
Black Bear Behavior and Movements Are Not Definitive Measures of Anthropogenic Food Use

 , and
, and 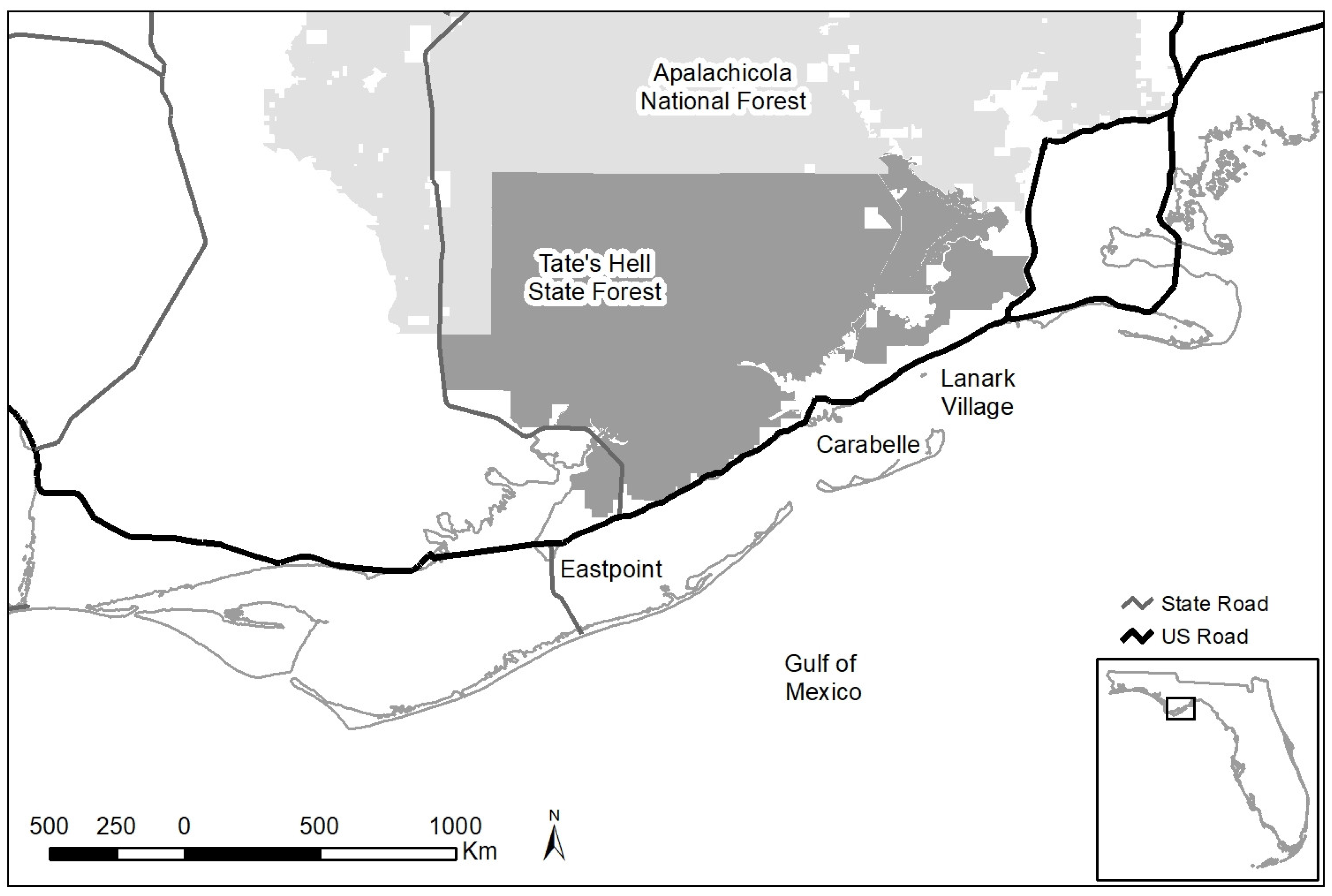{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Overview
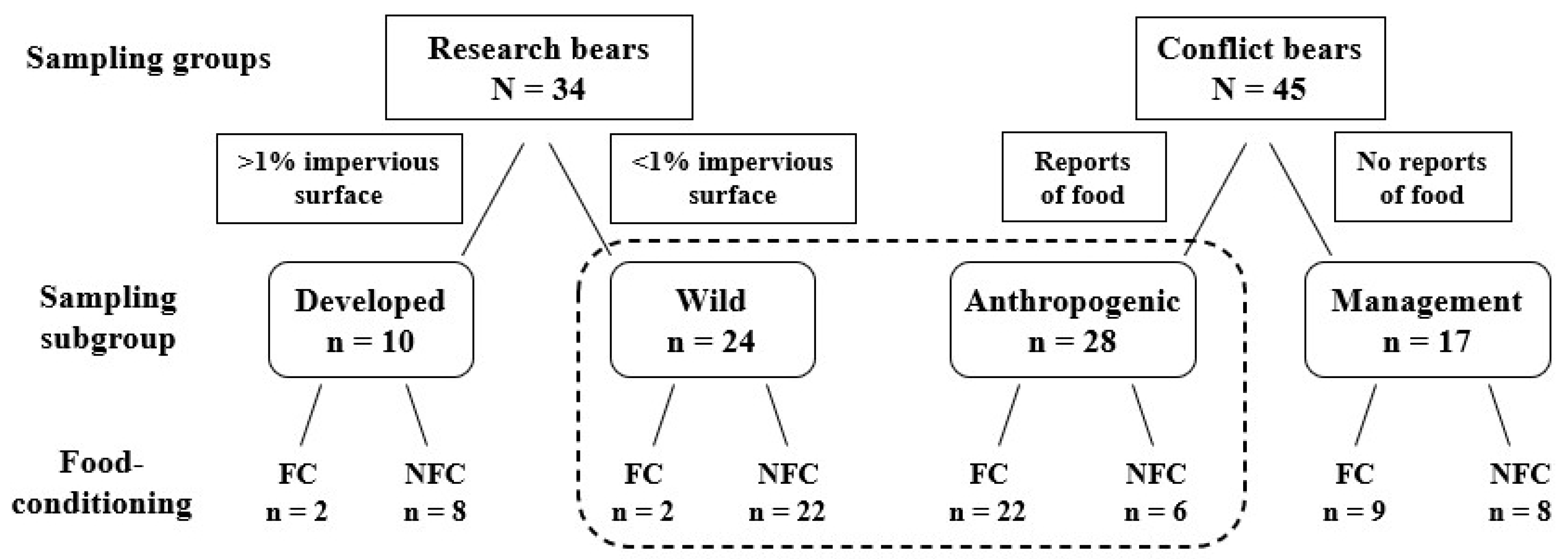
2.3. Classification of Bears
2.3.1. Capture and Monitoring of Research Bears
2.3.2. Research Bears
2.3.3. Conflict Bears
2.4. Stable Isotope Analysis
2.5. Statistical Analyses
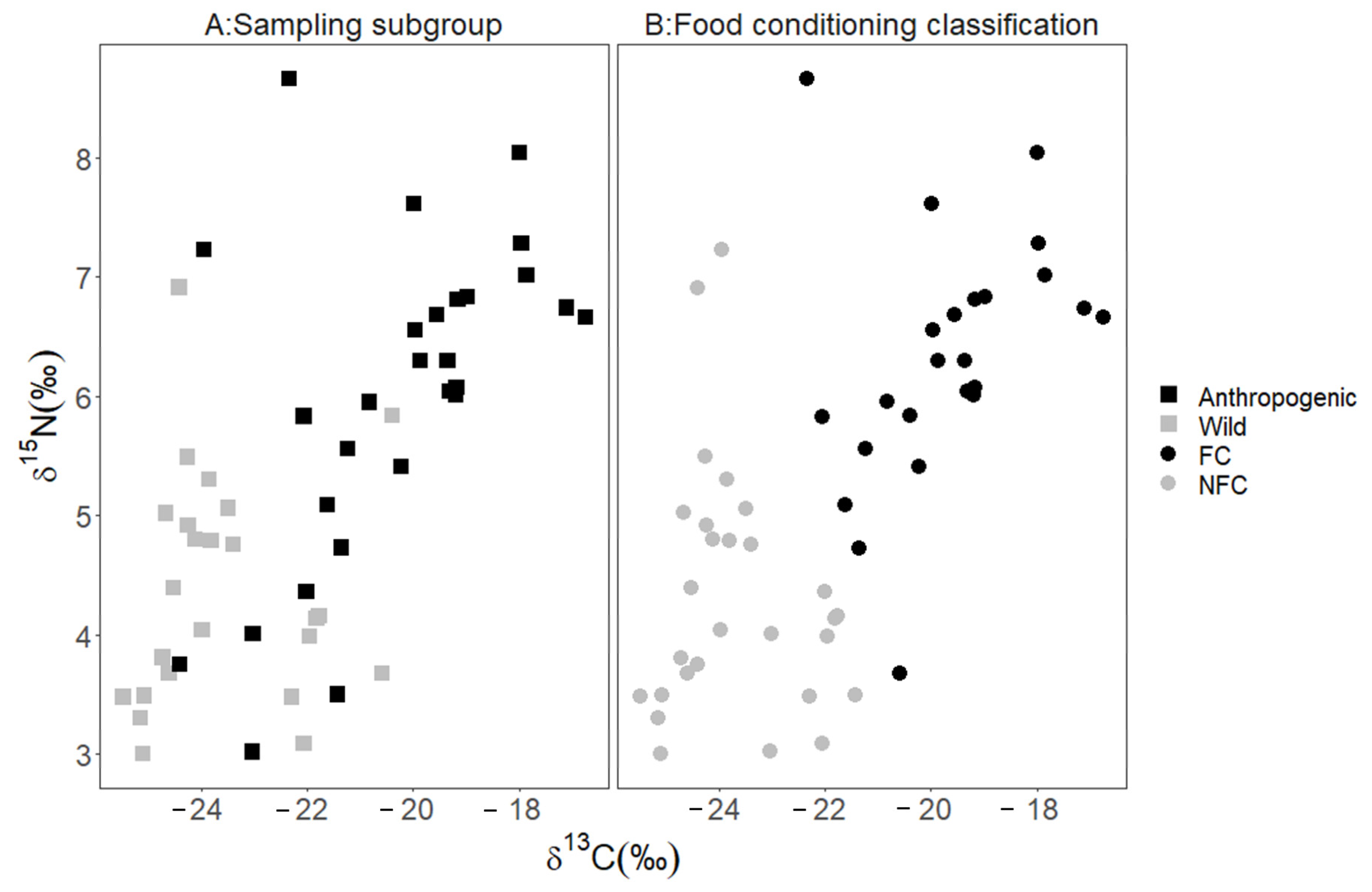
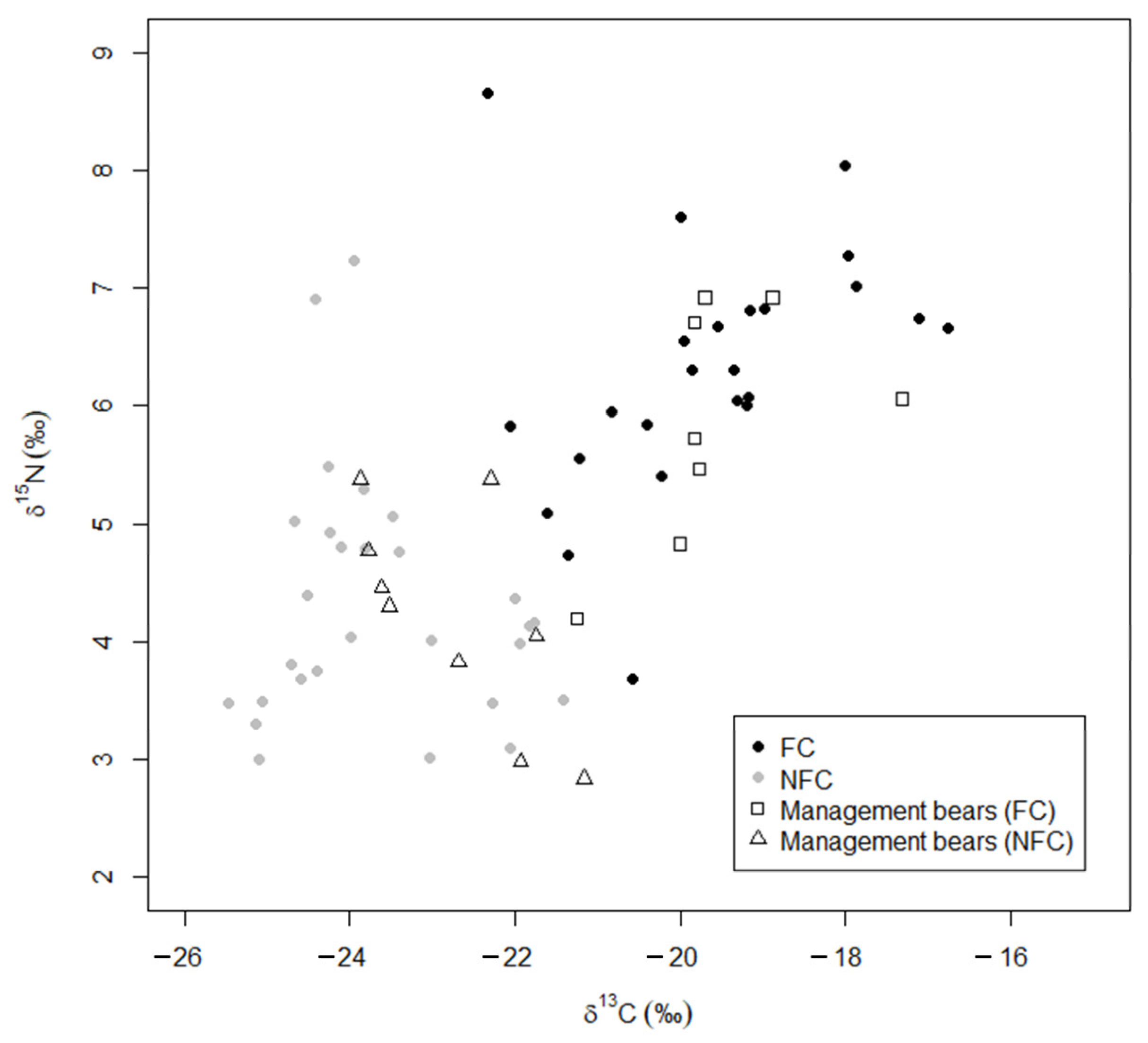
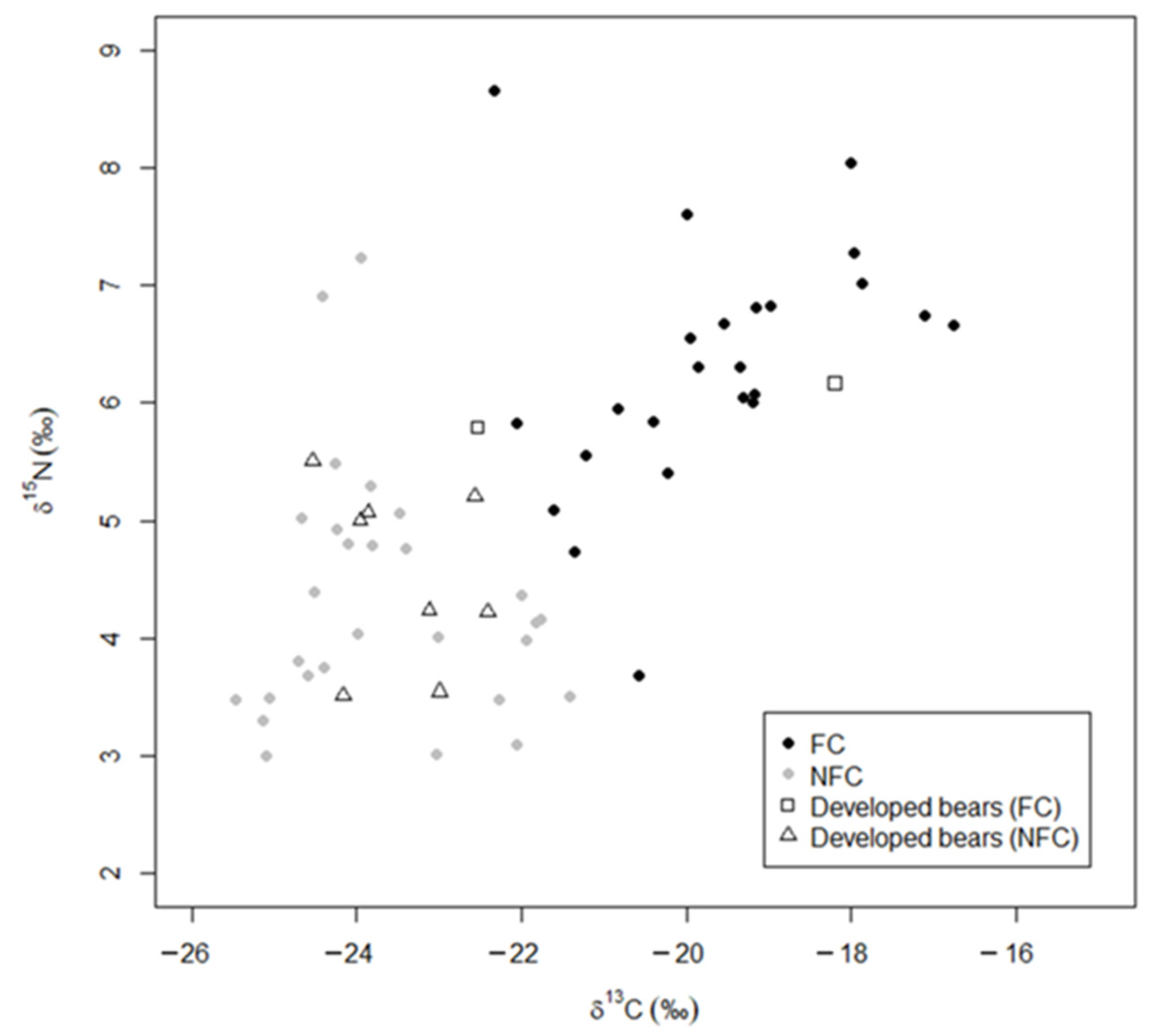
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linnell, J.D.C.; Swenson, J.E.; Andersen, R. Predators and people: Conservation of large carnivores is possible at high human densities if management policy is favourable. Anim. Conserv. 2001, 4, 345–349. [Google Scholar] [CrossRef]
- DeFries, R.S.; Foley, J.A.; Asner, G.P. Land-use choices: Balancing human needs and ecosystem function. Front. Ecol. Environ. 2004, 2, 249–257. [Google Scholar] [CrossRef]
- Marucco, F.; McIntire, E.J.B. Predicting spatio-temporal recolonization of large carnivore populations and livestock depredation risk: Wolves in the Italian Alps. J. Appl. Ecol. 2010, 47, 789–798. [Google Scholar] [CrossRef]
- Chapron, G.; Lopez-Bao, J.V. Conserving Carnivores: Politics in Play. Science 2014, 343, 1199–1200. [Google Scholar] [CrossRef]
- Hopkins, J.B.; Herrero, S.; Shideler, R.T.; Gunther, K.A.; Schwartz, C.C.; Kalinowski, S.T. A proposed lexicon of terms and concepts for human-bear management in North America. Ursus 2010, 21, 154–168. [Google Scholar] [CrossRef]
- Treves, A.; Jurewicz, R.R.; Naughton-Treves, L. Wolf depredation on domestic animals in Wisconsin, 1976–2000. Wildl. Soc. Bull. 2002, 30, 231–241. [Google Scholar]
- Treves, A.; Karanth, K.U. Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv. Biol. 2003, 17, 1491–1499. [Google Scholar] [CrossRef]
- Peterson, M.N.; Birckhead, J.L.; Leong, K.; Peterson, M.J.; Peterson, T.R. Rearticulating the myth of human-wildlife conflict. Conserv. Lett. 2010, 3, 74–82. [Google Scholar] [CrossRef]
- Oakleaf, J.K.; Mack, C.; Murray, D.L. Effects of wolves on livestock calf survival and movements in central Idaho. J. Wildl. Manag. 2003, 67, 299–306. [Google Scholar] [CrossRef]
- Treves, A.; Naughton-Treves, L.; Harper, E.K.; Mladenoff, D.J.; Rose, R.A.; Sickley, T.A.; Wydeven, A.P. Predicting human-carnivore conflict: A spatial model derived from 25 years of data on wolf predation on livestock. Conserv. Biol. 2004, 18, 114–125. [Google Scholar] [CrossRef]
- Alldredge, M.W.; Walsh, D.P.; Sweanor, L.L.; Davies, R.B.; Trujillo, A. Evaluation of Translocation of Black Bears Involved in Human-Bear Conflicts in South-Central Colorado. Wildl. Soc. Bull. 2015, 39, 334–340. [Google Scholar] [CrossRef]
- Baruch-Mordo, S.; Wilson, K.R.; Lewis, D.L.; Broderick, J.; Mao, J.S.; Breck, S.W. Stochasticity in Natural Forage Production Affects Use of Urban Areas by Black Bears: Implications to Management of Human-Bear Conflicts. PLoS ONE 2014, 9, e85122. [Google Scholar] [CrossRef]
- Nellemann, C.; Stoen, O.G.; Kindberg, J.; Swenson, J.E.; Vistnes, I.; Ericsson, G.; Katajisto, J.; Kaltenborn, B.P.; Martin, J.; Ordiz, A. Terrain use by an expanding brown bear population in relation to age, recreational resorts and human settlements. Biol. Conserv. 2007, 138, 157–165. [Google Scholar] [CrossRef]
- Ordiz, A.; Støen, O.-G.; Delibes, M.; Swenson, J.E. Predators or prey? Spatio-temporal discrimination of human-derived risk by brown bears. Oecologia 2011, 166, 59–67. [Google Scholar] [CrossRef]
- Nelson, R.A.; Folk Jr, G.E.; Pfeiffer, E.W.; Craighead, J.J.; Jonkel, C.J.; Steiger, D.L. Behavior, biochemistry, and hibernation in black, grizzly, and polar bears. Bears Biol. Manag. 1983, 5, 284–290. [Google Scholar] [CrossRef]
- Peine, J.D. Nuisance bears in communities: Strategies to reduce conflict. Hum. Dimens. Wildl. 2001, 6, 223–237. [Google Scholar] [CrossRef]
- Johnson, H.E.; Breck, S.W.; Baruch-Mordo, S.; Lewis, D.L.; Lackey, C.W.; Wilson, K.R.; Broderick, J.; Mao, J.S.; Beckmann, J.P. Shifting perceptions of risk and reward: Dynamic selection for human development by black bears in the western United States. Biol. Conserv. 2015, 187, 164–172. [Google Scholar] [CrossRef]
- Hristienko, H.; McDonald, J.E. Going into the 21(st) century: A perspective on trends and controversies in the management of the American black bear. Ursus 2007, 18, 72–88. [Google Scholar] [CrossRef]
- Spencer, R.D.; Beausoleil, R.A.; Martorello, D.A. How agencies respond to human-black bear conflicts: A survey of wildlife agencies in North America. Ursus 2007, 18, 217–229. [Google Scholar] [CrossRef]
- Merkle, J.A.; Robinson, H.S.; Krausman, P.R.; Alaback, P. Food availability and foraging near human developments by black bears. J. Mammal. 2013, 94, 378–385. [Google Scholar] [CrossRef]
- Lackey, C.W.; Breck, S.W.; Wakeling, B.F.; White, H.B. Human–Black Bear Conflicts: A Review of Common Management Practices. Hum. Widl. Interact. Monogr. 2018, 2, 1–68. [Google Scholar]
- Siemer, W.F.; Hart, P.S.; Decker, D.J.; Shanahan, J.E. Factors that influence concern about human–black bear interactions in residential settings. Hum. Dimens. Wildl. 2009, 14, 185–197. [Google Scholar] [CrossRef]
- Mazur, R.; Seher, V. Socially learned foraging behaviour in wild black bears, Ursus americanus. Anim. Behav. 2008, 75, 1503–1508. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Matthews, S.M.; Wright, R.G.; Beecham, J.J.; Leithead, H.M. Food habits of American black bears as a metric for direct management of human-bear conflict in Yosemite Valley, Yosemite National Park, California. Ursus 2009, 20, 94–101. [Google Scholar] [CrossRef]
- Newsome, T.M.; Dellinger, J.A.; Pavey, C.R.; Ripple, W.J.; Shores, C.R.; Wirsing, A.J.; Dickman, C.R. The ecological effects of providing resource subsidies to predators. Glob. Ecol. Biogeogr. 2015, 24, 1–11. [Google Scholar] [CrossRef]
- Veitch, A.; Harrington, F. Brown bears, black bears, and humans in northern Labrador: An historical perspective and outlook to the future. J. Wildl. Res. 1996, 1, 245–250. [Google Scholar]
- Stirling, I.; Derocher, A.E. Factors affecting the evolution and behavioral ecology of the modern bears. Bears Biol. Manag. 1990, 8, 189–204. [Google Scholar] [CrossRef]
- Matthews, S.M.; Beecham, J.J.; Quigley, H.; Greenleaf, S.S.; Leithead, H.M. Activity patterns of American black bears in Yosemite National Park. Ursus 2006, 17, 30–40. [Google Scholar] [CrossRef]
- Herrero, S.; Smith, T.; De Bruyn, T.D.; Gunther, K.; Matt, C.A. From the field: Brown bear habituation to people—Safety, risks, and benefits. Wildl. Soc. Bull. 2005, 33, 362–373. [Google Scholar] [CrossRef]
- Elfstrom, M.; Davey, M.L.; Zedrosser, A.; Muller, M.; De Barba, M.; Stoen, O.G.; Miquel, C.; Taberlet, P.; Hacklander, K.; Swenson, J.E. Do Scandinavian brown bears approach settlements to obtain high-quality food? Biol. Conserv. 2014, 178, 128–135. [Google Scholar] [CrossRef]
- Beckmann, J.P.; Berger, J. Rapid ecological and behavioural changes in carnivores: The responses of black bears (Ursus americanus) to altered food. J. Zool. 2003, 261, 207–212. [Google Scholar] [CrossRef]
- Gunther, K.A.; Haroldson, M.A.; Frey, K.; Cain, S.L.; Copeland, J.; Schwartz, C.C. Grizzly bear-human conflicts in the Greater Yellowstone ecosystem, 1992–2000. Ursus 2004, 15, 10–22. [Google Scholar] [CrossRef]
- Clark, J.E.; van Manen, F.T.; Pelton, M.R. Correlates of success for on-site releases of nuisance black bears in Great Smoky Mountains National Park. Wildl. Soc. Bull. 2002, 30, 104–111. [Google Scholar]
- Gore, M.L.; Siemer, W.F.; Shanahan, J.E.; Scheufele, D.; Decker, D.J. Effects on risk perception of media coverage of a black bear-related human fatality. Wildl. Soc. Bull. 2005, 33, 507–516. [Google Scholar] [CrossRef]
- Elfstrom, M.; Zedrosser, A.; Stoen, O.G.; Swenson, J.E. Ultimate and proximate mechanisms underlying the occurrence of bears close to human settlements: Review and management implications. Mamm. Rev. 2014, 44, 5–18. [Google Scholar] [CrossRef]
- Don Carlos, A.W.; Bright, A.D.; Teel, T.L.; Vaske, J.J. Human–black bear conflict in urban areas: An integrated approach to management response. Hum. Dimens. Wildl. 2009, 14, 174–184. [Google Scholar] [CrossRef]
- Braunstein, J.L.; Clark, J.D.; Williamson, R.H.; Stiver, W.H. Black Bear Movement and Food Conditioning in an Exurban Landscape. J. Wildl. Manag. 2020, 84, 1038–1050. [Google Scholar] [CrossRef]
- Whittaker, D.; Vaske, J.J.; Manfredo, M.J. Specificity and the cognitive hierarchy: Value orientations and the acceptability of urban wildlife management actions. Soc. Nat. Resour. 2006, 19, 515–530. [Google Scholar] [CrossRef]
- Vulla, E.; Hobson, K.A.; Korsten, M.; Leht, M.; Martin, A.J.; Lind, A.; Mannil, P.; Valdmann, H.; Saarma, U. Carnivory is positively correlated with latitude among omnivorous mammals: Evidence from brown bears, badgers and pine martens. Ann. Zool. Fenn. 2009, 46, 395–415. [Google Scholar] [CrossRef]
- Newsome, S.D.; Ralls, K.; Job, C.V.; Fogel, M.L.; Cypher, B.L. Stable isotopes evaluate exploitation of anthropogenic foods by the endangered San Joaquin kit fox (Vulpes macrotis mutica). J. Mammal. 2010, 91, 1313–1321. [Google Scholar] [CrossRef]
- Kelly, J.F. Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology. Can. J. Zool. 2000, 78, 1–27. [Google Scholar] [CrossRef]
- Hobson, K.A.; McLellan, B.N.; Woods, J.G. Using stable carbon (delta C-13) and nitrogen (delta N-15) isotopes to infer trophic relationships among black and grizzly bears in the upper Columbia River basin, British Columbia. Can. J. Zool. 2000, 78, 1332–1339. [Google Scholar] [CrossRef]
- Ben-David, M.; Flaherty, E.A. Stable isotopes in mammalian research: A beginner’s guide. J. Mammal. 2012, 93, 312–328. [Google Scholar] [CrossRef]
- Schwertl, M.; Auerswald, K.; Schnyder, H. Reconstruction of the isotopic history of animal diets by hair segmental analysis. Rapid Commun. Mass Spectrom. 2003, 17, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Mowat, G.; Heard, D.C. Major components of grizzly bear diet across North America. Can. J Zool. 2006, 84, 473–489. [Google Scholar] [CrossRef]
- Chesson, L.A.; Podlesak, D.W.; Thompson, A.H.; Cerling, T.E.; Ehleringer, J.R. Variation of hydrogen, carbon, nitrogen, and oxygen stable isotope ratios in an American diet: Fast food meals. J. Agric. Food Chem. 2008, 56, 4084–4091. [Google Scholar] [CrossRef]
- Hopkins, J.B.; Koch, P.L.; Schwartz, C.C.; Ferguson, J.M.; Greenleaf, S.S.; Kalinowski, S.T. Stable isotopes to detect food-conditioned bears and to evaluate human-bear management. J. Wildl. Manag. 2012, 76, 703–713. [Google Scholar] [CrossRef]
- Maehr, D.S.; Brady, J.R. Food-habits of Florida black bears. J. Wildl. Manag. 1984, 48, 230–235. [Google Scholar] [CrossRef]
- Humm, J.M.; McCown, J.W.; Scheick, B.K.; Clark, J.D. Spatially explicit population estimates for black bears based on cluster sampling. J. Wildl. Manag. 2017, 81, 1187–1201. [Google Scholar] [CrossRef]
- Johnson, K.G.; Pelton, M.R. Prebaiting and snaring techniques for black bears. Wildl. Soc. Bull. 1980, 8, 46–54. [Google Scholar]
- Scheick, B.K.; Cunningham, M.W.; McCown, J.W.; Orlando, M.A. Anchor modification for a foot-hold snare to capture American black bears. Ursus 2009, 20, 47–49. [Google Scholar] [CrossRef]
- Sutton, P.C.; Anderson, S.J.; Elvidge, C.D.; Tuttle, B.T.; Ghosh, T. Paving the planet: Impervious surface as proxy measure of the human ecological footprint. Prog. Phys. Geogr. 2009, 33, 510–527. [Google Scholar] [CrossRef]
- Schmidt, J.A.; McCleery, R.; Seavey, J.R.; Devitt, S.E.; Schmidt, P.M. Impacts of a half century of sea-level rise and development on an endangered mammal. Glob. Chang. Biol. 2012, 18, 3536–3542. [Google Scholar] [CrossRef]
- Calenge, C. The package “adehabitat” for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Modell. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Homer, C.H.; Fry, J.A.; Barnes, C.A. The national land cover database. US Geol. Surv. Fact Sheet 2012, 3020, 1–4. [Google Scholar]
- Hijmans, R.J.; Van Etten, J.; Cheng, J.; Mattiuzzi, M.; Sumner, M.; Greenberg, J.A.; Lamigueiro, O.P.; Bevan, A.; Racine, E.B.; Shortridge, A. Package ‘raster’. R Package 2015, 734, 473. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; ISBN 3-900051-07-0; Available online: http://www.R-project.org/ (accessed on 20 February 2018)ISBN 3-900051-07-0.
- Jimbo, M.; Matsumoto, N.; Sakamoto, H.; Yanagawa, Y.; Torii, Y.; Yamanaka, M.; Ishinazaka, T.; Shirane, Y.; Sashika, M.; Tsubota, T.; et al. Hair growth in brown bears and its application to ecological studies on wild bears. Mammal Study 2020, 45, 337–345. [Google Scholar] [CrossRef]
- Careddu, G.; Ciucci, P.; Mondovi, S.; Calizza, E.; Rossi, L.; Costantini, M.L. Gaining insight into the assimilated diet of small bear populations by stable isotope analysis. Sci. Rep. 2021, 11, 14118. [Google Scholar] [CrossRef]
- Bearhop, S.; Adams, C.E.; Waldron, S.; Fuller, R.A.; Macleod, H. Determining trophic niche width: A novel approach using stable isotope analysis. J. Anim. Ecol. 2004, 73, 1007–1012. [Google Scholar] [CrossRef]
- Jacoby, M.E.; Hilderbrand, G.V.; Servheen, C.; Schwartz, C.C.; Arthur, S.M.; Hanley, T.A.; Robbins, C.T.; Michener, R. Trophic relations of brown and black bears in several western North American ecosystems. J. Wildl. Manag. 1999, 63, 921–929. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Kirby, R.; Alldredge, M.W.; Pauli, J.N. The diet of black bears tracks the human footprint across a rapidly developing landscape. Biol. Conserv. 2016, 200, 51–59. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.; Stafford, S. Multivariate Statistics for Wildlife and Ecology Research; Spring: New York, NY, USA, 2000; 283p. [Google Scholar]
- Jepsen, D.B.; Winemiller, K.O. Structure of tropical river food webs revealed by stable isotope ratios. Oikos 2002, 96, 46–55. [Google Scholar] [CrossRef]
- Rosenberger, A.E.; Dunham, J.B. Validation of abundance estimates from mark-recapture and removal techniques for rainbow trout captured by electrofishing in small streams (vol 25, pg 1395, 2006). N. Am. J. Fish. Manag. 2006, 26, 502. [Google Scholar] [CrossRef]
- Bentzen, T.W.; Shideler, R.T.; O’Hara, T.M. Use of stable isotope analysis to identify food-conditioned grizzly bears on Alaska’s North Slope. Ursus 2014, 25, 14–23. [Google Scholar] [CrossRef]
- Beckmann, J.P.; Berger, J. Using black bears to test ideal-free distribution models experimentally. J. Mammal. 2003, 84, 594–606. [Google Scholar] [CrossRef]
- Barrett, M.A.; Telesco, D.J.; Barrett, S.E.; Widness, K.M.; Leone, E.H. Testing Bear-Resistant Trash Cans in Residential Areas of Florida. Southeast. Nat. 2014, 13, 26–39. [Google Scholar] [CrossRef]
- Johnson, H.E.; Lewis, D.L.; Lischka, S.A.; Breck, S.W. Assessing ecological and social outcomes of a bear-proofing experiment. J. Wildl. Manag. 2018, 82, 1102–1114. [Google Scholar] [CrossRef]
- Lischka, S.A.; Teel, T.L.; Johnson, H.E.; Crooks, K.R. Understanding and managing human tolerance for a large carnivore in a residential system. Biol. Conserv. 2019, 238, 108189. [Google Scholar] [CrossRef]
- Sikes, R.S. Animal Care and Use Committee of the American Society of Mammalogists. Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardeman, D.W., Jr.; Vander Zanden, H.B.; McCown, J.W.; Scheick, B.K.; McCleery, R.A. Black Bear Behavior and Movements Are Not Definitive Measures of Anthropogenic Food Use. Animals 2023, 13, 950. https://doi.org/10.3390/ani13050950
Hardeman DW Jr., Vander Zanden HB, McCown JW, Scheick BK, McCleery RA. Black Bear Behavior and Movements Are Not Definitive Measures of Anthropogenic Food Use. Animals. 2023; 13(5):950. https://doi.org/10.3390/ani13050950
Chicago/Turabian StyleHardeman, Don W., Jr., Hannah B. Vander Zanden, J. Walter McCown, Brian K. Scheick, and Robert A. McCleery. 2023. "Black Bear Behavior and Movements Are Not Definitive Measures of Anthropogenic Food Use" Animals 13, no. 5: 950. https://doi.org/10.3390/ani13050950
APA StyleHardeman, D. W., Jr., Vander Zanden, H. B., McCown, J. W., Scheick, B. K., & McCleery, R. A. (2023). Black Bear Behavior and Movements Are Not Definitive Measures of Anthropogenic Food Use. Animals, 13(5), 950. https://doi.org/10.3390/ani13050950