

Effects of Dietary Vegetable Oil Mixtures including Soybean Oil on Intestinal Oxidative Stress in Gilthead Sea Bream (Sparus aurata)
 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Fish and Feeding Trial
2.3. Sampling Procedures and Samples Preparations
2.4. Oxidative Stress Markers Analysis
2.5. RNA Extraction and cDNA Synthesis
2.6. Real-Time Quantitave-PCR (qPCR)
2.7. Statistical Analyses
3. Results
3.1. Lipid Peroxidation
3.2. Antioxidant Enzyme Activity
3.3. Gene Expression
3.4. Antioxidant Ratios
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of Food and Agriculture 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- Glencross, B.D. Exploring the nutritional demand for essential fatty acids by aquaculture species. Aquac. Res. 2009, 48, 71–124. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Fish and Shrimp; The Nacional Academies Press: Whasington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Caballero, M.J.; Gallardo, G.; Robaina, L.; Montero, D.; Fernández, A.; Izquierdo, M. Vegetable lipid sources in vitro biosynthesis of triacylglycerols and phospholipids in the intestine of sea bream (Sparus aurata). Br. J. Nutr. 2006, 95, 448–454. [Google Scholar] [CrossRef] [Green Version]
- Torrecillas, S.; Mompel, D.; Caballero, M.J.; Montero, D.; Merrifield, D.; Rodiles, A.; Robaina, L.; Zamorano, M.J.; Karalazos, V.; Kaushik, S.; et al. Effect of fishmeal and fish oil replacement by vegetable meals and oils on gut health of European sea bass (Dicentrarchus labrax). Aquaculture 2017, 468, 386–398. [Google Scholar] [CrossRef]
- Tarricone, S.; Iaffaldano, N.; Colonna, M.A.; Giannico, F.; Selvaggi, M.; Caputi Jambrenghi, A.; Cariglia, M.; Ragni, M. Effects of dietary red grape extract n the quality traits in juvenile European sea bass (Dicentrarchus labrax L.). Animals 2023, 13, 254. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M. Welfare and health of fish fed vegetable oils as alternative lipid sources to fish oil. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G., Ng, W.-K., Tocher, D.R., Eds.; CRC Press: Boca Raton, FL, USA, 2010; p. 522. ISBN 978-1-4398-0862-7. [Google Scholar]
- Benedito-Palos, L.; Ballester-Lozano, G.F.; Simó, P.; Karalazos, V.; Ortiz, Á.; Calduch-Giner, J.; Pérez-Sánchez, J. Lasting effects of butyrate and low FM/FO diets on growth performance, blood haematology/biochemistry and molecular growth-related markers in gilthead sea bream (Sparus aurata). Aquaculture 2016, 454, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Simó-Mirabet, P.; Felip, A.; Estensoro, I.; Martos-Sitcha, J.A.; de las Heras, V.; Calduch-Giner, J.; Puyalto, M.; Karalazos, V.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Impact of low fish meal and fish oil diets on the performance, sex steroid profile and male-female sex reversal of gilthead sea bream (Sparus aurata) over a three-year production cycle. Aquaculture 2018, 490, 64–74. [Google Scholar] [CrossRef]
- Gil-Solsona, R.; Calduch-Giner, J.A.; Nácher-Mestre, J.; Lacalle-Bergeron, L.; Sancho, J.V.; Hernández, F.; Pérez-Sánchez, J. Contributions of MS metabolomics to gilthead sea bream (Sparus aurata) nutrition. Serum fingerprinting of fish fed low fish meal and fish oil diets. Aquaculture 2019, 498, 503–512. [Google Scholar] [CrossRef]
- Morais, S.; Silva, T.; Cordeiro, O.; Rodrigues, P.; Guy, D.R.; Bron, J.E.; Taggart, J.B.; Bell, J.G.; Tocher, D.R. Effects of genotype and dietary fish oil replacement with vegetable oil on the intestinal transcriptome and proteome of Atlantic salmon (Salmo salar). BMC Genom. 2012, 13, 448. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.E.; Myklebust, R.; Kaino, T.; Ringø, E. Lipid digestibility and ultrastructural changes in the enterocytes of Arctic char (Salvelinus alpinus L.) fed linseed oil and soybean lecithin. Fish Physiol. Biochem. 1999, 21, 35–44. [Google Scholar] [CrossRef]
- Caballero, M.J.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Olsen, R.E.; Dragnes, B.T.; Myklebust, R.; Ringø, E. Effect of soybean oil and soybean lecithin on intestinal lipid composition and lipid droplet accumulation of rainbow trout, Oncorhynchus mykiss Walbaum. Fish Physiol. Biochem. 2003, 29, 181–192. [Google Scholar] [CrossRef]
- Caballero, M.J.; Izquierdo, M.S.; Kjørsvik, E.; Montero, D.; Socorro, J.; Fernández, A.J.; Rosenlund, G. Morphological aspects of intestinal cells from gilthead seabream (Sparus aurata) fed diets containing different lipid sources. Aquaculture 2003, 225, 325–340. [Google Scholar] [CrossRef]
- Ruyter, B.; Moya-Falcón, C.; Rosenlund, G.; Vegusdal, A. Fat content and morphology of liver and intestine of Atlantic salmon (Salmo salar): Effects of temperature and dietary soybean oil. Aquaculture 2006, 252, 441–452. [Google Scholar] [CrossRef]
- Bowyer, J.N.; Rout-Pitt, N.; Bain, P.A.; Stone, D.A.J.; Schuller, K.A. Dietary fish oil replacement with canola oil up-regulates glutathione peroxidase 1 gene expression in Yellowtail kingfish (Seriola lalandi). Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2012, 162, 100–106. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Xiao, W.; Jiang, W.; Feng, L.; Liu, Y.; Wu, P.; Jiang, J.; Zhang, Y.; Zhou, X. Effect of dietary enzyme-treated soy protein on the immunity and antioxidant status in the intestine of juvenile Jian carp (Cyprinus carpio Var. Jian). Aquac. Res. 2019, 50, 1411–1421. [Google Scholar] [CrossRef]
- Castro, C.; Diógenes, A.F.; Coutinho, F.; Panserat, S.; Corraze, G.; Pérez-Jiménez, A.; Peres, H.; Oliva-Teles, A. Liver and intestine oxidative status of gilthead sea bream fed vegetable oil and carbohydrate rich diets. Aquaculture 2016, 464, 665–672. [Google Scholar] [CrossRef]
- Kiron, V.; Thawonsuwan, J.; Panigrahi, A.; Scharsack, J.P.; Satoh, S. Antioxidant and immune defences of rainbow trout (Oncorhynchus mykiss) offered plant oils differing in fatty acid profiles from early stages. Aquac. Nutr. 2011, 17, 130–140. [Google Scholar] [CrossRef]
- Kjær, M.A.; Aursnes, I.A.; Berge, G.M.; Sørensen, M.; Marchenko, Y.; Gjøen, T.; Ruyter, B. The influence of different dietary oil qualities on growth rate, feed utilization and oxidative stress in Atlantic cod. Aquac. Nutr. 2014, 20, 192–204. [Google Scholar] [CrossRef]
- Mourente, G.; Díaz-Salvago, E.; Bell, J.G.; Tocher, D.R. Increased activities of hepatic antioxidant defence enzymes in juvenile gilthead sea bream (Sparus aurata L.) fed dietary oxidised oil: Attenuation by dietary vitamin E. Aquaculture 2002, 214, 343–361. [Google Scholar] [CrossRef]
- Bresciani, G.; da Cruz, I.B.M.; González-Gallego, J. Manganese superoxide dismutase and oxidative stress modulation. In Advances in Clinical Chemistry; Makoski, G., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 89–114. [Google Scholar]
- Bacchetta, C.; Rossi, A.S.; Cian, R.E.; Drago, S.R.; Cazenave, J. Dietary β-carotene improves growth performance and antioxidant status of juvenile Piaractus mesopotamicus. Aquac. Nutr. 2019, 25, 761–769. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, X.Y.; Zhou, X.Q.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Jiang, J.; Shi, D.; Zhou, X.Q.; Yin, L.; Feng, L.; Liu, Y.; Jiang, W.D.; Zhao, Y. Effects of glutamate on growth, antioxidant capacity, and antioxidant-related signaling molecule expression in primary cultures of fish enterocytes. Fish Physiol. Biochem. 2015, 41, 1143–1153. [Google Scholar] [CrossRef]
- Magalhães, R.; Guerreiro, I.; Santos, R.A.; Coutinho, F.; Couto, A.; Serra, C.R.; Olsen, R.E.; Peres, H.; Oliva-Teles, A. Oxidative status and intestinal health of gilthead sea bream (Sparus aurata) juveniles fed diets with different ARA/EPA/DHA ratios. Sci. Rep. 2020, 10, 13824. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, H.; Mokrani, A.; Ji, K.; Yu, H.; Ge, X.; Ren, M.; Xie, J.; Pan, L.; Sun, A. Dietary histidine affects intestinal antioxidant enzyme activities, antioxidant gene expressions and inflammatory factors in juvenile Blunt snout bream (Megalobrama amblycephala). Aquac. Nutr. 2019, 25, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Jiang, W.D.; Duan, X.D.; Feng, L.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Soybean glycinin caused NADPH-oxidase-regulated ROS overproduction and decreased ROS elimination capacity in the mid and distal intestine of juvenile grass carp (Ctenopharyngodon idella). Aquaculture 2020, 516, 734651. [Google Scholar] [CrossRef]
- Puangkaew, J.; Kiron, V.; Somamoto, T.; Okamoto, N.; Satoh, S.; Takeuchi, T.; Watanabe, T. Nonspecific immune response of rainbow trout (Oncorhynchus mykiss Walbaum) in relation to different status of vitamin E and highly unsaturated fatty acids. Fish Shellfish Immunol. 2004, 16, 25–39. [Google Scholar] [CrossRef]
- Østbye, T.K.; Kjær, M.A.; Rørå, A.M.B.; Torstensen, B.; Ruyter, B. High n-3 HUFA levels in the diet of Atlantic salmon affect muscle and mitochondrial membrane lipids and their susceptibility to oxidative stress. Aquac. Nutr. 2011, 17, 177–190. [Google Scholar] [CrossRef]
- Sánchez-Moya, A.; García-Meilán, I.; Riera-Heredia, N.; Vélez, E.J.; Lutfi, E.; Fontanillas, R.; Gutiérrez, J.; Capilla, E.; Navarro, I. Effects of different dietary vegetable oils on growth and intestinal performance, lipid metabolism and flesh quality in gilthead sea bream. Aquaculture 2020, 519, 734881. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Mccords, J.M.; Fridovich, I. Superoxide dismutase an enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar]
- Aebi, H. [13] Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.G.; Pirie, B.J.S.; Adron, J.W.; Cowey, C.B. Some effects of selenium deficiency on glutathione peroxidase (EC 1.11.1.9) activity and tissue pathology in rainbow trout (Salmo gairdneri). Br. J. Nutr. 1986, 55, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Habig, W.-H.; Pabst, M.J.; Fleischnert, G.; Gatmaitant, Z.; Ariast, I.M.; Jakoby, W.B. The identity of glutathione S-transferase B with ligandin, a major binding protein of liver (organic anion binding protein/Y protein). Proc. Nat. Acad. Sci. USA 1974, 71, 3879–3882. [Google Scholar] [CrossRef] [Green Version]
- Carlberg, I.; Mannervik, B. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem. 1975, 260, 5475–5480. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum information for publication of quantitative Real-Time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Guardiola, F.A.; Bahi, A.; Messina, C.M.; Mahdhi, A.; Santulli, A.; Arena, R.; Bakhrouf, A.; Esteban, M.A. Quality and antioxidant response of gilthead seabream (Sparus aurata L.) to dietary supplements of fenugreek (Trigonella foenum graecum) alone or combined with probiotic strains. Fish Shellfish Immunol. 2017, 63, 277–284. [Google Scholar] [CrossRef]
- Mansour, A.T.; Espinosa, C.; García-Beltrán, J.M.; Miao, L.; Ceballos Francisco, D.C.; Alsaqufi, A.S.; Esteban, M.Á. Dietary supplementation of Drumstick tree, Moringa oleifera, improves mucosal immune response in skin and gills of seabream, Sparus aurata, and attenuates the effect of hydrogen peroxide exposure. Fish Physiol. Biochem. 2020, 46, 981–996. [Google Scholar] [CrossRef]
- Pérez-Sánchez, J.; Borrel, M.; Bermejo-Nogales, A.; Benedito-Palos, L.; Saera-Vila, A.; Calduch-Giner, J.A.; Kaushik, S. Dietary oils mediate cortisol kinetics and the hepatic mRNA expression profile of stress-responsive genes in gilthead sea bream (Sparus aurata) exposed to crowding stress. Implications on energy homeostasis and stress susceptibility. Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- de Barboza, G.D.; Guizzardi, S.; Moine, L.; Tolosa de Talamoni, N. Oxidative stress, antioxidants and intestinal calcium absorption. World J. Gastroenterol. 2017, 23, 2841–2853. [Google Scholar] [CrossRef] [PubMed]
- Benedito-Palos, L.; Navarro, J.C.; Sitjà-Bobadilla, A.; Gordon Bell, J.; Kaushik, S.; Pérez-Sánchez, J. High levels of vegetable oils in plant protein-rich diets fed to gilthead sea bream (Sparus aurata L.): Growth performance, muscle fatty acid profiles and histological alterations of target tissues. Br. J. Nutr. 2008, 100, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.S.; Obach, A.; Arantzamendi, L.; Montero, D.; Robaina, L.; Rosenlund, G. Dietary lipid sources for seabream and seabass: Growth performance, tissue composition and flesh quality. Aquac. Nutr. 2003, 9, 397–407. [Google Scholar] [CrossRef]
- Guerreiro, I.; Couto, A.; Pérez-Jiménez, A.; Oliva-Teles, A.; Enes, P. Gut morphology and hepatic oxidative status of European sea bass (Dicentrarchus labrax) juveniles fed plant feedstuffs or fishmeal-based diets supplemented with short-chain fructo-oligosaccharides and xylo-oligosaccharides. Br. J. Nutr. 2015, 114, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Castro, C.; Coutinho, F.; Iglesias, P.; Oliva-Teles, A.; Couto, A. Chlorella sp. and Nannochloropsis sp. inclusion in plant-based diets modulate the intestine and liver antioxidant mechanisms of European sea bass juveniles. Front. Vet. Sci. 2020, 7, 607575. [Google Scholar] [CrossRef]
- Coutinho, F.; Castro, C.; Rufino-Palomares, E.; Ordóñez-Grande, B.; Gallardo, M.A.; Oliva-Teles, A.; Peres, H. Dietary glutamine supplementation effects on amino acid metabolism, intestinal nutrient absorption capacity and antioxidant response of gilthead sea bream (Sparus aurata) juveniles. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 191, 9–17. [Google Scholar] [CrossRef]
- Halliwell, B.; Zhao, K.; Whiteman, M. The gastrointestinal tract: A major site of antioxidant action? Free Radic. Res. 2000, 33, 819–830. [Google Scholar] [CrossRef]
- Box, A.; Sureda, A.; Galgani, F.; Pons, A.; Deudero, S. Assessment of environmental pollution at Balearic islands applying oxidative stress biomarkers in the mussel Mytilus galloprovincialis. Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2007, 146, 531–539. [Google Scholar] [CrossRef]
- Bakke, A.M.; Glover, C.; Krogdahl, A. Feeding, digestion and absorption of nutrients. In Multifunctional Gut of Fish; Grosell, M., Farrell, A., Brauner, C., Eds.; Academic Press: Cambridge, MA, USA, 2011; pp. 57–75. [Google Scholar]
- Verdile, N.; Pasquariello, R.; Scolari, M.; Scirè, G.; Brevini, T.A.L.; Gandolfi, F. A detailed study of rainbow trout (Onchorhynchus mykiss) intestine revealed that digestive and absorptive functions are not linearly distributed along its length. Animals 2020, 10, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rønnestad, I.; Rojas-Garcia, C.R.; Skadal, J. Retrograde peristalsis; a possible mechanism for filling the pyloric caeca? J. Fish Biol. 2000, 56, 216–218. [Google Scholar] [CrossRef]
- Jones, B.S.; Keightley, L.J.; Harris, J.O.; Wiklendt, L.; Spencer, N.J.; Dinning, P.G. Identification of neurogenic intestinal motility patterns in Silver perch (Bidyanus bidyanus) that persist over wide temperature ranges. Neurogastroenterol. Motil. 2021, 33, e14037. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Noh, H.; Numayama-Tsuruta, K.; Ishikawa, T. Mechanical roles of anterograde and retrograde intestinal peristalses after feeding in a larval fish (Danio rerio). Am. J. Physiol. Gastrointest. Liver. Physiol. 2020, 318, G1013–G1021. [Google Scholar] [CrossRef]
- García-Meilán, I.; Valentín, J.M.; Fontanillas, R.; Gallardo, M.A. Different protein to energy ratio diets for gilthead sea bream (Sparus aurata): Effects on digestive and absorptive processes. Aquaculture 2013, 412–413, 1–7. [Google Scholar] [CrossRef]
- García-Meilán, I.; Ordóñez-Grande, B.; Gallardo, M.A. Meal timing affects protein-sparing effect by carbohydrates in sea bream: Effects on digestive and absorptive processes. Aquaculture 2014, 434, 121–128. [Google Scholar] [CrossRef]
- García-Meilán, I.; Ordóñez-Grande, B.; Machahua, C.; Buenestado, S.; Fontanillas, R.; Gallardo, M.A. Effects of dietary protein-to-lipid ratio on digestive and absorptive processes in sea bass fingerlings. Aquaculture 2016, 463, 163–173. [Google Scholar] [CrossRef]
- García-Meilán, I.; Ordóñez-Grande, B.; Valentín, J.M.; Hernández, M.D.; García, B.; Fontanillas, R.; Gallardo, M.A. Modulation of digestive and absorptive processes with age and/or after a dietary change in gilthead sea bream. Aquaculture 2016, 459, 54–64. [Google Scholar] [CrossRef]
- García-Meilán, I.; Ordóñez-Grande, B.; Valentín, J.M.; Fontanillas, R.; Gallardo, M.A. High dietary carbohydrate inclusión by both protein and lipid replacement in gilthead sea bream. Changes in digestive and absorptive processes. Aquaculture 2020, 520, 734977. [Google Scholar] [CrossRef]
- Miegel, R.P.; Pain, S.J.; van Wettere, W.H.E.J.; Howarth, G.S.; Stone, D.A.J. Effect of water temperature on gut transit time, digestive enzyme activity and nutrient digestibility in Yellowtail kingfish (Seriola lalandi). Aquaculture 2010, 308, 145–151. [Google Scholar] [CrossRef]
- Guillaume, J.; Choubert, G. Digestive physiology and nutrient digestibility in fishes. In Nutrition and Feeding of Fish and Crustaceans; Guillaume, J., Ed.; Springer Praxis: Chichester, UK, 2001. [Google Scholar]
- Bjørgen, H.; Koppang, E.O. Anatomy of teleost fish immune structures and organs. Immunogenetics 2021, 73, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Chikwati, E.M.; Sahlmann, C.; Holm, H.; Penn, M.H.; Krogdahl, Å.; Bakke, A.M. Alterations in digestive enzyme activities during the development of diet-induced enteritis in Atlantic salmon, Salmo salar L. Aquaculture 2013, 402–403, 28–37. [Google Scholar] [CrossRef]
- Venold, F.F.; Penn, M.H.; Thorsen, J.; Gu, J.; Kortner, T.M.; Krogdahl, Å.; Bakke, A.M. Intestinal fatty acid binding protein (Fabp2) in Atlantic salmon (Salmo salar): Localization and alteration of expression during development of diet induced enteritis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 164, 229–240. [Google Scholar] [CrossRef]
- Kortner, T.M.; Skugor, S.; Penn, M.H.; Mydland, L.T.; Djordjevic, B.; Hillestad, M.; Krasnov, A.; Krogdahl, Å. Dietary soyasaponin supplementation to pea protein concentrate reveals nutrigenomic interactions underlying enteropathy in Atlantic salmon (Salmo salar). BMC Vet. Res. 2012, 8, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Diets | |||||
| Ingredients (%) | S | SL | SLR | SLP | SLRP |
| Fish oil | 4.64 | 4.64 | 4.64 | 4.64 | 4.64 |
| Palm oil | - | - | - | 4.81 | 2.43 |
| Linseed oil | - | 2.82 | 2.42 | 3.37 | 2.7 |
| Rapeseed oil | - | - | 5.80 | - | 5.84 |
| Soybean oil | 13.63 | 10.81 | 5.39 | 5.54 | 2.65 |
| Vit/Min premix | 1.78 | 1.78 | 1.8 | 1.69 | 1.79 |
| Proximate composition (%) | |||||
| Dry matter | 93.21 | 93.21 | 93.21 | 93.21 | 93.21 |
| Crude protein | 46.5 | 46.5 | 46.50 | 46.45 | 46.33 |
| Crude fat | 22.4 | 21.9 | 22.40 | 22.40 | 21.90 |
| Ash | 5.7 | 5.7 | 5.69 | 5.66 | 5.66 |
| Diets | |||||
|---|---|---|---|---|---|
| Fatty Acid (%) | S | SL | SLR | SLP | SLRP |
| C14:0 | 2.15 | 2.12 | 2.14 | 2.33 | 2.27 |
| C16:0 | 13.27 | 12.55 | 10.95 | 18.98 | 14.17 |
| C16:1n-7 | 2.34 | 2.31 | 2.37 | 2.36 | 2.40 |
| C16:2n-6 | 0.29 | 0.28 | 0.30 | 0.28 | 0.29 |
| C18:0 | 2.87 | 3.16 | 2.80 | 3.51 | 2.99 |
| C18:1n-9 | 19.88 | 19.19 | 27.30 | 23.02 | 29.81 |
| C18:1n-7 | 2.03 | 1.88 | 2.24 | 1.62 | 2.15 |
| C18:2n-6 | 36.51 | 31.95 | 24.93 | 21.98 | 19.10 |
| C18:3n-3 | 4.37 | 10.11 | 10.11 | 9.83 | 10.04 |
| C18:4n-3 | 0.75 | 0.73 | 0.73 | 0.70 | 0.74 |
| C20:1 sum. isomers | 1.80 | 1.85 | 2.15 | 1.70 | 2.05 |
| C20:4n-6 | 0.29 | 0.23 | 0.22 | 0.23 | 0.22 |
| C20:4n-3 | 0.22 | 0.23 | 0.22 | 0.25 | 0.24 |
| C20:5n-3 EPA | 3.02 | 3.02 | 3.10 | 3.02 | 3.15 |
| C22:1 sum. isomers | 2.13 | 2.06 | 2.24 | 2.18 | 2.17 |
| C22:5n-3 | 0.44 | 0.43 | 0.46 | 0.43 | 0.46 |
| C22:6n-3 DHA | 2.98 | 2.87 | 2.99 | 2.95 | 2.95 |
| C24:1n-9 | 0.25 | 0.23 | 0.27 | 0.27 | 0.26 |
| SFA not listed | 1.06 | 0.97 | 1.01 | 0.87 | 0.96 |
| Monoenes not listed | 0.10 | 0.14 | 0.12 | 0.10 | 0.11 |
| n-6 FA not listed | 0.20 | 0.24 | 0.25 | 0.21 | 0.22 |
| n-3 FA not listed | 0.22 | 0.20 | 0.20 | 0.17 | 0.20 |
| Others | 0.36 | 0.36 | 0.32 | 0.32 | 0.36 |
| Sum. SFA | 19.35 | 18.80 | 16.90 | 25.69 | 20.39 |
| Sum. MUFA | 28.53 | 27.66 | 36.69 | 31.25 | 38.95 |
| Sum. n-6 FA | 37.29 | 32.70 | 25.70 | 22.70 | 19.83 |
| Sum. n-3 FA | 12.0 | 17.59 | 17.81 | 17.35 | 17.78 |
| UFA/SFA | 4.02 | 4.15 | 4.75 | 2.78 | 3.75 |
| MUFA/PUFA | 0.58 | 0.55 | 0.84 | 0.78 | 1.04 |
| n-6/n-3 | 3.11 | 1.86 | 1.44 | 1.31 | 1.12 |
| Unknown | 2.50 | 2.90 | 2.60 | 2.70 | 2.70 |
| Gene | Sequence (5′-3′) | Tm (°C) | Accession Number | |
|---|---|---|---|---|
| β-actin | F R | TCCTGCGGAATCCATGAGA GACGTCGCACTTCATGATGCT | 60 | X89920 |
| ef1α | F R | CTTCAACGCTCAGGTCATCAT GCACAGCGAAACGACCAAGGGGA | 60 | AF184170 |
| rps18 | F R | GGGTGTTGGCAGACGTTAC CTTCTGCCTGTTGAGGAACCA | 60 | AM490061.1 |
| cat | F R | TTCCCGTCCTTCCATTCACTC CTCCAGAAGTCCCACACCAT | 60 | FG264808 |
| gpx1 | F R | GAAGGTGGATGTGAATGGAAAAGATG CTGACGGGACTCCAAATGATGG | 60 | DQ524992 |
| gpx4 | F R | TGCGTCTGATAGGGTCCACTGTC GTCTGCCAGTCCTCTGTCGG | 60 | AM977818 |
| gr | F R | CAAAGCGCAGTGTGATTGTGG CCACTCCGGAGTTTTGCATTTC | 60 | AJ937873 |
| gst3 | F R | CCAGATGATCAGTACGTGAAGACCGTC TGCTGATGTGAGGAATGTACCGTAAC | 60 | JQ308828 |
| sod1 | F R | CCATGGTAAGAATCATGGCGG CGTGGATCACCATGGTTCTG | 60 | AJ937872 |
| sod2 | F R | CCTGACCTGACCTACGACTATGG AGTGCCTCCTGATATTTCTCCTCTG | 60 | JQ308832 |
| Dietary Conditions | Pyloric Caeca | Proximal Intestine |
|---|---|---|
| S | 0.057 ± 0.006 c | 0.318 ± 0.039 mn* |
| SL | 2.907 ± 1.109 | 1.097 ± 0.502 |
| L-SL | 0.107 ± 0.006 b | 0.201 ± 0.015 n* |
| H-SL | 9.440 ± 2.750 a* | 2.889.2 ± 1.407 m |
| SLR | 0.139 ± 0.017 b | 0.643 ± 0.122 m* |
| SLP | 0.038 ± 0.005 d | 0.265 ± 0.019 no* |
| SLRP | 0.136 ± 0.022 b | 0.227 ± 0.011 o |
| Pyloric Caeca | |||||
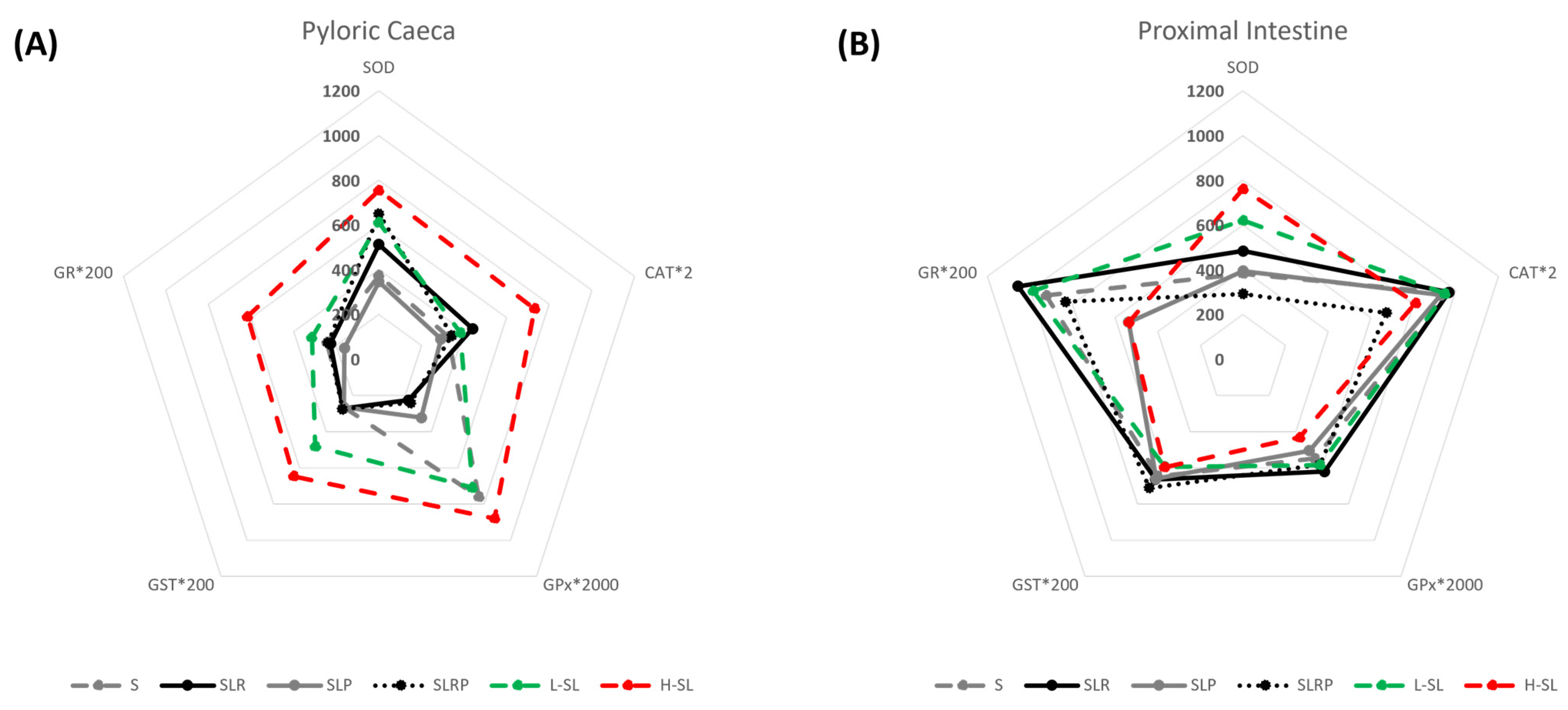
| Dietary Conditions | SOD | CAT | GPx | GST | GR |
| S | 374.6 ± 35.0 c | 163.0 ± 11.3 bc | 0.380 ± 0.111 ab* | 1.33 ± 0.11 b | 1.22 ± 0.10 b |
| L-SL | 611.6 ± 93.9 ab | 191.9 ± 9.0 ab | 0.355 ± 0.043 a | 2.41 ± 0.22 a | 1.57 ± 0.17 bc |
| H-SL | 754.4 ± 173 a | 365.8 ± 69.4 a | 0.440 ± 0.060 a* | 3.23 ± 0.57 a | 3.08 ± 0.34 a |
| SLR | 513.2 ± 41.8 b | 220.3 ± 4.4 a | 0.113 ± 0.009 c | 1.36 ± 0.06 b | 1.13 ± 0.09 b |
| SLP | 345.8 ± 43.3 c | 145.5 ± 6.2 c | 0.161 ± 0.012 b | 1.33 ± 0.06 b | 0.80 ± 0.07 c |
| SLRP | 650.5 ± 44.3 ab* | 171.6 ± 9.1 b | 0.121 ± 0.008 c | 1.38 ± 0.06 b | 1.19 ± 0.11 b |
| Proximal intestine | |||||
| Dietary conditions | SOD | CAT | GPx | GST | GR |
| S | 381.9 ± 33.4 op | 479.5 ± 16.3 m* | 0.274 ± 0.030 | 3.23 ± 0.22 mn* | 4.62 ± 0.45 m* |
| L-SL | 620.7 ± 72.8 m | 474.5 ± 42.5 m* | 0.292 ± 0.035 | 2.97 ± 0.12 n | 4.93 ± 0.46 m* |
| H-SL | 760.5 ± 151.5 m | 405.3 ± 18.9 m | 0.216 ± 0.019 | 2.99 ± 0.17 mn | 2.68 ± 0.26 n |
| SLR | 483.7 ± 56.62 mn | 483.5 ± 32.2 m* | 0.310 ± 0.030 * | 3.32 ± 0.19 mn* | 5.28 ± 0.66 m* |
| SLP | 394.4 ± 40.24 no | 464.4 ± 27.3 m* | 0.252 ± 0.018 * | 3.30 ± 0.15 mn* | 2.68 ± 0.12 n* |
| SLRP | 292.0 ± 29.53 p | 336.7 ± 10.5 n* | 0.292 ± 0.024 * | 3.55 ± 0.20 m* | 4.16 ± 0.29 m* |
| Pyloric Caeca | |||||||
| Dietary Conditions | sod1 | sod2 | cat | gpx1 | gpx4 | gst | gr |
| S | 2.15 ± 0.52 | 3.09 ± 1.00 a | 2.42 ± 0.74 | 1.91 ± 0.38 a | 3.32 ± 0.94 a | 1.70 ± 0.47 | 1.35 ± 0.43 |
| L-SL | 1.60 ± 0.49 | 2.05 ± 0.53 a | 1.47 ± 0.29 | 1.01 ± 0.22 b | 3.24 ± 2.05 ab | 0.89 ± 0.25 | 1.51 ± 0.74 |
| H-SL | 2.30 ± 1.08 | 3.60 ± 1.76 a | 3.13 ± 1.79 | 2.19 ± 0.98 ab | 4.05 ± 2.85 ab | 1.88 ± 0.74 | 1.98 ± 1.05 |
| SLR | 1.63 ± 0.39 | 1.79 ± 0.53 ab | 1.63 ± 0.27 | 1.43 ± 0.19 ab | 1.26 ± 0.30 bc | 1.51 ± 0.45 | 1.17 ± 0.25 |
| SLP | 0.57 ± 0.17 | 0.55 ± 0.17 b | 0.67 ± 0.22 | 0.37 ± 0.11 c | 0.45 ± 0.14 c | 0.63 ± 0.22 | 0.51 ± 0.20 |
| SLRP | 1.65 ± 0.27 | 1.55 ± 0.21 a | 1.26 ± 0.17 | 1.58 ± 0.32 ab | 1.29 ± 0.20 ab | 1.45 ± 0.28 | 1.36 ± 0.19 |
| Proximal intestine | |||||||
| Dietary conditions | sod1 | sod2 | cat | gpx1 | gpx4 | gst | gr |
| S | 0.87 ± 0.09 | 1.72 ± 0.19 | 0.87 ± 0.21 | 2.04 ± 0.20 | 0.92 ± 0.12 m | 1.04 ± 0.08 | 1.73 ± 0.19 |
| L-SL | 0.87 ± 0.20 | 1.87 ± 0.37 | 0.86 ± 0.09 | 1.79 ± 0.14 | 0.55 ± 0.04 o | 0.90 ± 0.30 | 1.95 ± 0.42 |
| H-SL | 0.69 ± 0.14 | 2.80 ± 1.21 | 1.01 ± 0.28 | 2.97 ± 1.81 | 0.65 ± 0.16 mno | 0.91 ± 0.14 | 3.54 ± 1.83 |
| SLR | 0.85 ± 0.10 | 1.80 ± 0.22 | 1.20 ± 0.18 | 1.93 ± 0.09 | 0.87 ± 0.06 m | 1.24 ± 0.25 | 2.17 ± 0.26 |
| SLP | 0.75 ± 0.21 | 1.60 ± 0.15 | 0.82 ± 0.15 | 1.69 ± 0.20 | 0.73 ± 0.06 mn | 0.97 ± 0.10 | 1.80 ± 0.25 |
| SLRP | 0.75 ± 0.11 | 1.44 ± 0.20 | 0.72 ± 0.11 | 1.80 ± 0.21 | 0.55 ± 0.08 no | 0.88 ± 0.16 | 1.79 ± 0.14 |
| Pyloric Caeca | |||
| Dietary Conditions | (CAT+GPx)/SOD | CAT/GPx | (GST+GPx)/GR |
| S | 0.468 ± 0.036 a | 569.8 ± 95.1 e | 1.566 ± 0.126 ab* |
| L-SL | 0.272 ± 0.017 b | 598.0 ± 40.0 e | 1.924 ± 0.21 a* |
| H-SL | 0.497 ± 0.041 a | 771.9 ± 47.5 d | 1.082 ± 0.116 c |
| SLR | 0.480 ± 0.047 a | 2289.3 ± 215.8 a* | 1.326 ± 0.101 bc* |
| SLP | 0.580 ± 0.096 a | 1043.5 ± 98.4 c | 1.982 ± 0.142 a* |
| SLRP | 0.311 ± 0.029 b | 1488.8 ± 67.0 b | 1.378 ± 0.166 abc |
| Proximal intestine | |||
| Dietary conditions | (CAT+GPx)/SOD | CAT/GPx | (GST+GPx)/GR |
| S | 1.388 ± 0.098 m* | 1997.4 ± 218.1 mn* | 0.693 ± 0.014 o |
| L-SL | 0.645 ± 0.023 o* | 1781.5 ± 303.5 mn* | 0.576 ± 0.015 o |
| H-SL | 0.730 ± 0.150 no | 2008.2 ± 240.1 mn* | 1.226 ± 0.053 m |
| SLR | 0.916 ± 0.038 n* | 1569.8 ± 148.7 n | 0.914 ± 0.090 n |
| SLP | 1.443 ± 0.121 m* | 1956.1 ± 125.4 m* | 1.399 ± 0.022 m |
| SLRP | 1.351 ± 0.115 m* | 1432.9 ± 101.3 n | 1.095 ± 0.023 n |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Meilán, I.; Fontanillas, R.; Gutiérrez, J.; Capilla, E.; Navarro, I.; Gallardo, Á. Effects of Dietary Vegetable Oil Mixtures including Soybean Oil on Intestinal Oxidative Stress in Gilthead Sea Bream (Sparus aurata). Animals 2023, 13, 1069. https://doi.org/10.3390/ani13061069
García-Meilán I, Fontanillas R, Gutiérrez J, Capilla E, Navarro I, Gallardo Á. Effects of Dietary Vegetable Oil Mixtures including Soybean Oil on Intestinal Oxidative Stress in Gilthead Sea Bream (Sparus aurata). Animals. 2023; 13(6):1069. https://doi.org/10.3390/ani13061069
Chicago/Turabian StyleGarcía-Meilán, Irene, Ramón Fontanillas, Joaquim Gutiérrez, Encarnación Capilla, Isabel Navarro, and Ángeles Gallardo. 2023. "Effects of Dietary Vegetable Oil Mixtures including Soybean Oil on Intestinal Oxidative Stress in Gilthead Sea Bream (Sparus aurata)" Animals 13, no. 6: 1069. https://doi.org/10.3390/ani13061069
APA StyleGarcía-Meilán, I., Fontanillas, R., Gutiérrez, J., Capilla, E., Navarro, I., & Gallardo, Á. (2023). Effects of Dietary Vegetable Oil Mixtures including Soybean Oil on Intestinal Oxidative Stress in Gilthead Sea Bream (Sparus aurata). Animals, 13(6), 1069. https://doi.org/10.3390/ani13061069