Simple Summary
Although glucocorticoids are frequently used for a variety of purposes, the effects and mechanisms of glucocorticoids on platelets are not fully understood. The present study investigates the effect of prednisolone and its mechanism of action on the regulation of platelet function. The results demonstrated that prednisolone affects platelet function by inhibiting thromboxane A2 (TxA2) generation through the regulation of cPLA2 phosphorylation, providing knowledge of glucocorticoids in coagulation and bleeding disorders.
Abstract
Glucocorticoids have been commonly used in the treatment of inflammation and immune-mediated diseases in human beings and small animals such as cats and dogs. However, excessive use can lead to Cushing’s syndrome along with several thrombotic and cardiovascular diseases. Although it is well-known that glucocorticoids exert a significant effect on coagulation, the effect of cortisol on platelet function is much less clear. Thus, we aimed to study the effects of prednisolone, one of the commonly used glucocorticoids, on the regulation of platelet function using murine platelets. We first evaluated the concentration-dependent effect of prednisolone on 2-MeSADP-induced platelet function and found that the 2-MeSADP-induced secondary wave of aggregation and dense granule secretion were completely inhibited from 500 nM prednisolone. Since 2-MeSADP-induced secretion and the resultant secondary wave of aggregation are mediated by TxA2 generation, this result suggested a role of prednisolone in platelet TxA2 generation. Consistently, prednisolone did not affect the 2-MeSADP-induced aggregation in aspirinated platelets, where the secondary wave of aggregation and secretion were blocked by eliminating the contribution of TxA2 generation by aspirin. In addition, thrombin-induced platelet aggregation and secretion were inhibited in the presence of prednisolone by inhibiting the positive-feedback effect of TxA2 generation on platelet function. Furthermore, prednisolone completely inhibited 2-MeSADP-induced TxA2 generation, confirming the role of prednisolone in TxA2 generation. Finally, Western blot analysis revealed that prednisolone significantly inhibited 2-MeSADP-induced cytosolic phospholipase A2 (cPLA2) and ERK phosphorylation in non-aspirinated platelets, while only cPLA2 phosphorylation, but not ERK phosphorylation, was significantly inhibited by prednisolone in aspirinated platelets. In conclusion, prednisolone affects platelet function by the inhibition of TxA2 generation through the regulation of cPLA2 phosphorylation, thereby shedding light on its clinical characterization and treatment efficacy in dogs with hypercortisolism in the future.
1. Introduction
Prednisolone is one of the most commonly used glucocorticoids in the treatment of various diseases including inflammatory and immune-mediated diseases. Prednisolone has the ability to reduce the recruitment of inflammatory cells including eosinophils, lymphocytes, mast cells, and dendritic cells by reducing the production of chemotactic mediators and adhesion molecules and the ability of inflammatory cells to survive [1]. Due to the high efficacy of glucocorticoids, the prescription rate has climbed high during the past years, leading to excessive glucocorticoid abuse that raises blood cortisol levels, resulting in iatrogenic Cushing’s syndrome [2,3,4,5].
Cushing’s syndrome is a disorder characterized by polyuria, polydipsia, muscle and skin atrophy, weight loss, truncal obesity, and hair loss [6]. Excessive use of glucocorticoids in thrombotic, hemostatic, and cardiovascular diseases is associated with increased morbidity and mortality [7]. Glucocorticoid medication has been shown to increase the incidence of hypertension, hyperglycemia, and hyperglyceridemia leading to coronary and ischemic heart disease, heart failure, and unexpected death [8,9]. Moreover, high levels of glucocorticoids have been shown to elevate levels of von Willebrand factor (vWF), anti-hemophilic factor, fibrinogen, plasminogen activator inhibitor-1, and platelet count [10,11,12], which can result in embolic disorder and further increase the risk of the coagulative disorder [13]. The pathogenesis and clinical symptoms of Cushing’s syndrome are similar in both humans and dogs. However, the incident rate is 1000 times higher in dogs compared to humans [6]. As a result, Cushing’s syndrome-induced thromboembolic complications are around four times more common in dogs which leads to a four times higher mortality rate, making it a disease of interest in veterinary medicine [14].
It has been well-established that platelets play a crucial role in thrombosis and hemostasis and have been known to play a vital role in the development of blood-related disorders including diabetes and cardiovascular diseases in humans. Various signaling mechanisms are involved in the activation of platelets. Initially, platelets are activated by the exposure of collagen and vWF from the extracellular matrix upon vascular damage. Activated platelets further release adenosine diphosphate (ADP) and generate thromboxane A2 (TxA2) that enables the recruitment of circulating platelets, activates the recruited platelets, and forms the hemostatic plug in the presence of thrombin by converting fibrinogen to fibrin. Collagen and vWF induce platelet activation by the glycoprotein (GP) VI-mediated signaling pathway through phospholipase C (PLC) γ2 activation [15]. In contrast, G protein-coupled receptor (GPCR) agonists ADP, TxA2, and thrombin cause platelet aggregation by the activation of the PLCβ pathway. ADP stimulates Gq-coupled P2Y1 and Gi-coupled P2Y12 receptor-mediated signaling pathways [16]. Likewise, thrombin activates platelets by coupling to Gq and G12/13 through protease-activated receptors (PARs) [17]. TxA2 activates platelets through the thromboxane prostanoid (TP) receptor by coupling to Gq and G12/13 [18,19]. In addition, ADP-induced TxA2 generation is regulated by calcium binding to cytosolic phospholipase A2 (cPLA2) [20,21]. Thrombin-induced TxA2 generation is mediated by the P2Y12 receptor [22]. The generated TxA2 then causes w positive-feedback effect on platelet functional response [23].
Several studies have suggested the occurrence of hyper-coagulation in patients with Cushing’s syndrome whose blood cortisol levels are high [12,24,25]. The study of Manetti et al. has shown that venous thromboembolism occurred in 7.5% of Cushing’s syndrome patients, whereas it did not occur in patients with remission of the disease [26]. Additionally, glucocorticoid administration is regarded as a first-line treatment option in several bleeding disorders, including immune thrombocytopenic purpura and acquired hemophilia A [27]. In contrast, in vitro studies using platelet-rich plasma (PRP) have shown that high glucocorticoid levels exert an inhibitory effect on platelet function [28,29,30]. Until now, the mechanisms involved in the effects of glucocorticoids on the production of thromboxane in platelets have not been elucidated.
In this study, we demonstrated the effect of prednisolone on platelet function and its underlying mechanism using washed murine platelets. We have shown that prednisolone inhibits 2-MeSADP-induced platelet secretion and the resultant secondary wave of aggregation. In aspirinated platelets, prednisolone had no effect on the 2-MeSADP-induced platelet aggregation as well as secretion, suggesting that prednisolone exerts its antiplatelet effect by regulating TxA2 generation. We have further shown that 2-MeSADP- and thrombin-induced TxA2 generation and 2-MeSADP-induced cPLA2 phosphorylation are inhibited in the presence of prednisolone. In conclusion, prednisolone affects platelet function by inhibiting TxA2 generation through the regulation of cPLA2 phosphorylation.
2. Materials and Methods
2.1. Materials
2-MeSADP, thrombin, apyrase, prostaglandin E1 (PGE1), sodium citrate, prednisolone, and acetylsalicylic acid (ASA) were purchased from Sigma (St. Louis, MO, USA). Anti-phospho-cPLA2 (Ser505), anti-cPLA2, anti-phospho-ERK (Thr202/Tyr204), and anti-total-ERK antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA). HRP-linked secondary antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Thromboxane B2 (TxB2) ELISA kit was purchased from Enzo Life Sciences (Exeter, UK). All other reagents were of reagent grade.
2.2. Animals
Thirty C57BL/6 mice were purchased from Orient Bio, Inc. at the age of eight weeks (Seongnam-si, Gyeonggi-Do, Republic of Korea) and housed in a semi-pathogen-free facility. No more than 5 mice were housed per cage and mice were freely provided with food and water under constant conditions of temperature (23 °C) and humidity (60%). All animal procedures were approved by the Chungbuk National University Animal Ethics Committee (CBNUA-873-15-02).
2.3. Platelet Preparation
Blood was collected via cardiac puncture from healthy mice in 3.8% sodium citrate as described previously [31]. PRP was obtained by centrifugation at 100× g for 10 min at room temperature (RT) and 1 µM PGE1 was added. For aspirin treatment, the PRP was treated with 1 mM ASA for 30 min at 37 °C. By centrifuging plasma at 400× g for 10 min, platelet pellets were obtained from the plasma and re-suspended in Tyrode’s buffer (pH 7.4) with 0.05 units/mL of apyrase. The platelet count was adjusted to 1 × 108 cells/mL.
2.4. Platelet Aggregation and Secretion Assay
The light transmission through a 250 μL sample of washed platelets (1 × 108 cells/mL) at 37 °C in a Lumi-aggregometer (Chrono-Log, Havertown, PA, USA) was measured to assess agonist-induced platelet aggregation while stirring at 900 rpm. Amounts of 250 nM, 500 nM, and 1000 nM prednisolone were pre-incubated for 5 min with washed platelets before stimulation with agonists. ATP release from platelets was assessed using a luciferin luciferase reagent to detect platelet dense granule secretion.
2.5. Measurement of TxA2 Generation
Washed platelets (1 × 108 cells/mL) were stimulated for 3.5 min in a platelet aggregometer under 900 rpm stirring conditions at 37 °C and the reaction was halted by snap-freezing the sample in liquid nitrogen. Samples were stored at −80 °C until TxB2 analysis was performed. Briefly, samples were centrifuged at 3000× g for 10 min at 4 °C after being thawed at RT, and the supernatant was diluted (1:20) with buffer. The diluted samples were measured to evaluate the levels of TxB2 in duplicates using a TxB2 ELISA Kit (Enzo Life Sciences, Exeter, UK), according to the manufacturer’s instructions.
2.6. Western Blot Analysis
Washed platelets were activated with 2-MeSADP for 2 min at 37 °C under stirring conditions, and the reaction was stopped by adding 6.6 N perchloric acid. Platelet samples were washed with distilled water, re-suspended with sample buffer, and denatured for 10 min. Platelet samples were separated on a 10% SDS polyacrylamide gel and transferred to polyvinylidene difluoride membranes. Nonspecific binding sites were blocked by SuperBlock® blocking buffer (Thermo Fisher Scientific, Waltham, MA, USA) at RT for 1 h, and membranes were incubated overnight with anti-phospho-cPLA2 (Ser505), anti-cPLA2, anti-phospho-ERK (Thr202/Tyr204), or anti-total-ERK antibodies diluted to a concentration of 1:1000 with gentle agitation. After 3 washes for 5 min each with Tris-buffered saline with 0.1% Tween 20, the membranes were incubated with appropriate secondary antibody at RT for 1 h. After washing, membranes were incubated with chemiluminescence substrate (Pierce, Rockford, IL, USA) for 5 min at RT, and immune reactivity was detected by using iBrightTM CL1500 (Thermo Fisher Scientific, Waltham, MA, USA).
2.7. Statistical Analysis
All statistical calculations were performed using Prism software (version 9.1). The data were presented as mean ± standard error (SE). Statistical significance was established using the one-way analysis of variance (ANOVA) followed by a post hoc Dunnett’s multiple comparison test. The normality test was conducted by using a Shapiro–Wilk test and data were found to be normally distributed (p > 0.05).
3. Results
3.1. Prednisolone Regulates 2-MeSADP-Induced Secondary Wave of Platelet Aggregation and Secretion in Platelets
In order to determine the role of prednisolone in platelet function, we first stimulated the platelets with 2-MeSADP and measured the platelet aggregation and dense granule secretion. As shown in Figure 1A, platelet aggregation and dense granule secretion induced by 2-MeSADP were inhibited in the presence of prednisolone in a concentration-dependent manner. 2-MeSADP-induced dense granule secretion and the secondary wave of aggregation were not completely inhibited by 250 nM prednisolone, while both dense granule secretion and the secondary wave of aggregation were completely inhibited in the presence of 500 nM prednisolone. Therefore, we used 500 nM prednisolone throughout the experiments. Interestingly, prednisolone treatment did not affect 2-MeSADP-induced primary aggregation. Thus, our findings suggest that prednisolone inhibits 2-MeSADP-induced dense granule secretion and the resultant secondary wave of aggregation.
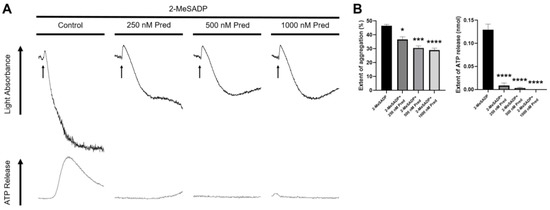
Figure 1.
The effect of prednisolone on 2-MeSADP-induced platelet aggregation and secretion. (A) Washed murine platelets were stimulated with 100 nM 2-MeSADP in the presence of different concentrations of prednisolone for 3.5 min under stirring conditions. Platelet aggregation and dense granule secretion were analyzed by a Lumi-aggregometer. The entire set of data represents three separate experiments (n = 3). (B) The extent of aggregation and dense granule secretion were quantified from panel A. Data are shown as mean ± SE. *, p < 0.05; ***, p < 0.001; ****, p < 0.0001.
3.2. The Effect of Prednisolone in 2-MeSADP-Induced Platelet Function in Aspirinated Platelets
Consistent with the previous result, prednisolone inhibited 2-MeSADP-induced platelet secretion and the resultant secondary wave of aggregation at both low and high concentrations of agonist (Figure 2A). It is well-established that ADP-induced platelet secretion and the secondary wave of aggregation require TxA2 generation [32,33]. In order to evaluate the effect of prednisolone on TxA2 generation, we stimulated the platelets with 2-MeSADP in the presence of aspirin, which blocks the positive-feedback effect of the generated TxA2. As shown in Figure 2B, prednisolone showed no further inhibition at both low and high concentrations of 2-MeSADP-induced platelet aggregation in aspirinated platelets compared to non-aspirinated platelets. These results suggest that prednisolone may inhibit 2-MeSADP-induced platelet aggregation and dense granule secretion by inhibiting TxA2 generation.
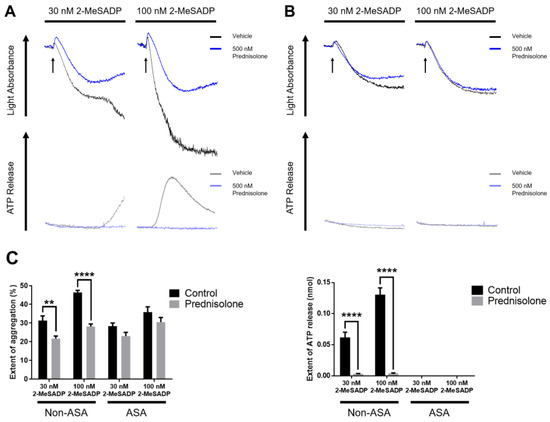
Figure 2.
The effect of prednisolone on 2-MeSADP-induced platelet aggregation and secretion in aspirinated platelets. (A) Non-aspirinated and (B) aspirinated washed platelets were stimulated with 30 nM and 100 nM 2-MeSADP in the presence or absence of 500 nM prednisolone and platelet aggregation and secretion were measured. The entire set of data represents three separate experiments (n = 6). (C) The extent of aggregation and dense granule secretion were quantified from panels A and B. Data are shown as mean ± SE. **, p < 0.01; ****, p < 0.0001.
3.3. Prednisolone Regulates Only Low Concentrations of Thrombin-Induced Platelet Aggregation and Secretion
To check the role of prednisolone in PAR-mediated platelet function, we stimulated the platelets with various concentrations of thrombin. As shown in Figure 3A, prednisolone inhibited a low concentration of thrombin-induced platelet aggregation and dense granule secretion. However, prednisolone could only partially inhibit platelet secretion when stimulated with a high concentration of thrombin. Additionally, prednisolone did not show any further inhibitory effect on thrombin-induced platelet aggregation and dense granule secretion in aspirinated platelets (Figure 3B). It has been known that platelet aggregation and dense granule secretion only require a positive-feedback effect of generated TxA2 when stimulated with a low concentration of thrombin [34,35]. Taken together, our data suggest that prednisolone affects low concentration of thrombin-induced platelet aggregation and dense granule secretion by regulating the positive-feedback effect of TxA2 generation.
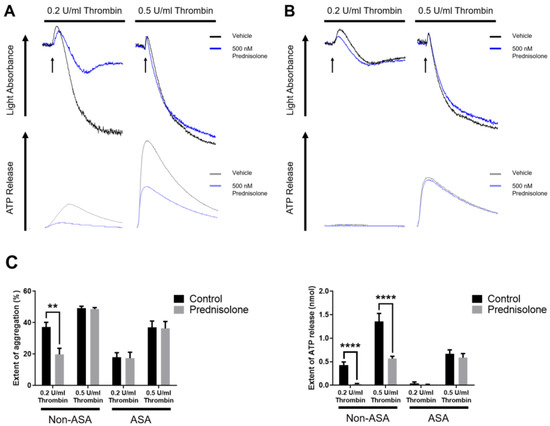
Figure 3.
The effect of prednisolone on thrombin-induced platelet aggregation and secretion. (A) Non-aspirinated and (B) aspirinated washed platelets were stimulated with 0.2 U/mL and 0.5 U/mL thrombin in the presence or absence of 500 nM prednisolone, and platelet aggregation and secretion were measured. The entire set of data represents three separate experiments (n = 6). (C) The extent of aggregation and dense granule secretion were quantified from panels A and B. Data are presented as mean ± SE. **, p < 0.01; ****, p < 0.0001.
3.4. Prednisolone Inhibits 2-MeSADP- and Thrombin-Induced TxA2 Generation in Platelets
To confirm the effect of prednisolone on TxA2 generation, we stimulated the platelets with 2-MeSADP and thrombin and measured the amount of TxA2 release. As shown in Figure 4, both low and high concentrations of 2-MeSADP- and thrombin-induced TxA2 generation were significantly inhibited in the presence of prednisolone, confirming the inhibitory effect of prednisolone on TxA2 generation.
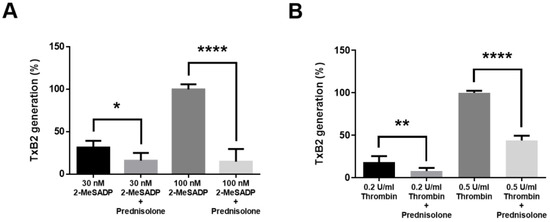
Figure 4.
The effect of prednisolone on TxA2 generation induced by 2-MeSADP and thrombin in platelets. Washed platelets were stimulated with different concentrations of (A) 2-MeSADP and (B) thrombin for 3.5 min in the presence or absence of 500 nM prednisolone and TxB2 generation was measured (n = 6). Data were expressed as percentages compared to 100 nM 2MeSADP and 0.5 U/mL thrombin groups and are shown as mean ± SE. *, p < 0.05; **, p < 0.01; ****, p < 0.0001.
3.5. Prednisolone Inhibits 2-MeSADP-Induced cPLA2 and ERK Phosphorylation
ERK phosphorylation is one of the most important upstream signaling molecules in TxA2 generation in platelets [36]. To understand the molecular mechanism involved in the regulation of TxA2 generation by prednisolone, we stimulated the platelets with 2-MeSADP and measured the cPLA2 ERK phosphorylation in non-aspirinated and aspirinated platelets. As shown in Figure 5, prednisolone completely inhibited 2-MeSADP-induced cPLA2 phosphorylation in both non-aspirinated and aspirinated platelets. However, 2-MeSADP-induced ERK phosphorylation was significantly inhibited by prednisolone only in non-aspirinated platelets, while prednisolone had no effect on 2-MeSADP-induced ERK phosphorylation in aspirinated platelets where there is no contribution of generated TxA2. These results confirm that prednisolone directly inhibits cPLA2 phosphorylation to regulate TxA2 generation in platelets.
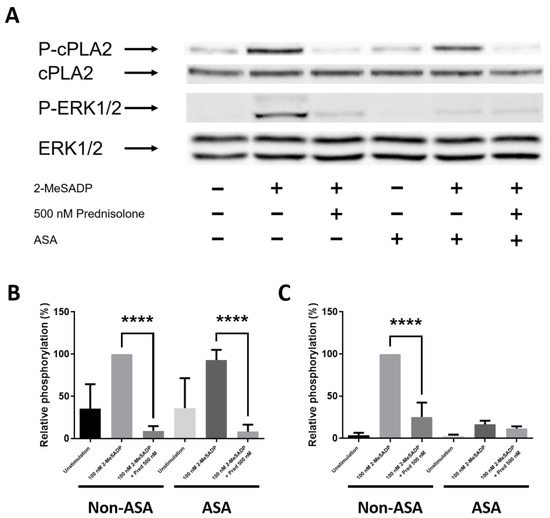
Figure 5.
The effect of prednisolone on 2-MeSADP-induced cPLA2 and ERK phosphorylation in platelets. (A) Non-aspirinated and aspirnated washed platelets were stimulated with 100 nM 2-MeSADP in the presence of 500 nM prednisolone. Proteins were immunoblotted using anti-phospho-cPLA2, anti-cPLA2, anti-phospho-ERK, and anti-ERK antibodies (n = 9). Relative phosphorylation of (B) cPLA2 and (C) ERK were expressed as a percentage compared to the 100 nM 2-MeSADP group in non-aspirinated platelets and are shown as mean ± SE. ****, p < 0.0001.
4. Discussion
Glucocorticoids are one of the most commonly used steroid medications to treat numerous types of allergies, inflammatory diseases, autoimmune disorders, and cancers [37]. Risks associated with glucocorticoid therapy are associated with dose, duration of therapy, and specific therapy used. Furthermore, chronic glucocorticoid administration can result in toxicities and undesirable effects such as Cushing’s syndrome even at physiological doses [38]. Thus, it is important to comprehend the precise mechanisms of the pathophysiological and therapeutic effects of glucocorticoids. One of the various side effects of glucocorticoid administration is hypercoagulopathy, which is known to be triggered by thrombocytosis, hyperglycemia, hypertension, and dyslipidemia [8,9,39]. However, there are discrepancies on how glucocorticoids affect platelet function, which is known to play a crucial role in coagulopathy. In contrast to the known role of glucocorticoids in hypercoagulability, previous studies have reported the inhibitory effect of glucocorticoids on platelet function using PRP [28,29,30,40]. PRP contains various coagulative factors that influence platelet functional responses. To rule out the contribution of the contents in PRP and thoroughly understand the molecular mechanism involved in the regulation of platelet function by glucocorticoids, we used a washed platelet system and investigated the effect of prednisolone, one of the most commonly used glucocorticoids in platelets.
Sex hormones, another type of steroid, have a well-established role in platelet function [41,42,43]. As corticosteroids and sex hormones are generated from cholesterol via the same steroidogenic pathway, they share similar effects on target cells [44,45]. 17β-estradiol and progestin, sex hormones, have been known to inhibit ADP-induced platelet activation [41]. Consistently, we observed that 2-MeSADP-induced platelet dense granule secretion and the resultant secondary wave of aggregation were completely inhibited in the presence of prednisolone. It has been shown that ADP-induced secretion and the resultant secondary wave of aggregation are dependent on the positive-feedback effect of generated TxA2 [46]. When the contribution of TxA2 generation was blocked by aspirin treatment, prednisolone had no further inhibitory effect on 2-MeSADP-induced platelet aggregation and secretion compared to non-aspirinated platelets, suggesting that prednisolone inhibits the 2-MeSADP-induced secondary wave of aggregation and secretion through the inhibition of the TxA2 generation in platelets.
Thrombin is the most potent agonist that activates platelets via the PAR-mediated pathway [47]. It has been shown that 17β-estradiol and progestin inhibit thrombin-induced platelet activation as well [41]. In contrast, others have reported that 17β-estradiol and progesterone potentiate thrombin-induced platelet activation [42,43]. We found that thrombin-induced platelet aggregation and dense granule secretion were inhibited in the presence of prednisolone. However, unlike the effect of prednisolone on 2-MeSADP-induced platelet function, we observed that prednisolone only inhibited low-thrombin concentration-induced platelet aggregation and secretion, and only partially inhibited high-thrombin concentration-induced secretion. In contrast to ADP, it has been known that TxA2 generation and the subsequent granule secretion play a role at low-thrombin concentration-induced platelet activation [48]. Therefore, our data suggest that prednisolone affects thrombin-induced platelet function by explicitly inhibiting TxA2 generation. More importantly, we found that 2-MeSADP-and thrombin-induced TxA2 generation was significantly inhibited by prednisolone, confirming the effect of prednisolone on TxA2 generation.
Glucocorticoids are known to inhibit cPLA2 in several cells [49,50]. cPLA2 is an upstream molecule of TxA2 generation that separates arachinonic acid, a precursor of TxA2, from phospholipids [51]. Glucocorticoids have also been shown to activate ERK1/2 in PC12 cells (a pheochromocytoma cell) and vascular smooth muscle cells [52,53,54]. However, in mast cells, dexamethasone has suppressed the phosphorylation of proteins that are associated with the activation of the mitogen-activated protein kinases (MAPKs) [55]. So far, the effect of glucocorticoids on cPLA2 and MAPKs has not been determined in platelets. In our study, we found that 2-MeSADP-induced platelet cPLA2 phosphorylation was completely inhibited by prednisolone in both non-aspirinated and aspirinated platelets. However, 2-MeSADP-induced ERK phosphorylation was significantly inhibited only in non-aspirinated platelets, whereas no additional inhibitory effect of prednisolone on ERK phosphorylation was observed in aspirinated platelets. It has been known that ERK plays a major role in TxA2 generation downstream of P2Y1, P2Y12, and PARs in platelets [22,33,56]. Further, it has been known that TxA2 can induce ERK phosphorylation by itself in platelets [57]. Thus, our data indicate that prednisolone affects platelet activation by inhibiting TxA2 generation by directly regulating cPLA2 phosphorylation, and that the effect of prednisolone on ERK phosphorylation is mediated by inhibition of TxA2 generation in platelets. High glucocorticoids have been shown to be effective in combination with first-line medication for some bleeding disorders because of their coagulation function [27]. It has also been demonstrated that in a majority of bleeding disorders, it is difficult to control the use of glucocorticoids as it may also lead to hypercoagulability such as in venous thromboembolism [58]. Furthermore, using glucocorticoids has been linked to the development of a number of metabolic disorders [59]. In contrast, glucocorticoids have also been known to cause bleeding disorders in the uterus and gastrointestinal tract [58,60]. Our study has demonstrated that prednisolone inhibits platelet activity, pointing to its potential role in hypo-coagulation and contradicting the prevalent notion that glucocorticoids promote coagulation.
5. Conclusions
Cushing’s syndrome is a disease that severely affects both humans and dogs and is potentially life-threatening if it remains untreated [61]. It significantly reduces the quality of life and, thus, requires effective treatment [62]. In this study, we demonstrate that prednisolone, a glucocorticoid, inhibits platelet function by the inhibition of TxA2 generation through the regulation of cPLA2 phosphorylation. Moreover, it indirectly regulates generated TxA2-mediated ERK phosphorylation to control platelet functional responses. Overall, our study provides evidence that could be applied to the therapeutic use of prednisolone in coagulation and bleeding disorders as well as the clinical management of Cushing’s syndrome in dogs in the future.
Author Contributions
Conceptualization, S.K. (Sanggu Kim) and S.K. (Soochong Kim); methodology, S.K. (Sanggu Kim) and P.K.C.; validation, S.K. (Soochong Kim); investigation, S.K. (Sanggu Kim) and P.K.C.; writing―original draft, S.K. (Sanggu Kim); writing―review and editing, P.K.C. and S.K. (Soochong Kim); supervision, S.K. (Soochong Kim). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Foundation of Korea (NRF-2022R1A2C1003638), the Basic Research Lab Program (2022R1A4A1025557) through the NRF of Korea funded by the Ministry of Science and ICT, the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Agriculture, Food and Rural Affairs Convergence Technologies Program for Educating Creative Global Leader, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA; 320005-4), and the National IT Industry Promotion Agency (NIPA) of Korea grant funded by the Korea government (MSIT)(S2005-22-1031).
Institutional Review Board Statement
The animal study protocol was approved by the Chungbuk National University Animal Ethics Committee (CBNUA-873-15-02).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barnes, P.J. Molecular mechanisms and cellular effects of glucocorticosteroids. Immunol. Allergy Clin. North Am. 2005, 25, 451–468. [Google Scholar] [CrossRef] [PubMed]
- Fardet, L.; Petersen, I.; Nazareth, I. Prevalence of long-term oral glucocorticoid prescriptions in the UK over the past 20 years. Rheumatology 2011, 50, 1982–1990. [Google Scholar] [CrossRef] [PubMed]
- Bénard-Laribière, A.; Pariente, A.; Pambrun, E.; Bégaud, B.; Fardet, L.; Noize, P. Prevalence and prescription patterns of oral glucocorticoids in adults: A retrospective cross-sectional and cohort analysis in France. BMJ Open 2017, 7, e015905. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Shang, N.; Zhang, Y.; Wang, D. Evaluation of Off-label Use of Oral Glucocorticoids in Outpatients. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2015, 37, 430–434. [Google Scholar] [PubMed]
- Chen, G.; Hu, W.; Xuan, N.; Fan, H.; Zhu, J.; Cui, W.; Zhang, G. Survey on the use of glucocorticoids in severe community-acquired pneumonia in intensive care unit of forty-five hospitals in Zhejiang Province. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2019, 31, 488–492. [Google Scholar]
- Gilor, C.; Graves, T.K. Interpretation of laboratory tests for canine Cushing’s syndrome. Top. Companion Anim. Med. 2011, 26, 98–108. [Google Scholar] [CrossRef]
- Graversen, D.; Vestergaard, P.; Stochholm, K.; Gravholt, C.; Jørgensen, J. Mortality in Cushing’s syndrome: A systematic review and meta-analysis. Eur. J. Intern. Med. 2012, 23, 278–282. [Google Scholar] [CrossRef]
- Wei, L.; MacDonald, T.M.; Walker, B.R. Taking glucocorticoids by prescription is associated with subsequent cardiovascular disease. Ann. Intern. Med. 2004, 141, 764–770. [Google Scholar] [CrossRef]
- Fardet, L.; Fève, B. Systemic glucocorticoid therapy: A review of its metabolic and cardiovascular adverse events. Drugs 2014, 74, 1731–1745. [Google Scholar] [CrossRef]
- Park, F.M.; Blois, S.L.; Abrams-Ogg, A.C.; Wood, R.D.; Allen, D.G.; Nykamp, S.G.; Downie, A. Hypercoagulability and ACTH-dependent hyperadrenocorticism in dogs. J. Vet. Intern. Med. 2013, 27, 1136–1142. [Google Scholar] [CrossRef]
- Casonato, A.; Pontara, E.; Boscaro, M.; Sonino, N.; Sartorello, F.; Ferasin, S.; Girolami, A. Abnormalities of von Willebrand factor are also part of the prothrombotic state of Cushing’s syndrome. Blood Coagul. Fibrinolysis 1999, 10, 145–151. [Google Scholar] [CrossRef]
- Erem, C.; Nuhoglu, I.; Yilmaz, M.; Kocak, M.; Demirel, A.; Ucuncu, O.; Ersoz, H.O. Blood coagulation and fibrinolysis in patients with Cushing’s syndrome: Increased plasminogen activator inhibitor-1, decreased tissue factor pathway inhibitor, and unchanged thrombin-activatable fibrinolysis inhibitor levels. J. Endocrinol. Investig. 2009, 32, 169–174. [Google Scholar] [CrossRef]
- Rose, L.; Dunn, M.E.; Bedard, C. Effect of canine hyperadrenocorticism on coagulation parameters. J. Vet. Intern. Med. 2013, 27, 207–211. [Google Scholar] [CrossRef]
- Jacoby, R.C.; Owings, J.T.; Ortega, T.; Gosselin, R.; Feldman, E.C. Biochemical basis for the hypercoagulable state seen in Cushing syndrome. Arch. Surg. 2001, 136, 1003–1007. [Google Scholar] [CrossRef]
- Wu, Y.-P.; Vink, T.; Schiphorst, M.; van Zanten, G.H.; IJsseldijk, M.J.; de Groot, P.G.; Sixma, J.J. Platelet thrombus formation on collagen at high shear rates is mediated by von Willebrand factor–glycoprotein Ib interaction and inhibited by von Willebrand factor–glycoprotein IIb/IIIa interaction. Arter. Thromb. Vasc. Biol. 2000, 20, 1661–1667. [Google Scholar] [CrossRef]
- Jin, J.; Kunapuli, S.P. Coactivation of two different G protein-coupled receptors is essential for ADP-induced platelet aggregation. Proc. Natl. Acad. Sci. USA 1998, 95, 8070–8074. [Google Scholar] [CrossRef]
- Offermanns, S. Activation of platelet function through G protein–coupled receptors. Circ. Res. 2006, 99, 1293–1304. [Google Scholar] [CrossRef]
- Knezevic, I.; Borg, C.; Le Breton, G.C. Identification of Gq as one of the G-proteins which copurify with human platelet thromboxane A2/prostaglandin H2 receptors. J. Biol. Chem. 1993, 268, 26011–26017. [Google Scholar] [CrossRef]
- Djellas, Y.; Manganello, J.M.; Antonakis, K.; Le Breton, G.C. Identification of Gα13 as one of the G-proteins that couple to human platelet thromboxane A2 receptors. J. Biol. Chem. 1999, 274, 14325–14330. [Google Scholar] [CrossRef]
- Channon, J.; Leslie, C.C. A calcium-dependent mechanism for associating a soluble arachidonoyl-hydrolyzing phospholipase A2 with membrane in the macrophage cell line RAW 264.7. J. Biol. Chem. 1990, 265, 5409–5413. [Google Scholar] [CrossRef]
- Kramer, R.M.; Roberts, E.F.; Um, S.L.; Börsch-Haubold, A.G.; Watson, S.P.; Fisher, M.J.; Jakubowski, J.A. p38 mitogen-activated protein kinase phosphorylates cytosolic phospholipase A2 (cPLA2) in thrombin-stimulated platelets: Evidence that proline-directed phosphorylation is not required for mobilization of arachidonic acid by cPLA2. J. Biol. Chem. 1996, 271, 27723–27729. [Google Scholar] [CrossRef] [PubMed]
- Shankar, H.; Garcia, A.; Prabhakar, J.; Kim, S.; Kunapuli, S. P2Y12 receptor-mediated potentiation of thrombin-induced thromboxane A2 generation in platelets occurs through regulation of Erk1/2 activation. J. Thromb. Haemost. 2006, 4, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.Z.; Jin, J.; Kunapuli, S.P. Molecular Mechanism of Thromboxane A2-induced Platelet Aggregation: ESSENTIAL ROLE FOR P2TAC and α2ARECEPTORS. J. Biol. Chem. 1999, 274, 29108–29114. [Google Scholar] [CrossRef] [PubMed]
- Fatti, L.; Bottasso, B.; Invitti, C.; Coppola, R.; Cavagnini, F.; Mannucci, P. Markers of activation of coagulation and fibrinolysis in patients with Cushing’s syndrome. J. Endocrinol. Investig. 2000, 23, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Patrassi, G.; Zanon, R.D.B.; Boscaro, M.; Martinelli, S.; Girolami, A. Further studies on the hypercoagulable state of patients with Cushing’s syndrome. Thromb. Haemost. 1985, 54, 518–520. [Google Scholar] [CrossRef]
- Manetti, L.; Bogazzi, F.; Giovannetti, C.; Raffaelli, V.; Genovesi, M.; Pellegrini, G.; Ruocco, L.; Iannelli, A.; Martino, E. Changes in coagulation indexes and occurrence of venous thromboembolism in patients with Cushing’s syndrome: Results from a prospective study before and after surgery. Eur. J. Endocrinol. 2010, 163, 783. [Google Scholar] [CrossRef]
- Curnow, J. Managing and supporting surgery in patients with bleeding disorders. Semin Thromb Hemost. 2017, 43, 653–671. [Google Scholar] [CrossRef]
- Casella, S.; Giudice, E.; Giannetto, C.; Marafioti, S.; Piccione, G. Effects of hydrocortisone and aminophylline on the aggregation of equine platelets in vitro. J. Vet. Sci. 2011, 12, 215–219. [Google Scholar] [CrossRef]
- Casella, S.; Giudice, E.; Alberghina, D.; Giannetto, C.; Marafioti, S.; Piccione, G. Hydrocortisone inhibition of adenosine diphosphate (ADP)-induced platelet aggregation in horse. Comp. Clin. Pathol. 2011, 20, 327–331. [Google Scholar] [CrossRef]
- Liverani, E.; Banerjee, S.; Roberts, W.; Naseem, K.M.; Perretti, M. Prednisolone exerts exquisite inhibitory properties on platelet functions. Biochem. Pharmacol. 2012, 83, 1364–1373. [Google Scholar] [CrossRef]
- Kim, S.; Cipolla, L.; Guidetti, G.; Okigaki, M.; Jin, J.; Torti, M.; Kunapuli, S.P. Distinct role of Pyk2 in mediating thromboxane generation downstream of both G12/13 and integrin αIIbβ3 in platelets. J. Biol. Chem. 2013, 288, 18194–18203. [Google Scholar] [CrossRef]
- Jin, J.; Quinton, T.M.; Zhang, J.; Rittenhouse, S.E.; Kunapuli, S.P. Adenosine diphosphate (ADP)–induced thromboxane A2generation in human platelets requires coordinated signaling through integrin αIIbβ3 and ADP receptors. Blood 2002, 99, 193–198. [Google Scholar] [CrossRef]
- Garcia, A.; Kim, S.; Bhavaraju, K.; Schoenwaelder, S.M.; Kunapuli, S.P. Role of phosphoinositide 3-kinase β in platelet aggregation and thromboxane A2 generation mediated by Gi signalling pathways. Biochem. J. 2010, 429, 369–377. [Google Scholar] [CrossRef]
- Chung, A.W.; Jurasz, P.; Hollenberg, M.D.; Radomski, M.W. Mechanisms of action of proteinase-activated receptor agonists on human platelets. Br. J. Pharmacol. 2002, 135, 1123. [Google Scholar] [CrossRef]
- Braun, M.; Kramann, J.; Strobach, H.; Schrör, K. Incomplete inhibition of platelet secretion by low-dose aspirin. Platelets 1994, 5, 325–331. [Google Scholar] [CrossRef]
- Yacoub, D.; Théorêt, J.-F.; Villeneuve, L.; Abou-Saleh, H.; Mourad, W.; Allen, B.G.; Merhi, Y. Essential role of protein kinase Cδ in platelet signaling, αIIbβ3 activation, and thromboxane A2 release. J. Biol. Chem. 2006, 281, 30024–30035. [Google Scholar] [CrossRef]
- Ronchetti, S.; Ayroldi, E.; Ricci, E.; Gentili, M.; Migliorati, G.; Riccardi, C. A glance at the use of glucocorticoids in rare inflammatory and autoimmune diseases: Still an indispensable pharmacological tool? Front. Immunol. 2021, 11, 613435. [Google Scholar] [CrossRef]
- Williams, D.M. Clinical pharmacology of corticosteroids. Respir. Care 2018, 63, 655–670. [Google Scholar] [CrossRef]
- Woolcock, A.; Keenan, A.; Cheung, C.; Christian, J.; Moore, G. Thrombocytosis in 715 Dogs (2011–2015). J. Vet. Intern. Med. 2017, 31, 1691–1699. [Google Scholar] [CrossRef]
- Filkova, A.A.; Martyanov, A.A.; Garzon Dasgupta, A.K.; Panteleev, M.A.; Sveshnikova, A.N. Quantitative dynamics of reversible platelet aggregation: Mathematical modelling and experiments. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Nakano, Y.; Oshima, T.; Matsuura, H.; Kajiyama, G.; Kambe, M. Effect of 17β-estradiol on inhibition of platelet aggregation in vitro is mediated by an increase in NO synthesis. Arter. Thromb. Vasc. Biol. 1998, 18, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Moro, L.; Reineri, S.; Piranda, D.; Pietrapiana, D.; Lova, P.; Bertoni, A.; Graziani, A.; Defilippi, P.; Canobbio, I.; Torti, M. Nongenomic effects of 17β-estradiol in human platelets: Potentiation of thrombin-induced aggregation through estrogen receptor β and Src kinase. Blood 2005, 105, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Blackmore, P.F. Progesterone metabolites rapidly stimulate calcium influx in human platelets by a src-dependent pathway. Steroids 2008, 73, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J. Sex hormones and glucocorticoids: Interactions with the immune system. Ann. N. Y. Acad. Sci. 1999, 876, 102–118. [Google Scholar] [CrossRef]
- Bereshchenko, O.; Bruscoli, S.; Riccardi, C. Glucocorticoids, sex hormones, and immunity. Front. Immunol. 2018, 9, 1332. [Google Scholar] [CrossRef]
- Zucker, M.B.; Peterson, J. Inhibition of adenosine diphosphate-induced secondary aggregation and other platelet functions by acetylsalicylic acid ingestion. Proc. Soc. Exp. Biol. Med. 1968, 127, 547–551. [Google Scholar] [CrossRef]
- Savage, B.; Cattaneo, M.; Ruggeri, Z.M. Mechanisms of platelet aggregation. Curr. Opin. Hematol. 2001, 8, 270–276. [Google Scholar] [CrossRef]
- Kim, S.; Foster, C.; Lecchi, A.; Quinton, T.M.; Prosser, D.M.; Jin, J.; Cattaneo, M.; Kunapuli, S.P. Protease-activated receptors 1 and 4 do not stimulate Gi signaling pathways in the absence of secreted ADP and cause human platelet aggregation independently of Gisignaling. Blood 2002, 99, 3629–3636. [Google Scholar] [CrossRef]
- Dolan–O’Keefe, M.; Nick, H.S. Inhibition of cytoplasmic phospholipase A2 expression by glucocorticoids in rat intestinal epithelial cells. Gastroenterology 1999, 116, 855–864. [Google Scholar] [CrossRef]
- Goppelt-Struebe, M.; Wolter, D.; Resch, K. Glucocorticoids inhibit prostaglandin synthesis not only at the level of phospholipase A2 but also at the level of cyclo-oxygenase/PGE isomerase. Br. J. Pharmacol. 1989, 98, 1287–1295. [Google Scholar] [CrossRef]
- Liu, N.-K.; Xu, X.-M. Phospholipase A 2 and its Molecular Mechanism after Spinal Cord Injury. Mol. Neurobiol. 2010, 41, 197–205. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, P.; Jing, Q.; Zhang, W.; Li, X.; Zhong, Y.; Sun, G.; Pei, G.; Chen, Y. Rapid activation of ERK1/2 mitogen-activated protein kinase by corticosterone in PC12 cells. Biochem. Biophys. Res. Commun. 2001, 287, 1017–1024. [Google Scholar] [CrossRef]
- Li, X.; Qiu, J.; Wang, J.; Zhong, Y.; Zhu, J.; Chen, Y. Corticosterone-induced rapid phosphorylation of p38 and JNK mitogen-activated protein kinases in PC12 cells. FEBS Lett. 2001, 492, 210–214. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, W.L.; Tasker, J.G.; Zhou, J.R.; Peng, Y.L.; Miao, C.Y.; Yang, Y.J.; Jiang, C.L. Dexamethasone induces rapid promotion of norepinephrine-mediated vascular smooth muscle cell contraction. Mol. Med. Rep. 2013, 7, 549–554. [Google Scholar] [CrossRef]
- Rider, L.G.; Hirasawa, N.; Santini, F.; Beaven, M.A. Activation of the mitogen-activated protein kinase cascade is suppressed by low concentrations of dexamethasone in mast cells. J. Immunol. 1996, 157, 2374–2380. [Google Scholar] [CrossRef]
- Garcia, A.; Shankar, H.; Murugappan, S.; Kim, S.; Kunapuli, S.P. Regulation and functional consequences of ADP receptor-mediated ERK2 activation in platelets. Biochem. J. 2007, 404, 299–308. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, P.; Zhu, L.; Li, N.; Hu, H. P2X1-initiated p38 signalling enhances thromboxane A2-induced platelet secretion and aggregation. Thromb. Haemost. 2014, 112, 142–150. [Google Scholar]
- Luo, X.; Feng, H.; Jiang, L.; Chen, Q. Abnormal uterine bleeding induced by glucocorticoid treatment for pemphigus: Case series and proposed treatment algorithm. Saudi Med. J. 2016, 37, 1025. [Google Scholar] [CrossRef]
- Bauerle, K.T.; Harris, C. Glucocorticoids and diabetes. Mo. Med. 2016, 113, 378. [Google Scholar]
- Narum, S.; Westergren, T.; Klemp, M. Corticosteroids and risk of gastrointestinal bleeding: A systematic review and meta-analysis. BMJ Open 2014, 4, e004587. [Google Scholar] [CrossRef]
- Nagata, N.; Kojima, K.; Yuki, M. Comparison of survival times for dogs with pituitary-dependent hyperadrenocorticism in a primary-care hospital: Treated with trilostane versus untreated. J. Vet. Intern. Med. 2017, 31, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Mc Bride, M.; Crespo, I.; Webb, S.M.; Valassi, E. Quality of life in Cushing’s syndrome. Best Pr. Res. Clin. Endocrinol. Metab. 2021, 35, 101505. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).