
6-Gingerol Improves In Vitro Porcine Embryo Development by Reducing Oxidative Stress
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Oocyte Collection and IVM
2.2. Parthenogenetic Activation and In Vitro Embryo Culture
2.3. Cell Proliferation Analysis
2.4. Terminal Deoxynucleotidyl Transferase-Mediated dUTP-Biotin Nick End-Labeling (TUNEL) Assays
2.5. ROS and Glutathione (GSH) Assays
2.6. Mitochondrial Membrane Potential (MMP, ΔΨm) Assay
2.7. Determination of ATP Levels
2.8. Immunofluorescence
2.9. RNA Extraction and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
3.1. 6-G Improved the Blastocyst Formation Rate
3.2. 6-G Enhanced Cell Proliferation
3.3. 6-G Reduced Apoptosis of Porcine Embryos
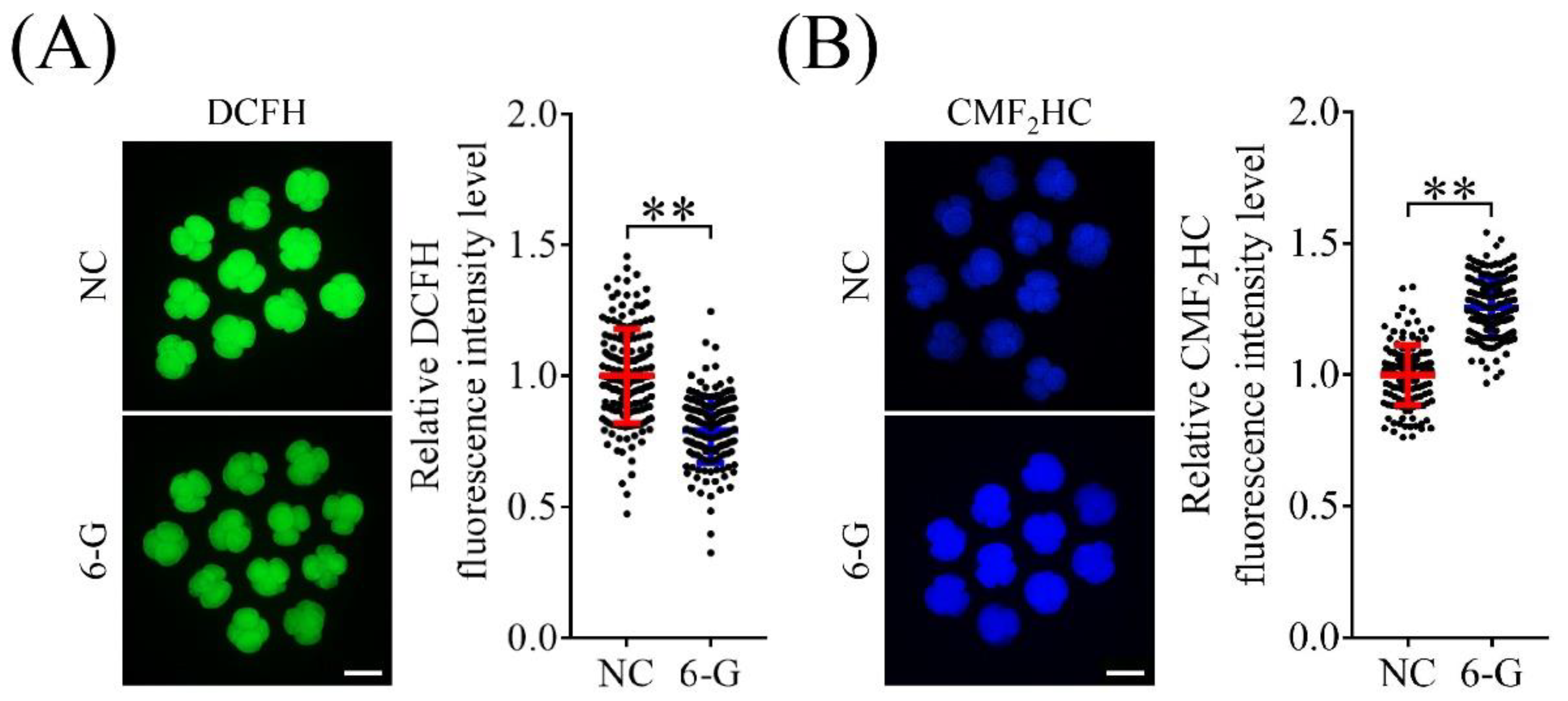
3.4. 6-G Enhanced Antioxidant Capacity in Porcine Embryos
3.5. 6-G Improved Mitochondrial Function
3.6. 6-G Inhibited Autophagy in Blastocysts
3.7. 6-G Regulated Embryo Pluripotency, Apoptosis, Autophagy, and Proliferation-Related Gene Expression in Blastocysts
3.8. 6-G Regulated MAPKs Activations in Blastocysts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fowler, K.E.; Mandawala, A.A.; Griffin, D.K.; Walling, G.A.; Harvey, S.C. The production of pig preimplantation embryos in vitro: Current progress and future prospects. Reprod. Biol. 2018, 18, 203–211. [Google Scholar] [CrossRef]
- Abeydeera, L.R. In vitro fertilization and embryo development in pigs. Reprod. (Camb. Engl.) Suppl. 2001, 58, 159–173. [Google Scholar] [CrossRef]
- Gil, M.A.; Cuello, C.; Parrilla, I.; Vazquez, J.M.; Roca, J.; Martinez, E.A. Advances in swine in vitro embryo production technologies. Reprod. Domest. Anim. 2010, 45 (Suppl. S2), 40–48. [Google Scholar] [CrossRef] [PubMed]
- Ashkar, F.A.; Semple, E.; Schmidt, C.H.; St John, E.; Bartlewski, P.M.; King, W.A. Thyroid hormone supplementation improves bovine embryo development in vitro. Hum. Reprod. 2010, 25, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukaya, T.; Yamanaka, T.; Terada, Y.; Murakami, T.; Yajima, A. Growth hormone improves mouse embryo development in vitro, and the effect is neutralized by growth hormone receptor antibody. Tohoku J. Exp. Med. 1998, 184, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Chen, Y.; Shu, Y.; Cheng, Y.; Qiao, J.; Behr, B.; Pera, R.A.; Hsueh, A.J. Promotion of human early embryonic development and blastocyst outgrowth in vitro using autocrine/paracrine growth factors. PLoS ONE 2012, 7, e49328. [Google Scholar] [CrossRef] [Green Version]
- Grupen, C.G. The evolution of porcine embryo in vitro production. Theriogenology 2014, 81, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Booth, P.J.; Holm, P.; Callesen, H. The effect of oxygen tension on porcine embryonic development is dependent on embryo type. Theriogenology 2005, 63, 2040–2052. [Google Scholar] [CrossRef] [PubMed]
- Im, G.S.; Lai, L.; Liu, Z.; Hao, Y.; Wax, D.; Bonk, A.; Prather, R.S. In vitro development of preimplantation porcine nuclear transfer embryos cultured in different media and gas atmospheres. Theriogenology 2004, 61, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Cheng, D.; Gao, X.; Bao, J.; Ma, X.; Wang, H. Vitamin C enhances the in vitro development of porcine pre-implantation embryos by reducing oxidative stress. Reprod. Domest. Anim. 2012, 47, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Nisenblat, V.; Lu, C.; Li, R.; Qiao, J.; Zhen, X.; Wang, S. Pretreatment with coenzyme Q10 improves ovarian response and embryo quality in low-prognosis young women with decreased ovarian reserve: A randomized controlled trial. Reprod. Biol. Endocrinol. 2018, 16, 29. [Google Scholar] [CrossRef] [Green Version]
- Singletary, K. Ginger: An overview of health benefits. Nutr. Today 2010, 45, 171–183. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Clinical aspects and health benefits of ginger (Zingiber officinale) in both traditional Chinese medicine and modern industry. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2019, 69, 546–556. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.; Rahmat, A.J.B.C.; Medicine, A. Optimization protocol for the extraction of 6-gingerol and 6-shogaol from Zingiber officinale var. rubrum Theilade and improving antioxidant and anticancer activity using response surface methodology. BMC Complement. Altern. Med. 2015, 15, 258. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, E.; Bava, S.V.; Narayanan, S.S.; Nath, L.R.; Thulasidasan, A.K.T.; Soniya, E.V.; Anto, R.J. [6]-Gingerol induces caspase-dependent apoptosis and prevents PMA-induced proliferation in colon cancer cells by inhibiting MAPK/AP-1 signaling. PLoS ONE 2014, 9, e104401. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, X.; Jiang, Y.; Wang, N.; Li, F.; Xin, H. 6-Gingerol Attenuates Ischemia-Reperfusion-Induced Cell Apoptosis in Human AC16 Cardiomyocytes through HMGB2-JNK1/2-NF-kappaB Pathway. Evid.-Based Complement. Altern. Med. 2019, 2019, 8798653. [Google Scholar] [CrossRef]
- Bing, Y.; Che, L.; Hirao, Y.; Takenouchi, N.; Rodríguez-Martínez, H.; Nagai, T. Parthenogenetic activation and subsequent development of porcine oocytes activated by a combined electric pulse and butyrolactone I treatment. J. Reprod. Dev. 2003, 49, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuma, T.; Kondo, T.; Ikeda, S.; Imai, H.; Yamada, M. Effects of EDTA saturated with Ca2+ (Ca-EDTA) on pig, bovine and mouse oocytes at the germinal vesicle stage during maturation culture and the involvement of chelation of Zn2+ in pronuclear formation induction by Ca-EDTA. Reproduction 2002, 124, 235–240. [Google Scholar] [CrossRef]
- Pournaderi, P.S.; Yaghmaei, P.; Khodaei, H.; Noormohammadi, Z.; Hejazi, S.H. The effects of 6-Gingerol on reproductive improvement, liver functioning and Cyclooxygenase-2 gene expression in estradiol valerate—Induced polycystic ovary syndrome in Wistar rats. Biochem. Biophys. Res. Commun. 2017, 484, 461–466. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Ojo, M.; Afolabi, T.T.; Arowoogun, M.D.; Nwawolor, D.; Farombi, E.O. Protective properties of 6-gingerol-rich fraction from Zingiber officinale (Ginger) on chlorpyrifos-induced oxidative damage and inflammation in the brain, ovary and uterus of rats. Chem. Biol. Interact. 2017, 270, 15–23. [Google Scholar] [CrossRef]
- Kitagawa, Y.; Suzuki, K.; Yoneda, A.; Watanabe, T. Effects of oxygen concentration and antioxidants on the in vitro developmental ability, production of reactive oxygen species (ROS), and DNA fragmentation in porcine embryos. Theriogenology 2004, 62, 1186–1197. [Google Scholar] [CrossRef]
- Lin, J.; Li, X.; Chen, L.; Lu, W.; Chen, X.; Han, L.; Chen, D.J. Protective Effect Against Hydroxyl Radical-induced DNA Damage and Antioxidant Mechanism of [6]-gingerol: A Chemical Study. Bull. Korean Chem. Soc. 2014, 35, 1633–1638. [Google Scholar] [CrossRef] [Green Version]
- Mohd Sahardi, N.F.N.; Makpol, S. Ginger (Zingiber officinale Roscoe) in the Prevention of Ageing and Degenerative Diseases: Review of Current Evidence. Evid.-Based Complement. Altern. Med. 2019, 2019, 5054395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Hyttel, P.; Laurincik, J.; Viuff, D.; Fair, T.; Zakhartchenko, V.; Rosenkranz, C.; Avery, B.; Rath, D.; Niemann, H.; Thomsen, P.D.; et al. Activation of ribosomal RNA genes in preimplantation cattle and swine embryos. Anim. Reprod. Sci. 2000, 60–61, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Bjerregaard, B.; Wrenzycki, C.; Strejcek, F.; Laurincik, J.; Holm, P.; Ochs, R.L.; Rosenkranz, C.; Callesen, H.; Rath, D.; Niemann, H.; et al. Expression of Nucleolar-Related Proteins in Porcine Preimplantation Embryos Produced In Vivo and In Vitro1. Biol. Reprod. 2004, 70, 867–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppertz, B.; Herrler, A. Regulation of proliferation and apoptosis during development of the preimplantation embryo and the placenta. Birth Defects Res. Part C 2005, 75, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishayee, K.; Chakraborty, D.; Ghosh, S.; Boujedaini, N.; Khuda-Bukhsh, A.R. Lycopodine triggers apoptosis by modulating 5-lipoxygenase, and depolarizing mitochondrial membrane potential in androgen sensitive and refractory prostate cancer cells without modulating p53 activity: Signaling cascade and drug-DNA interaction. Eur. J. Pharmacol. 2013, 698, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, M.K.; Hagen, T.M.; Ames, B.N. Oxidative damage and mitochondrial decay in aging. Proc. Natl. Acad. Sci. USA 1994, 91, 10771–10778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.Y.; Lee, K.C.; Chen, S.Y.; Chang, H.H. 6-Gingerol inhibits ROS and iNOS through the suppression of PKC-α and NF-κB pathways in lipopolysaccharide-stimulated mouse macrophages. Biochem. Biophys. Res. Commun. 2009, 382, 134–139. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Yao, L.; Ma, P.; Chen, Z.; Han, T.-L.; Yuan, C.; Zhang, J.; Jiang, L.; Liu, L.J.T.; et al. 6-gingerol ameliorates age-related hepatic steatosis: Association with regulating lipogenesis, fatty acid oxidation, oxidative stress and mitochondrial dysfunction. Toxicol. Appl. Pharmacol. 2019, 362, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-K.; Kim, Y.; Na, K.-M.; Surh, Y.-J.; Kim, T.-Y. [6]-Gingerol prevents UVB-induced ROS production and COX-2 expression in vitro and in vivo. Free Radic. Res. 2007, 41, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Park, G.H.; Kim, C.-Y.; Jang, J.-H. [6]-Gingerol attenuates β-amyloid-induced oxidative cell death via fortifying cellular antioxidant defense system. Food Chem. Toxicol. 2011, 49, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Kang, M.; Han, Y.; Zhang, T.; Quan, W.; Gao, J. 6-Gingerols (6G) reduces hypoxia-induced PC-12 cells apoptosis and autophagy through regulation of miR-103/BNIP3. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1653–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-L.; Zhuang, X.-D.; Xu, Z.-W.; Lu, L.-H.; Guo, H.-L.; Wu, W.-K.; Liao, X.-X. Higenamine combined with [6]-gingerol suppresses doxorubicin-triggered oxidative stress and apoptosis in cardiomyocytes via upregulation of PI3K/Akt pathway. Evid.-Based Complement. Altern. Med. 2013, 2013, 970490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unal, E.B.; Uhlitz, F.; Bluthgen, N. A compendium of ERK targets. FEBS Lett. 2017, 591, 2607–2615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xia, Y.; Liu, T.; Wang, J.; Dai, W.; Wang, F.; Zheng, Y.; Chen, K.; Li, S.; Abudumijiti, H.; et al. Protective effects of astaxanthin on ConA-induced autoimmune hepatitis by the JNK/p-JNK pathway-mediated inhibition of autophagy and apoptosis. PLoS ONE 2015, 10, e0120440. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, J.-G.; Qu, J.; Zhang, Q.; Prasad, C.; Wei, Z.-J. Assessment of anti-cancerous potential of 6-gingerol (Tongling White Ginger) and its synergy with drugs on human cervical adenocarcinoma cells. Food Chem. Toxicol. 2017, 109, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.-F.; Zong, S.-H.; Zhang, Z.-Y.; Fu, S.-W.; Li, K.-K.; Fang, Y.; Lu, L.; Xiao, D.-Q. The role of 6-gingerol on inhibiting amyloid β protein-induced apoptosis in PC12 cells. Rejuvenation Res. 2015, 18, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Khazaei, M.; Aghaz, F. Reactive Oxygen Species Generation and Use of Antioxidants during In Vitro Maturation of Oocytes. Int. J. Fertil. Steril. 2017, 11, 63–70. [Google Scholar] [CrossRef]
- Truong, T.; Gardner, D.K. Antioxidants improve IVF outcome and subsequent embryo development in the mouse. Hum. Reprod. 2017, 32, 2404–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.T.; Guo, J.; Niu, Y.J.; Cui, X.S. The toxic effect of aflatoxin B1 on early porcine embryonic development. Theriogenology 2018, 118, 157–163. [Google Scholar] [CrossRef]
- Murakami, M.; Ichisaka, T.; Maeda, M.; Oshiro, N.; Hara, K.; Edenhofer, F.; Kiyama, H.; Yonezawa, K.; Yamanaka, S. mTOR is essential for growth and proliferation in early mouse embryos and embryonic stem cells. Mol. Cell. Biol. 2004, 24, 6710–6718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanuma, K.; Tsuji, N.; Endoh, T.; Yagihashi, A.; Watanabe, N. Survivin enhances Fas ligand expression via up-regulation of specificity protein 1-mediated gene transcription in colon cancer cells. J. Immunol. 2004, 172, 3922–3929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Kim, E.Y.; Cui, X.S.; Tae, J.C.; Lee, W.D.; Kim, N.H.; Park, S.P.; Lim, J.H. Increase in DNA fragmentation and apoptosis-related gene expression in frozen-thawed bovine blastocysts. Zygote 2006, 14, 125–131. [Google Scholar] [CrossRef]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Negron-Perez, V.M.; Zhang, Y.; Hansen, P.J. Single-cell gene expression of the bovine blastocyst. Reproduction 2017, 154, 627–644. [Google Scholar] [CrossRef]
- Ortega, M.S.; Kelleher, A.M.; O’Neil, E.; Benne, J.; Cecil, R.; Spencer, T.E. NANOG is required to form the epiblast and maintain pluripotency in the bovine embryo. Mol. Reprod. Dev. 2019, 87, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, T.; Tada, M.; Kubota, H.; Kimura, H.; Hatano, S.Y.; Suemori, H.; Nakatsuji, N.; Tada, T. Octamer and Sox elements are required for transcriptional cis regulation of Nanog gene expression. Mol. Cell. Biol. 2005, 25, 2475–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of nanog by OCT4 and SOX2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnani, L.; Cabot, R.A. In vitro and in vivo derived porcine embryos possess similar, but not identical, patterns of Oct4, Nanog, and Sox2 mRNA expression during cleavage development. Mol. Reprod. Dev. 2008, 75, 1726–1735. [Google Scholar] [CrossRef]
- Shah, B.H.; Catt, K.J. Metabolism. Roles of LPA3 and COX-2 in implantation. Trends Endocrinol. Metab. 2005, 16, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Stanfield, K.; Bell, R.; Lisowski, A.; English, M.; Saldeen, S.; Khan, K.N.M. Expression of cyclooxygenase-2 in embryonic and fetal tissues during organogenesis and late pregnancy. Birth Defects Res. Part A 2003, 67, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Y.; Li, Y.; Yang, L.; Ma, Y.; Peng, X.; Yang, S.; Liu, J.; Li, H. Autophagy: A novel mechanism of chemoresistance in cancers. Biomed. Pharm. 2019, 119, 109415. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.D.; Qin, Z.H. Beclin 1, Bcl-2 and Autophagy. Adv. Exp. Med. Biol. 2019, 1206, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H. Regulation of Autophagy by mTOR Signaling Pathway. Adv. Exp. Med. Biol. 2019, 1206, 67–83. [Google Scholar] [CrossRef]
- Deleyto-Seldas, N.; Efeyan, A. The mTOR-Autophagy Axis and the Control of Metabolism. Front. Cell Dev. Biol. 2021, 9, 655731. [Google Scholar] [CrossRef]
- Gump, J.M.; Thorburn, A. Autophagy and apoptosis: What is the connection? Trends Cell. Biol. 2011, 21, 387–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, C.; Yang, G.; Yang, Y. Biological properties of 6-gingerol: A brief review. Nat. Prod. Commun. 2014, 9, 1027–1030. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.P.; Wang, X.Q.; Wang, J.; Dan, L.; Li, Y.H.; Jiang, H.; Xu, Y.N.; Kim, N.H. Oroxin A reduces oxidative stress, apoptosis, and autophagy and improves the developmental competence of porcine embryos in vitro. Reprod. Domest. Anim. 2022, 57, 1255–1266. [Google Scholar] [CrossRef]
- Yu, W.J.; Chen, C.Z.; Peng, Y.X.; Li, Z.; Gao, Y.; Liang, S.; Yuan, B.; Kim, N.H.; Jiang, H.; Zhang, J.B. Schisanhenol improves early porcine embryo development by regulating the phosphorylation level of MAPK. Theriogenology 2021, 175, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.R.; Ji, H.W.; He, S.Y.; Liu, R.P.; Wang, X.Q.; Wang, J.; Huang, C.M.; Xu, Y.N.; Li, Y.H.; Kim, N.H. Chrysoeriol Improves In Vitro Porcine Embryo Development by Reducing Oxidative Stress and Autophagy. Vet. Sci. 2023, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. C Embryo Today 2007, 81, 155–162. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Kim, J.; Lim, J.; Lee, E. Anthocyanin stimulates in vitro development of cloned pig embryos by increasing the intracellular glutathione level and inhibiting reactive oxygen species. Theriogenology 2010, 74, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.S.; Gupta, K.; Gupta, S. Green tea polyphenols causes cell cycle arrest and apoptosis in prostate cancer cells by suppressing class I histone deacetylases. Carcinogenesis 2012, 33, 377–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.-Y.; Seeram, N.P.; Nair, M.G.; Bourquin, L.D. Tart cherry anthocyanins inhibit tumor development in ApcMin mice and reduce proliferation of human colon cancer cells. Cancer Lett. 2003, 194, 13–19. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Modulation of signaling pathways in prostate cancer by green tea polyphenols. Biochem. Pharmacol. 2013, 85, 667–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, W.; Peng, Y.; Peng, X.; Li, Z.; Liu, C.; Yang, L.; Gao, Y.; Liang, S.; Yuan, B.; Chen, C.; et al. 6-Gingerol Improves In Vitro Porcine Embryo Development by Reducing Oxidative Stress. Animals 2023, 13, 1315. https://doi.org/10.3390/ani13081315
Yu W, Peng Y, Peng X, Li Z, Liu C, Yang L, Gao Y, Liang S, Yuan B, Chen C, et al. 6-Gingerol Improves In Vitro Porcine Embryo Development by Reducing Oxidative Stress. Animals. 2023; 13(8):1315. https://doi.org/10.3390/ani13081315
Chicago/Turabian StyleYu, Wenjie, Yanxia Peng, Xinyue Peng, Ze Li, Chang Liu, Liu Yang, Yan Gao, Shuang Liang, Bao Yuan, Chengzhen Chen, and et al. 2023. "6-Gingerol Improves In Vitro Porcine Embryo Development by Reducing Oxidative Stress" Animals 13, no. 8: 1315. https://doi.org/10.3390/ani13081315
APA StyleYu, W., Peng, Y., Peng, X., Li, Z., Liu, C., Yang, L., Gao, Y., Liang, S., Yuan, B., Chen, C., Kim, N.-h., Jiang, H., & Zhang, J. (2023). 6-Gingerol Improves In Vitro Porcine Embryo Development by Reducing Oxidative Stress. Animals, 13(8), 1315. https://doi.org/10.3390/ani13081315