
HV1 mtDNA Reveals the High Genetic Diversity and the Ancient Origin of Vietnamese Dogs
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
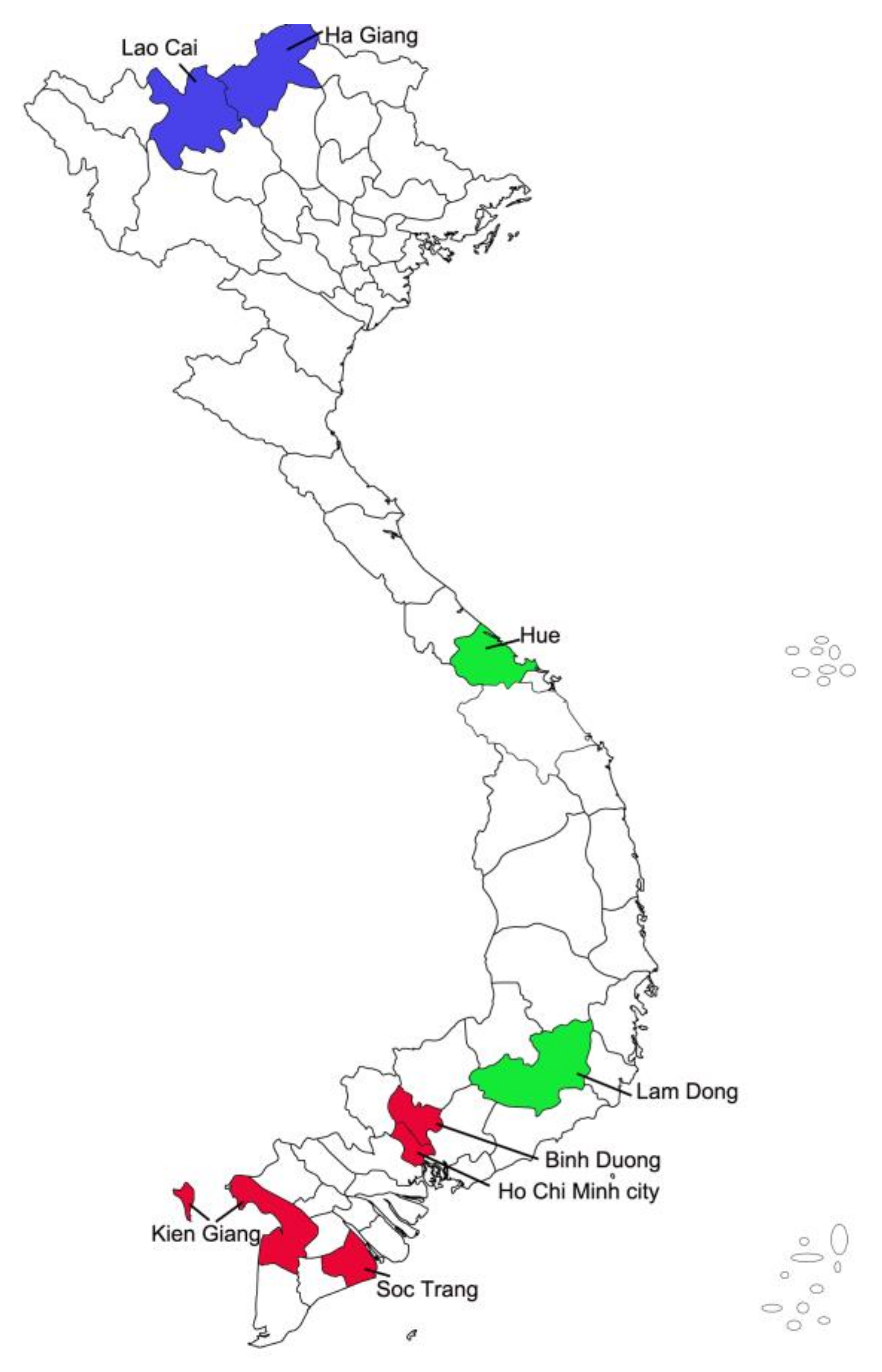
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. DNA Amplification
2.4. DNA Sequencing
2.5. Haplotyping DNA Sequencing
2.6. Data Analysis DNA Sequencing
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leonard, J.A.; Wayne, R.K.; Wheeler, J.; Valadez, R.; Guillen, S.; Vila, C. Ancient DNA evidence for Old World origin of New World dogs. Science 2002, 298, 1613–1616. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, P.; Zhang, Y.P.; Luo, J.; Lundeberg, J.; Leitner, T. Genetic evidence for an East Asian origin of domestic dogs. Science 2002, 298, 1610–1613. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V.J.; Sawyer, S.K.; Greenfield, D.L.; Germonpre, M.B.; Sablin, M.V.; Lopez-Giraldez, F.; Domingo-Roura, X.; et al. Complete mitochondrial genomes of ancient canids suggest a European origin of domestic dogs. Science 2013, 342, 871–874. [Google Scholar] [CrossRef]
- Frantz, L.A.; Mullin, V.E.; Pionnier-Capitan, M.; Lebrasseur, O.; Ollivier, M.; Perri, A.; Linderholm, A.; Mattiangeli, V.; Teasdale, M.D.; Dimopoulos, E.A.; et al. Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science 2016, 352, 1228–1231. [Google Scholar] [CrossRef] [PubMed]
- Ardalan, A.; Oskarsson, M.C.R.; Asch, B.V.; Rabakonandriania, E.; Savolainen, P. African origin for Madagascan dogs revealed by mtDNA analysis. R. Soc. Open Sci. 2015, 2, 140552. [Google Scholar] [CrossRef]
- Van Asch, B.; Zhang, A.-B.; Oskarsson, M.C.R.; Klütsch, C.F.C.; Amorim, A.; Savolainen, P. Pre-Columbian origins of Native American dog breeds, with only limited replacement by European dogs, confirmed by mtDNA analysis. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280, 20131142. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Z.; Li, Y.; Zhao, X.; Dong, L.; Pan, Z.; Sun, Y.; Li, N.; Xu, Y.; Xie, Z. Origin and phylogenetic analysis of Tibetan Mastiff based on the mitochondrial DNA sequence. J. Genet. Genom. 2008, 35, 335–340. [Google Scholar] [CrossRef]
- Bekaert, B.; Larmuseau, M.H.D.; Vanhove, M.P.M.; Opdekamp, A.; Decorte, R. Automated DNA extraction of single dog hairs without roots for mitochondrial DNA analysis. Forensic Sci. Int. Genet. 2012, 6, 277–281. [Google Scholar] [CrossRef]
- Kim, K.S.; Lee, S.E.; Jeong, H.W.; Ha, J.H. The complete nucleotide sequence of the domestic dog (Canis familiaris) mitochondrial genome. Mol. Phylogenetics Evol. 1998, 10, 210–220. [Google Scholar] [CrossRef]
- Pang, J.-F.; Kluetsch, C.; Zou, X.-J.; Zhang, A.-B.; Luo, L.-Y.; Angleby, H.; Ardalan, A.; Ekstrom, C.; Skollermo, A.; Lundeberg, J.; et al. mtDNA data indicate a single origin for dogs south of Yangtze River, less than 16,300 years ago, from numerous wolves. Mol. Biol. Evol. 2009, 26, 2849–2864. [Google Scholar] [CrossRef]
- Hillbertz, N.H.C.S. The origin of the ridge and associated anomalies in Rhodesian Ridgebacks; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2007. [Google Scholar]
- Quan, T.K.; Tu, N.V.; Trinh, T.N.; Hieu, H.V.; Dung, C.A.; Dung, T.H. Evaluation of genetic diversity of Phu Quoc ridgeback dogs based on mitochondrial DNA Hypervariable-1 region. Vietnam J. Biotechnol. 2016, 14, 245–253. [Google Scholar]
- Quan, T.K.; Tu, N.V.; Hieu, H.V.; Cong, N.T.; Dung, T.H. A simple protocol for DNA extraction from dog hairs. Vietnam J. Biol. 2016, 38, 124–132. [Google Scholar] [CrossRef]
- Gundry, R.L.; Allard, M.W.; Moretti, T.R.; Honeycutt, R.L.; Wilson, M.R.; Monson, K.L.; Foran, D.R. Mitochondrial DNA analysis of the domestic dog: Control region variation within and among breeds. J. Forensic Sci. 2007, 52, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Thai, Q.K.; Chung, D.A.; Tran, H.-D. Canis mtDNA HV1 database: A web-based tool for collecting and surveying Canis mtDNA HV1 haplotype in public database. BMC Genet. 2017, 18, 60. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Van Asch, B.; Pereira, L.; Pereira, F.; Santa-Rita, P.; Lima, M.; Amorim, A. MtDNA diversity among four Portuguese autochthonous dog breeds: A fine-scale characterisation. BMC Genet. 2005, 6, 37. [Google Scholar] [CrossRef]
- Koban, E.; Gökçek Saraç, Ç.; Açan, S.C.; Savolainen, P.; Togan, İ. Genetic relationship between Kangal, Akbash and other dog populations. Discret. Appl. Math. 2009, 157, 2335–2340. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.; Thai Ke, Q.; Ardalan, A.; Boonyaprakob, U.; Savolainen, P. Complete range of the universal mtDNA gene pool and high genetic diversity in the thai dog population. Genes 2020, 11, 253. [Google Scholar] [CrossRef]
- Ren, Z.; Chen, H.; Yang, X.; Zhang, C. Phylogenetic analysis of Tibetan mastiffs based on mitochondrial hypervariable region I. J. Genet. 2017, 96, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Takahasi, S.; Miyahara, K.; Ishikawa, H.; Ishiguro, N.; Suzuki, M. Lineage classification of canine inheritable disorders using mitochondrial DNA haplotypes. J. Vet. Med. Sci. 2002, 64, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Okumura, N.; Ishiguro, N.; Nakano, M.; Matsui, A.; Sahara, M. Intra- and interbreed genetic variations of mitochondrial DNA major non-coding regions in Japanese native dog breeds (Canis familiaris). Anim. Genet. 1996, 27, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Klutsch, C.F.; Seppala, E.H.; Fall, T.; Uhlen, M.; Hedhammar, A.; Lohi, H.; Savolainen, P. Regional occurrence, high frequency but low diversity of mitochondrial DNA haplogroup d1 suggests a recent dog-wolf hybridization in Scandinavia. Anim. Genet. 2011, 42, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Oskarsson, M.C.; Klutsch, C.F.; Boonyaprakob, U.; Wilton, A.; Tanabe, Y.; Savolainen, P. Mitochondrial DNA data indicate an introduction through Mainland Southeast Asia for Australian dingoes and Polynesian domestic dogs. Proc. R. Soc. B 2012, 279, 967–974. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y. High genetic diversity of Tibetan Mastiffs revealed by mtDNA sequences. Chin. Sci. Bull. 2012, 57, 1483–1487. [Google Scholar] [CrossRef]
- Imes, D.L.; Wictum, E.J.; Allard, M.W.; Sacks, B.N. Identification of single nucleotide polymorphisms within the mtDNA genome of the domestic dog to discriminate individuals with common HVI haplotypes. Forensic Sci. Int. Genet. 2012, 6, 630–639. [Google Scholar] [CrossRef]
- Wang, G.-D.; Zhai, W.; Yang, H.-C.; Wang, L.; Zhong, L.; Liu, Y.-H.; Fan, R.-X.; Yin, T.-T.; Zhu, C.-L.; Poyarkov, A.D.; et al. Out of southern East Asia: The natural history of domestic dogs across the world. Cell Res. 2016, 26, 21–33. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Regions | ABC (DEF) | nA (%) | nB (%) | nC (%) | nE (%) | nHT | nHTres44 | HTuq | Haplotype Diversity | Nucleotide Diversity | Mean of Pairwise Differences |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | 394 (35) | 232 (54.1) | 97 (22.6) | 65 (15.2) | 35 (8.2) | 56 | 18.96 (± 2.32) | 14 | 0.9210 ± 0.0065 | 0.014819 ± 0.007586 | 8.639566 ± 3.998569 |
| SVN | 284 (34) | 153 (48.1) | 75 (23.6) | 56 (17.6) | 34 (10.7) | 31 | 15.16 (± 1.28) | 3 | 0.8994 ± 0.0079 | 0.014596 ± 0.007486 | 8.509454 ± 3.945443 |
| Rach Gia | 78 (5) | 49 (59) | 15 (18.1) | 14 (16.9) | 5 (6) | 18 | 1 * | ||||
| Ha Tien | 15 (0) | 8 (53.3) | 5 (33.3) | 2 (13.3) | 0 (0) | 7 | 0 | ||||
| Phu Quoc | 118 (15) | 56 (42.1) | 39 (29.3) | 23 (17.3) | 15 (11.3) | 17 | 1 * | ||||
| Soc Trang | 21 (0) | 12 (57.1) | 4 (19) | 5 (23.8) | 0 (0) | 14 | 1 | ||||
| Binh Duong | 7 (0) | 6 (85.7) | 1 (14.3) | 0 (0) | 0 (0) | 4 | 0 | ||||
| Ho Chi minh city | 45 (14) | 22 (37.3) | 11 (18.6) | 12 (20.3) | 14 (23.7) | 17 | 1 | ||||
| CVN | 44 (0) | 23 (52.3) | 15 (34.1) | 6 (13.7) | 0 (0) | 23 | 23 | 5 | 0.9154 ± 0.0327 | 0.015095 ± 0.007880 | 8.800211 ± 4.137619 |
| Lam Dong | 26 (0) | 12 (46.2) | 9 (34.6) | 5 (19.2) | 0 (0) | 12 | 2 * | ||||
| Hoi An | 3 (0) | 2 (66.7) | 0 (0) | 1 (33.3) | 0 (0) | 12 | 0 | ||||
| Hue | 15 (0) | 9 (60) | 6 (40) | 0 (0) | 0 (0) | 3 | 3 | ||||
| NVN | 55 (0) | 50 (90.9) | 3 (5.5) | 2 (3.6) | 0 (0) | 26 | 22.4 (± 1.41) | 7 | 0.9219 ± 0.0242 | 0.009830 ± 0.005304 | 5.721212 ± 2.782501 |
| Sapa | 9 (0) | 9 (100) | 0 (0) | 0 (0) | 0 (0) | 7 | 0 | ||||
| Bac Ha | 15 (0) | 14 (93.3) | 1 (6.7) | 0 (0) | 0 (0) | 11 | 2 | ||||
| Hmong | 31 (0) | 27 (87.1) | 2 (6.5) | 2 (6.5) | 0 (0) | 15 | 5 | ||||
| Other | 11 (1) | 6 (50) | 4 (33.3) | 1 (8.3) | 1 (8.3) | 8 | 0 |
| Dog Population | Haplotype Diversity | Nucleotide Diversity |
|---|---|---|
| Serra da Estrela [19] | 0.8520 ± 0.0299 | 0.016842 ± 0.008789 |
| Portuguese sheepdog [19] | 0.4841 ± 0.1094 | 0.008291 ± 0.004644 |
| German Shepherd [22] | 0.6842 ± 0.0917 | 0.008681 ± 0.004928 |
| Maltese dog [23] | 0.8046 ± 0.0697 | 0.011758 ± 0.006335 |
| Kangal [2,20] | 0.8407 ± 0.0274 | 0.015272 ± 0.007852 |
| Tibetan mastiff [10,22] | 0.8063 ± 0.0181 | 0.006645 ± 0.003724 |
| Shiba [2,24] | 0.8161 ± 0.0447 | 0.012221 ± 0.006563 |
| Jindo [2,10] | 0.7308 ± 0.0640 | 0.006645 ± 0.003762 |
| Pungsan [2,10] | 0.9064 ± 0.0244 | 0.011367 ± 0.006090 |
| Thai dog [2,10] | 0.9493 ± 0.0124 | 0.009599 ± 0.005145 |
| Vietnamese dog | 0.9210 ± 0.0065 | 0.014819 ± 0.007586 |
| Population | n | No. of Haplotype E Individuals | No. of Individuals |
|---|---|---|---|
| Jindo | 53 | 1 | 3 (6%) |
| Pungsan | 40 | 1 | 3 (7.5%) |
| Shiba | 30 | 1 | 1 (3.33%) |
| Thai village dog | 105 | 2 | 4 (3.8%) |
| Vietnamese village dog | 429 | 2 | 35 (8.2%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thai, Q.K.; Nguyen, T.-C.; Le, C.-T.; Chung, A.-D.; Nguyen, T.M.-L.; Tran, Q.-D.; Savolainen, P.; Quan, Q.-D.; Tran, D.-L.; Tran, H.-D. HV1 mtDNA Reveals the High Genetic Diversity and the Ancient Origin of Vietnamese Dogs. Animals 2023, 13, 1036. https://doi.org/10.3390/ani13061036
Thai QK, Nguyen T-C, Le C-T, Chung A-D, Nguyen TM-L, Tran Q-D, Savolainen P, Quan Q-D, Tran D-L, Tran H-D. HV1 mtDNA Reveals the High Genetic Diversity and the Ancient Origin of Vietnamese Dogs. Animals. 2023; 13(6):1036. https://doi.org/10.3390/ani13061036
Chicago/Turabian StyleThai, Quan Ke, Thanh-Cong Nguyen, Cong-Trieu Le, Anh-Dung Chung, Tran Minh-Ly Nguyen, Quoc-Dung Tran, Peter Savolainen, Quoc-Dang Quan, Dai-Long Tran, and Hoang-Dung Tran. 2023. "HV1 mtDNA Reveals the High Genetic Diversity and the Ancient Origin of Vietnamese Dogs" Animals 13, no. 6: 1036. https://doi.org/10.3390/ani13061036