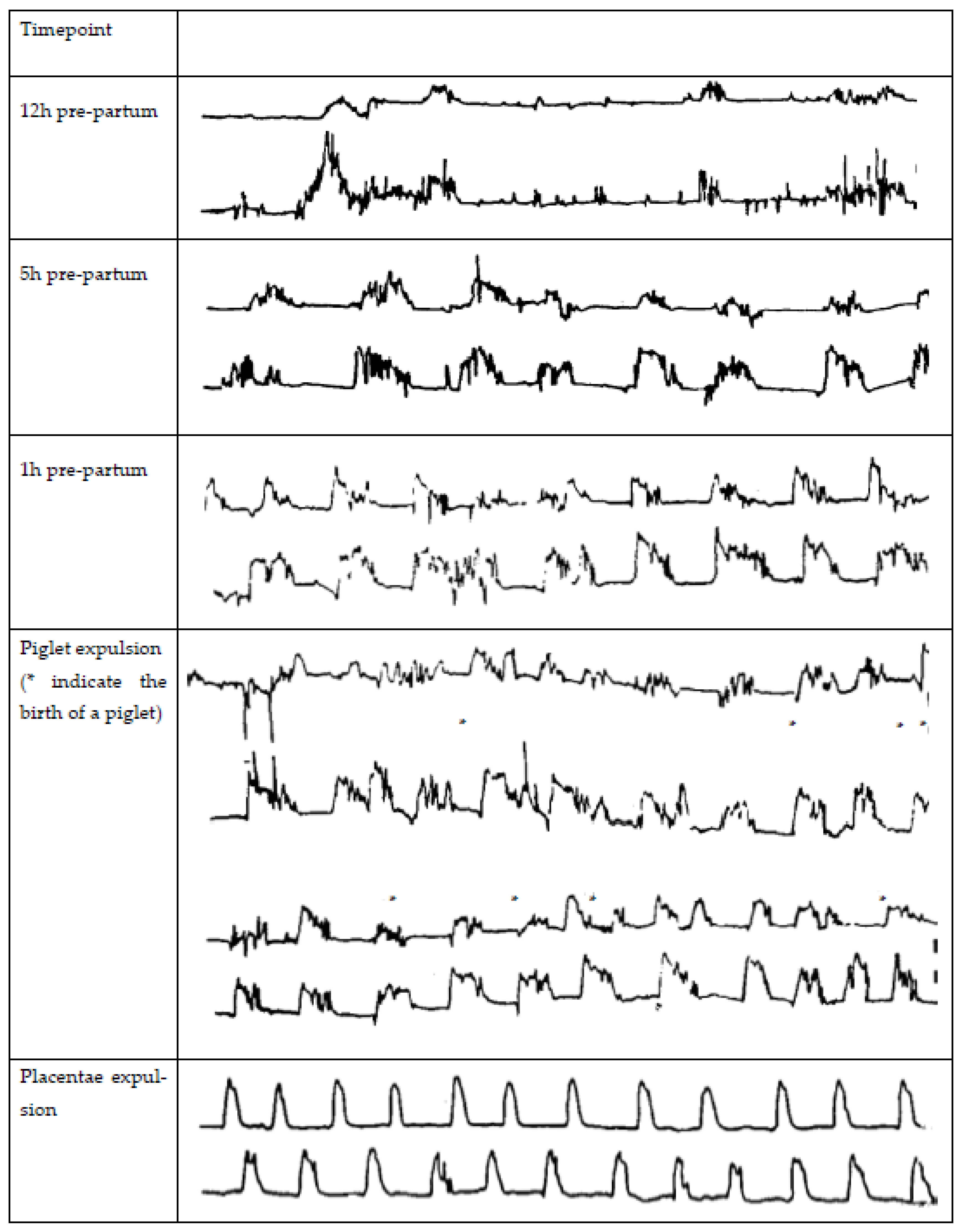
2. Uterine Contractions
3. Uterine Blood Flow
4. Placental and Umbilical Cord Functionality
5. The Potential of Maternal Nutrition to Reduce Farrowing Duration
5.1. Energy
5.2. Glucose as a Source of Energy during Farrowing
5.3. Other Carbohydrates
5.4. Calcium and Magnesium
5.5. Vasoactive Components
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rutherford, K.M.D.; Baxter, E.M.; D’Eath, R.B.; Turner, S.P.; Arnott, G.; Roehe, R.; Ask, B.; Sandøe, P.; Moustsen, V.A.; Thorup, F.; et al. The welfare implications of large litter size in the domestic pig I: Biological factors. Anim. Welf. 2013, 22, 199–218. [Google Scholar] [CrossRef] [Green Version]
- Rootwelt, V.; Reksen, O.; Farstad, W.; Framstad, T. Postpartum deaths: Piglet, placental, and umbilical characteristics. J. Anim. Sci. 2013, 91, 2647–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Algers, B.; Uvnäs-Moberg, K. Maternal behavior in pigs. Horm. Behav. 2007, 52, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, O.; Björkman, S.; Oliviero, C. Parturition effects on reproductive health in the gilt and sow. Reprod. Domest. Anim. 2016, 51, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosnier, E.; Etienne, M.; Ramaekers, P.; Père, M.C. The metabolic status during the peri partum period affects the voluntary feed intake and the metabolism of the lactating multiparous sow. Livest. Sci. 2010, 127, 127–136. [Google Scholar] [CrossRef]
- Pere, M.C.; Etienne, M. Uterine blood flow in sows: Effects of pregnancy stage and litter size. Reprod. Nutr. Dev. 2000, 40, 369–382. [Google Scholar] [CrossRef] [Green Version]
- Boulot, S.; Quesnel, H.; Quiniou, N. Management of high prolificacy in French herds: Can we alleviate side effects on piglet survival. Adv. Pork Prod. 2008, 19, 213–220. [Google Scholar]
- Quiniou, N.; Dagorn, J.; Gaudré, D. Variation of piglets’ birth weight and consequences on subsequent performance. Livest. Prod. Sci. 2002, 78, 63–70. [Google Scholar] [CrossRef]
- Rootwelt, V.; Reksen, O.; Farstad, W.; Framstad, T. Associations between intrapartum death and piglet, placental, and umbilical characteristics. J. Anim. Sci. 2012, 90, 4289–4296. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.W.; Bazer, F.W.; Thatcher, W.W.; Franke, D.E.; Wallace, H.D. Conceptus development in intact and unilaterally hysterectomized-ovariectomized gilts: Interrelations among hormonal status, placental development, fetal fluids and fetal growth. J. Anim. Sci. 1977, 44, 620–637. [Google Scholar] [CrossRef]
- Wilson, M.E.; Biensen, N.J.; Ford, S.P. Novel insight into the control of litter size in pigs, using placental efficiency as a selection tool. J. Anim. Sci. 1999, 77, 1654–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rens, B.T.T.M.; Van Der Lende, T. Parturition in gilts: Duration of farrowing, birth intervals and placenta expulsion in relation to maternal, piglet and placental traits. Theriogenology 2004, 62, 331–352. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, C.; Heinonen, M.; Valros, A.; Peltoniemi, O. Environmental and sow-related factors affecting the duration of farrowing. Anim. Reprod. Sci. 2010, 119, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, A.J.; Van Rens, B.T.T.M.; Van Der Lende, T.; Taverne, M.A.M. Factors affecting duration of the expulsive stage of parturition and piglet birth intervals in sows with uncomplicated, spontaneous farrowings. Theriogenology 2005, 64, 1573–1590. [Google Scholar] [CrossRef] [PubMed]
- Maffeo, G.; Vigo, D.; Ballabio, R.; Olivia, O.; Cairoli, F.; Jöchle, W. Uterine Motility in Sows During Spontaneous Parturition and Induced Parturitions with the PGF Analog Alfbprostol and Oxytocin. Reprod. Domest. Anim. 2007, 25, 36–43. [Google Scholar] [CrossRef]
- van Dijk, A.J.; van Loon, J.P.A.M.; Taverne, M.A.M.; Jonker, F.H. Umbilical cord clamping in term piglets: A useful model to study perinatal asphyxia? Theriogenology 2008, 70, 662–674. [Google Scholar] [CrossRef] [Green Version]
- Feyera, T.; Pedersen, T.F.; Krogh, U.; Foldager, L.; Theil, P.K. Impact of sow energy status during farrowing on farrowing kinetics, frequency of stillborn piglets, and farrowing assistance. J. Anim. Sci. 2018, 96, 2320–2331. [Google Scholar] [CrossRef] [Green Version]
- Guillemet, R.; Hamard, A.; Quesnel, H.; Père, M.C.; Etienne, M.; Dourmad, J.Y.; Meunier-Salaün, M.C. Dietary fibre for gestating sows: Effects on parturition progress, behaviour, litter and sow performance. Animal 2007, 1, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Le Cozler, Y.; Beaumal, V.; Neil, M.; David, C.; Dourmad, J.Y. Changes in the concentrations of glucose, non-esterifed fatty acids, urea, insulin, cortisol and some mineral elements in the plasma of the primiparous sow before, during and after induced parturition. Reprod. Nutr. Dev. 1999, 39, 161–169. [Google Scholar] [CrossRef]
- Quiniou, N. Effect of feed allowance during late gestation on farrowing progress, piglets’ vitality and lactation performance. J. Rech. Porc. 2005, 37, 187–194. [Google Scholar]
- Tydlitát, D.; Vinkler, A.; Czanderlová, L. Influence of crude protein intake on the duration of delivery and litter size in sows. Acta Vet. Brno 2008, 77, 25–30. [Google Scholar] [CrossRef]
- Vallet, J.L.; Miles, J.R.; Rempel, L.A. Effect of creatine supplementation during the last week of gestation on birth intervals, stillbirth, and preweaning mortality in pigs. J. Anim. Sci. 2013, 91, 2122–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.E.T. Observations on Parturition in the Sow: Part II: The Parturient and Post-Parturient Phases. Br. Vet. J. 1966, 122, 471–478. [Google Scholar] [CrossRef]
- Ellendorff, F.; Taverne, M.; Elsaesser, F.; Forsling, M.; Parvizi, N.; Naaktgeboren, C.; Smidt, D. Endocrinology of parturition in the pig. Anim. Reprod. Sci. 1979, 2, 323–334. [Google Scholar] [CrossRef]
- Peltoniemi, O.; Oliviero, C. Housing, management and environment during farrowing and early lactation. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 231–247. [Google Scholar]
- Purohit, G. Parturition in domestic animals: A review. WebmedCentral Reprod. 2010, 1, WMC00748. [Google Scholar]
- Walls, A.; Hatze, B.; Lomax, S.; Bathgate, R. Defining "Normal" in Pig Parturition. Animals 2022, 12, 2754. [Google Scholar] [CrossRef] [PubMed]
- King, G.; Wathes, D. Relaxin, progesterone and estrogen profiles in sow plasma during natural and induced parturitions. Anim. Reprod. Sci. 1989, 20, 213–220. [Google Scholar] [CrossRef]
- Decaluwe, R.; Janssens, G.; Declerck, I.; de Kruif, A.; Maes, D. Induction of parturition in the sow. Vlaams Diergeneeskd. Tijdschr. 2012, 81, 158–165. [Google Scholar] [CrossRef]
- Ivell, R.; Kimura, T.; Müller, D.; Augustin, K.; Abend, N.; Bathgate, R.; Telgmann, R.; Balvers, M.; Tillmann, G.; Fuchs, A.-R. The structure and regulation of the oxytocin receptor. Exp. Physiol. 2001, 86, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Taverne, M.A.M. Myometrial activity during pregnancy and parturition in the pig. Control. Pig Reprod. I 1982, 11, 419–436. [Google Scholar] [CrossRef]
- Senger, P.L. Placentation, the Endocrinology of the gestation and Parturition. In Pathways to Pregnancy and Parturition; Current Conceptions Inc.: Pullman, WA, USA, 2003; pp. 304–325. [Google Scholar]
- Vallet, J.L.; Miles, J.R.; Brown-Brandl, T.M.; Nienaber, J.A. Proportion of the litter farrowed, litter size, and progesterone and estradiol effects on piglet birth intervals and stillbirths. Anim. Reprod. Sci. 2010, 119, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taverne, M.; Naaktgeboren, C.; Elsaesser, F.; Forsling, M.; Van der Weyden, G.; Ellendorff, F.; Smidt, D. Myometrial electrical activity and plasma concentrations of progesterone, estrogens and oxytocin during late pregnancy and parturition in the miniature pig. Biol. Reprod. 1979, 21, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Zerobin, K. Untersuchungen über die Uterusmotorik des Schweines. Zent. Für Veterinärmedizin Reihe A 1968, 15, 777–798. [Google Scholar] [CrossRef]
- Kuwabara, Y.; Takeda, S.; Mizuno, M.; Sakamoto, S. Oxytocin levels in maternal and fetal plasma, amniotic fluid, and neonatal plasma and urine. Arch. Gynecol. Obstet. 1987, 241, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Theil, P.K.; Farmer, C.; Feyera, T. Physiology and nutrition of late gestating and transition sows. J. Anim. Sci. 2022, 100, skac176. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Martínez-Burnes, J.; Trujillo, M.E.; Lopez, A.; Rosales, A.M.; Ramírez, R.; Orozco, H.; Merino, A.; Alonso-Spilsbury, M. Uterine and fetal asphyxia monitoring in parturient sows treated with oxytocin. Anim. Reprod. Sci. 2005, 86, 131–141. [Google Scholar] [CrossRef]
- Randall, G.C. Observations on parturition in the sow. II. Factors influencing stillbirth and perinatal mortality. Vet. Rec. 1972, 90, 183–186. [Google Scholar] [CrossRef]
- Tucker, J.M.; Hauth, J.C. Intrapartum Assessment of Fetal Well-Being. Clin. Obstet. Gynecol. 1990, 33, 515–525. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Nava-Ocampo, A.A.; Trujillo, M.E.; Velázquez-Armenta, Y.; Ramírez-Necoechea, R.; Martínez-Burnes, J. Dose minimization study of oxytocin in early labor in sows: Uterine activity and fetal outcome. Reprod. Toxicol. 2005, 20, 255–259. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Villanueva-García, D.; Velazquez-Armenta, E.Y.; Nava-Ocampo, A.A.; Ramírez-Necoechea, R.; Alonso-Spilsbury, M.; Trujillo, M.E. Influence of time at which oxytocin is administered during labor on uterine activity and perinatal death in pigs. Biol. Res. 2007, 40, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Leibrecht, R. Über die Struktur der Ringmuskelschicht am Uterus von Rind und Schwein und ihre funktionelle Bedeutung; Ludwig-Maximilians Universität München: München, Germany, 1953. [Google Scholar]
- Zaleski, H.M.; Hacker, R.R. Effect of oxygen and neostigmine on stillbirth and pig viability. J. Anim. Sci. 1993, 71, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Vanderhaeghe, C.; Dewulf, J.; de Kruif, A.; Maes, D. Non-infectious factors associated with stillbirth in pigs: A review. Anim. Reprod. Sci. 2013, 139, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Björkman, S.; Oliviero, C.; Rajala-Schultz, P.J.; Soede, N.M.; Peltoniemi, O.A.T. The effect of litter size, parity and farrowing duration on placenta expulsion and retention in sows. Theriogenology 2017, 92, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Hales, J.; Moustsen, V.A.; Devreese, A.M.; Nielsen, M.B.F.; Hansen, C.F. Comparable farrowing progress in confined and loose housed hyper-prolific sows. Livest. Sci. 2015, 171, 64–72. [Google Scholar] [CrossRef]
- Oliviero, C.; Kothe, S.; Heinonen, M.; Valros, A.; Peltoniemi, O. Prolonged duration of farrowing is associated with subsequent decreased fertility in sows. Theriogenology 2013, 79, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Langendijk, P.; Plush, K. Parturition and its relationship with stillbirths and asphyxiated piglets. Animals 2019, 9, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, G.C.B. Perinatal adaptation in animals. Anim. Reprod. Sci. 1992, 28, 309–318. [Google Scholar] [CrossRef]
- Randall, G. Studies on the effect of acute asphyxia on the fetal pig in utero. Neonatology 1979, 36, 63–69. [Google Scholar] [CrossRef]
- Singer, D. Neonatal tolerance to hypoxia: A comparative-physiological approach. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1999, 123, 221–234. [Google Scholar] [CrossRef]
- Pigozzi, F.; Giombini, A.; Fagnani, F.; Parisi, A. CHAPTER 3 - The Role of Diet and Nutritional Supplements. In Clinical Sports Medicine; Frontera, W.R., Herring, S.A., Micheli, L.J., Silver, J.K., Young, T.P., Eds.; W.B. Saunders: Edinburgh, Scotland, 2007; pp. 23–36. [Google Scholar]
- Omo-Aghoja, L. Maternal and fetal Acid-base chemistry: A major determinant of perinatal outcome. Ann. Med. Health Sci. Res. 2014, 4, 8–17. [Google Scholar] [CrossRef] [Green Version]
- English, P.R.; Wilkinson, V. Management of the sow and litter in late pregnancy and lactation in relation to piglet survival and growth. In Control Pig Reproduction; Cole, D.J.A., Foxcroft, G.R., Eds.; Butterworths: Lond, UK, 1982; pp. 479–506. [Google Scholar]
- Langendijk, P.; Fleuren, M.; van Hees, H.; van Kempen, T. The course of parturition affects piglet condition at birth and survival and growth through the nursery phase. Animals 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langendijk, P.; Fleuren, M.; van Kempen, T.A. Birth interval or duration of parturition: Which is relevant to risk of stillbirth and intervention? In Proceedings of the 69th Annual Meeting of the European Federation of Animal Science, Dubrovnik, Croatia, 27–31 August 2018; p. 112. [Google Scholar]
- Oliviero, C.; Kokkonen, T.; Heinonen, M.; Sankari, S.; Peltoniemi, O. Feeding sows with high fibre diet around farrowing and early lactation: Impact on intestinal activity, energy balance related parameters and litter performance. Res. Vet. Sci. 2009, 86, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Theil, P.T. Transition feeding of sows. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publisher: Wageningen, The Netherlands, 2015; pp. 147–172. [Google Scholar]
- Thorsen, C.K.; Schild, S.-L.A.; Rangstrup-Christensen, L.; Bilde, T.; Pedersen, L.J. The effect of farrowing duration on maternal behavior of hyperprolific sows in organic outdoor production. Livest. Sci. 2017, 204, 92–97. [Google Scholar] [CrossRef]
- Bosch, v.d.M.; Wijnen, H.J.; Linde, v.d.I.B.; Wesel, v.A.A.M.; Melchior, D.; Kemp, B.; Brand, v.d.H.; Clouard, C.M. Effects of maternal dietary nitrate supplementation on farrowing and placental characteristics, level of asphyxiation at birth and piglet vitality. Theriogenology 2019, 129, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Udomchanya, J.; Suwannutsiri, A.; Sripantabut, K.; Pruchayakul, P.; Juthamanee, P.; Nuntapaitoon, M.; Tummaruk, P. Association between the incidence of stillbirths and expulsion interval, piglet birth weight, litter size and carbetocin administration in hyper-prolific sows. Livest. Sci. 2019, 227, 128–134. [Google Scholar] [CrossRef]
- Gourley, K.M.; Swanson, A.J.; Royall, R.Q.; DeRouchey, J.M.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; Hastad, C.W.; Woodworth, J.C. Effects of timing and size of meals prior to farrowing on sow and litter performance. Transl. Anim. Sci. 2020, 4, 724–736. [Google Scholar] [CrossRef]
- Nam, N.H.; Sukon, P. Associated Factors for Farrowing Duration in Sows with Natural Parturition in Intensive Conditions. World 2020, 10, 320–324. [Google Scholar]
- Nielsen, S.E.; Feyera, T.; Skovmose, S.J.; Krogh, U.; Eskildsen, M.; Theil, P.K. Intravenous infusion of glucose improved farrowing performance of hyperprolific crossbred sows. J. Anim. Sci. 2021, 99, skab061. [Google Scholar] [CrossRef]
- Thongkhuy, S.; Chuaychu, S.H.B.; Burarnrak, P.; Ruangjoy, P.; Juthamanee, P.; Nuntapaitoon, M.; Tummaruk, P. Effect of backfat thickness during late gestation on farrowing duration, piglet birth weight, colostrum yield, milk yield and reproductive performance of sows. Livest. Sci. 2020, 234, 103983. [Google Scholar] [CrossRef]
- Dantzer, V.; Leiser, R. Initial vascularisation in the pig placenta: I. Demonstration of nonglandular areas by histology and corrosion casts. Anat. Rec. 1994, 238, 177–190. [Google Scholar] [CrossRef]
- Biensen, N.J.; Wilson, M.E.; Ford, S.P. The impact of either a Meishan or Yorkshire uterus on Meishan or Yorkshire fetal and placental development to days 70, 90, and 110 of gestation. J. Anim. Sci. 1998, 76, 2169–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.E.; Biensen, N.J.; Youngs, C.R.; Ford, S.P. Development of Meishan and Yorkshire littermate conceptuses in either a Meishan or Yorkshire uterine environment to day 90 of gestation and to term. Biol. Reprod. 1998, 58, 905–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, L.P.; Ford, S.P.; Ferrell, C.L. Blood Flow and Steroid and Nutrient Uptake of the Gravid Uterus and Fetus of Sows1. J. Anim. Sci. 1985, 61, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, A.D.; Aalhus, J.L.; Williams, N.H.; Patience, J.F. Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork. J. Anim. Sci. 2010, 88, 2767–2778. [Google Scholar] [CrossRef]
- Baxter, E.M.; Jarvis, S.; D’Eath, R.B.; Ross, D.W.; Robson, S.K.; Farish, M.; Nevison, I.M.; Lawrence, A.B.; Edwards, S.A. Investigating the behavioural and physiological indicators of neonatal survival in pigs. Theriogenology 2008, 69, 773–783. [Google Scholar] [CrossRef]
- Smith, D.G.; Schenk, M.P. A Dissection Guide & Atlas to the Fetal Pig; Morton Publishing Company: Englewood, Colorado, 2011; p. 72. [Google Scholar]
- Tantius, B.; Rothschild, M.A.; Valter, M.; Michael, J.; Banaschak, S. Experimental studies on the tensile properties of human umbilical cords. Forensic Sci. Int. 2014, 236, 16–21. [Google Scholar] [CrossRef]
- Leenhouwers, J.I.; Knol, E.F.; De Groot, P.N.; Vos, H.; Van Der Lende, T. Fetal development in the pig in relation to genetic merit for piglet survival. J. Anim. Sci. 2002, 80, 1759–1770. [Google Scholar] [CrossRef]
- Randall, G.C. Form and development of the umbilical cord in pigs and their association with delivery of viable pigs. Am. J. Vet. Res. 1989, 50, 1512–1515. [Google Scholar]
- Curtis, S.E. Responses of the piglet to perinatal stressors. J. Anim. Sci. 1974, 38, 1031–1036. [Google Scholar] [CrossRef]
- Tabeling, R.; Schwier, S.; Kamphues, J. Effects of different feeding and housing conditions on dry matter content and consistency of faeces in sows. J. Anim. Physiol. Anim. Nutr. 2003, 87, 116–121. [Google Scholar] [CrossRef]
- Cerisuelo, A.; Sala, R.; Gasa, J.; Carrion, D.; Coma, J.; Chapinal, N.; Baucells, M.D. Effects of extra feeding in mid-pregnancy for three successive parities on lean sows’ productive performance and longevity. Can. J. Anim. Sci. 2010, 90, 521–528. [Google Scholar] [CrossRef]
- Peltoniemi, O.; Oliviero, C.; Yun, J.; Grahofer, A.; Björkman, S. Management practices to optimize the parturition process in the hyperprolific sow. J. Anim. Sci. 2020, 98, S96–S106. [Google Scholar] [CrossRef] [PubMed]
- Feyera, T.; Skovmose, S.J.W.; Nielsen, S.E.; Vodolazska, D.; Bruun, T.S.; Theil, P.K. Optimal feed level during the transition period to achieve faster farrowing and high colostrum yield in sows. J. Anim. Sci. 2021, 99. [Google Scholar] [CrossRef] [PubMed]
- Van Kempen, T. Supplements to Facilitate Parturition and Reduce Perinatal Mortality in Pigs; Nottingham University Press: Nottinghan, UK, 2007. [Google Scholar]
- Gourdine, J.-L.; Bidanel, J.P.; Noblet, J.; Renaudeau, D. Rectal temperature of lactating sows in a tropical humid climate according to breed, parity and season. Asian-Australas. J. Anim. Sci. 2007, 20, 832–841. [Google Scholar] [CrossRef]
- van der Peet-Schwering, C.; Bikker, P. Energy and Amino Acid Requirements of Gestating and Lactating Sows; Wageningen Livestock Research: Wageningen, The Netherlands, 2019. [Google Scholar]
- Noblet, J.; Close, W.; Heavens, R.; Brown, D. Studies on the energy metabolism of the pregnant sow: 1. Uterus and mammary tissue development. Br. J. Nutr. 1985, 53, 251–265. [Google Scholar] [CrossRef] [Green Version]
- Van Milgen, J.; Noblet, J.; Dubois, S.; Bernier, J.-F. Dynamic aspects of oxygen consumption and carbon dioxide production in swine. Br. J. Nutr. 1997, 78, 397–410. [Google Scholar] [CrossRef] [Green Version]
- Che, L.; Hu, L.; Wu, C.; Xu, Q.; Zhou, Q.; Peng, X.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B. Effects of increased energy and amino acid intake in late gestation on reproductive performance, milk composition, metabolic, and redox status of sows. J. Anim. Sci. 2019, 97, 2914–2926. [Google Scholar] [CrossRef]
- Challis, J.R.; Matthews, S.G.; Gibb, W.; Lye, S.J. Endocrine and paracrine regulation of birth at term and preterm. Endocr. Rev. 2000, 21, 514–550. [Google Scholar]
- Rizzo, A.; Angioni, S.; Spedicato, M.; Minoia, G.; Mutinati, M.; Trisolini, C.; Sciorsci, R.L. Uterine contractility is strongly influenced by steroids and glucose metabolism: An in vitro study on bovine myometrium. Gynecol. Endocrinol. 2011, 27, 636–640. [Google Scholar] [CrossRef]
- Fereya, T. Nutrition of Transition Sows: Impacts on Farrowing Duration, Stillbirth Rate and Colostrogenesis. Ph.D. Thesis, Science and Technology Aarhus University, Aarhus, Denmark, 2018; pp. 15–16. [Google Scholar]
- Serena, A.; Hedemann, M.S.; Bach Knudsen, K.E. Feeding high fibre diets changes luminal environment and morphology in the intestine of sows. Livest. Sci. 2007, 109, 115–117. [Google Scholar] [CrossRef]
- O’Reilly, J.; Wong, S.H.; Chen, Y. Glycaemic index, glycaemic load and exercise performance. Sport. Med. 2010, 40, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Jamurtas, A.Z.; Deli, C.K.; Georgakouli, K.; Fatouros, I.G. Chapter 2—Glycemic Index, Food Exchange Values, and Exercise Performance. In Nutrition and Enhanced Sports Performance, 2nd ed.; Bagchi, D., Nair, S., Sen, C.K., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 11–29. [Google Scholar]
- Carsten, M.E.; Miller, J.D. A new look at uterine muscle contraction. Am. J. Obstet. Gynecol. 1987, 157, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Forman, A.; Andersson, K.; Ulmsten, U. Inhibition of myometrial activity by calcium antagonists. Semin. Perinatol. 1981, 5, 288–294. [Google Scholar] [PubMed]
- Geishauser, T.; Wegner, K.; Keller, A. Studies on the effect of a single dose of supplemental oral calcium prepartum on the incidence of dystocia in sows. Züchtungskunde 2012, 84, 330–339. [Google Scholar]
- Le Cozler, Y.; Guyomarc’h, C.; Pichodo, X.; Quinio, P.Y.; Pellois, H. Factors associated with stillborn and mummified piglets in high-prolific sows. Anim. Res. 2002, 51, 261–268. [Google Scholar] [CrossRef]
- Mercer, B.M.; Merlino, A.A.; Medicine, S.f.M.-F. Magnesium sulfate for preterm labor and preterm birth. Obstet. Gynecol. 2009, 114, 650–668. [Google Scholar] [CrossRef] [Green Version]
- Plush, K.J.; Lines, D.S.; Huser, J.S.; Hallett, S.H.; van Wettere, W.H.E.J. Farrowing performance of sows with increased magnesium concentrations in a transition diet. Anim. Prod. Sci. 2017, 57, 2446. [Google Scholar] [CrossRef]
- Crenshaw, T.D. Calcium, phosphorus, vitamin D, and vitamin K in swine nutrition. In Swine Nutrition; CRC Press: Boca Raton, FL, USA, 2000; pp. 207–232. [Google Scholar]
- Hamilton, B. Vitamin D and human skeletal muscle. Scand. J. Med. Sci. Sport. 2010, 20, 182–190. [Google Scholar] [CrossRef] [Green Version]
- Lauridsen, C.; Halekoh, U.; Larsen, T.; Jensen, S.K. Reproductive performance and bone status markers of gilts and lactating sows supplemented with two different forms of vitamin D1. J. Anim. Sci. 2010, 88, 202–213. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Nutrient Requirements of Swine; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Flohr, J.R.; Woodworth, J.C.; Bergstrom, J.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; DeRouchey, J.M. Evaluating the impact of maternal vitamin D supplementation: I. Sow performance, serum vitamin metabolites, and neonatal muscle characteristics. J. Anim. Sci. 2016, 94, 4629–4642. [Google Scholar] [CrossRef] [Green Version]
- Flohr, J.R.; Woodworth, J.C.; Bergstrom, J.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; DeRouchey, J.M. Evaluating the impact of maternal vitamin D supplementation on sow performance: II. Subsequent growth performance and carcass characteristics of growing pigs. J. Anim. Sci. 2016, 94, 4643–4653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thayer, M.T.; Nelssen, J.L.; Langemeier, A.J.; Morton, J.M.; Gonzalez, J.M.; Kruger, S.R.; Ou, Z.; Makowski, A.J.; Bergstrom, J.R. The effects of maternal dietary supplementation of cholecalciferol (vitamin D3) and 25 (OH) D3 on sow and progeny performance. Transl. Anim. Sci. 2019, 3, 692–708. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, Y.; Lv, G.; Zhuo, Y.; Lin, Y.; Feng, B.; Fang, Z.; Che, L.; Li, J.; Xu, S. Improving maternal vitamin D status promotes prenatal and postnatal skeletal muscle development of pig offspring. Nutrition 2016, 32, 1144–1152. [Google Scholar] [CrossRef] [PubMed]
- Palencia, J.; Lemes, M.; Garbossa, C.; Abreu, M.; Pereira, L.; Zangeronimo, M. Arginine for gestating sows and foetal development: A systematic review. J. Anim. Physiol. Anim. Nutr. 2018, 102, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Johnson, G.A.; Hou, Y. BOARD-INVITED REVIEW: Arginine nutrition and metabolism in growing, gestating, and lactating swine1,2. J. Anim. Sci. 2018, 96, 5035–5051. [Google Scholar] [CrossRef]
- van den Bosch, M.; Wijnen, J.; van de Linde, I.B.; van Wesel, A.A.M.; Melchior, D.; Kemp, B.; Clouard, C.; van den Brand, H. Effects of maternal dietary nitrate supplementation during the perinatal period on piglet survival, body weight, and litter uniformity. Transl. Anim. Sci. 2019, 3, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Moncada, S.; Higgs, A. The L-arginine-nitric oxide pathway. N. Engl. J. Med. 1993, 329, 2002–2012. [Google Scholar]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Govoni, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic. Biol. Med. 2004, 37, 395–400. [Google Scholar] [CrossRef]
- Webb, A.J.; Patel, N.; Loukogeorgakis, S.; Okorie, M.; Aboud, Z.; Misra, S.; Rashid, R.; Miall, P.; Deanfield, J.; Benjamin, N.; et al. Acute blood pressure lowering, vasoprotective, and antiplatelet properties of dietary nitrate via bioconversion to nitrite. Hypertension 2008, 51, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Bird, I.M.; Zhang, L.; Magness, R.R. Possible mechanisms underlying pregnancy-induced changes in uterine artery endothelial function. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2003, 284, R245–R258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [PubMed] [Green Version]
- Langendijk, P. Latest Advances in Sow Nutrition during Early Gestation. Animals 2021, 11, 1720. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Jiang, Z.; Lin, Y.; Zheng, C.; Zhou, G.; Chen, F.; Yang, L.; Wu, G. Dietary l-arginine supplementation enhances placental growth and reproductive performance in sows. Amino Acids 2012, 42, 2207–2214. [Google Scholar] [CrossRef]
- Quesnel, H.; Quiniou, N.; Roy, H.; Lottin, A.; Boulot, S.; Gondret, F. Supplying dextrose before insemination and L-arginine during the last third of pregnancy in sow diets: Effects on within-litter variation of piglet birth weight. J. Anim. Sci. 2014, 92, 1445–1450. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Che et al. [87] | Feyera et al. [81] | Average |
|---|---|---|---|
| Average litter size | 14.8 | 20.4 | |
| Average farrowing duration (min) | 238.4 | 359.8 | |
| Optimal daily energy intake determined based on the shortest duration of farrowing (MJ ME/sow/day) | 33.8 | 53.0 | |
| Energy requirement on the day of farrowing per piglet (MJ ME) | 2.28 | 2.60 | 2.44 |
| Energy requirement on the day of farrowing per 60 min of farrowing (MJ ME) | 8.49 | 8.83 | 8.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van den Bosch, M.; Soede, N.; Kemp, B.; van den Brand, H. Sow Nutrition, Uterine Contractions, and Placental Blood Flow during the Peri-Partum Period and Short-Term Effects on Offspring: A Review. Animals 2023, 13, 910. https://doi.org/10.3390/ani13050910
van den Bosch M, Soede N, Kemp B, van den Brand H. Sow Nutrition, Uterine Contractions, and Placental Blood Flow during the Peri-Partum Period and Short-Term Effects on Offspring: A Review. Animals. 2023; 13(5):910. https://doi.org/10.3390/ani13050910
Chicago/Turabian Stylevan den Bosch, Moniek, Nicoline Soede, Bas Kemp, and Henry van den Brand. 2023. "Sow Nutrition, Uterine Contractions, and Placental Blood Flow during the Peri-Partum Period and Short-Term Effects on Offspring: A Review" Animals 13, no. 5: 910. https://doi.org/10.3390/ani13050910
APA Stylevan den Bosch, M., Soede, N., Kemp, B., & van den Brand, H. (2023). Sow Nutrition, Uterine Contractions, and Placental Blood Flow during the Peri-Partum Period and Short-Term Effects on Offspring: A Review. Animals, 13(5), 910. https://doi.org/10.3390/ani13050910