
Chromosomes of Asian Cyprinid Fishes: Genomic Differences in Conserved Karyotypes of ‘Poropuntiinae’ (Teleostei, Cyprinidae)

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | 2n | NF | Karyotype | NORs/18S rDNA Pairs | Reference |
|---|---|---|---|---|---|
| Amblyrhynchichthys truncatus | 50 | 78 | 16m + 12sm + 22a | - | [25] |
| Barbonymus altus | 50 | 86 | 12m + 14sm + 10st + 14a | 2 | [26] |
| Barbonymus gonionotus | 50 | 66 | 2m + 4sm + 10st + 34a | - | [27] |
| Barbonymus gonionotus | 50 | 72 | 2m + 20sm + 4st + 24a | - | [28,29] |
| Barbonymus gonionotus | 50 | 74 | 12m + 12sm + 4st + 22a | - | [30] |
| Balantiocheilos melanopterus | 50 | 70 | 14m + 6sm + 10st + 20a | - | [31] |
| Balantiocheilos melanopterus | 50 | 74 | 6m + 18sm + 16st + 10a | 2 | [32] |
| Barbonymus schwanenfeldi | 50 | 76 | 6m + 6sm + 14st + 24a | - | [27] |
| Barbonymus schwanenfeldii | 50 | 84 | 6m + 28sm/st + 16a | - | [33] |
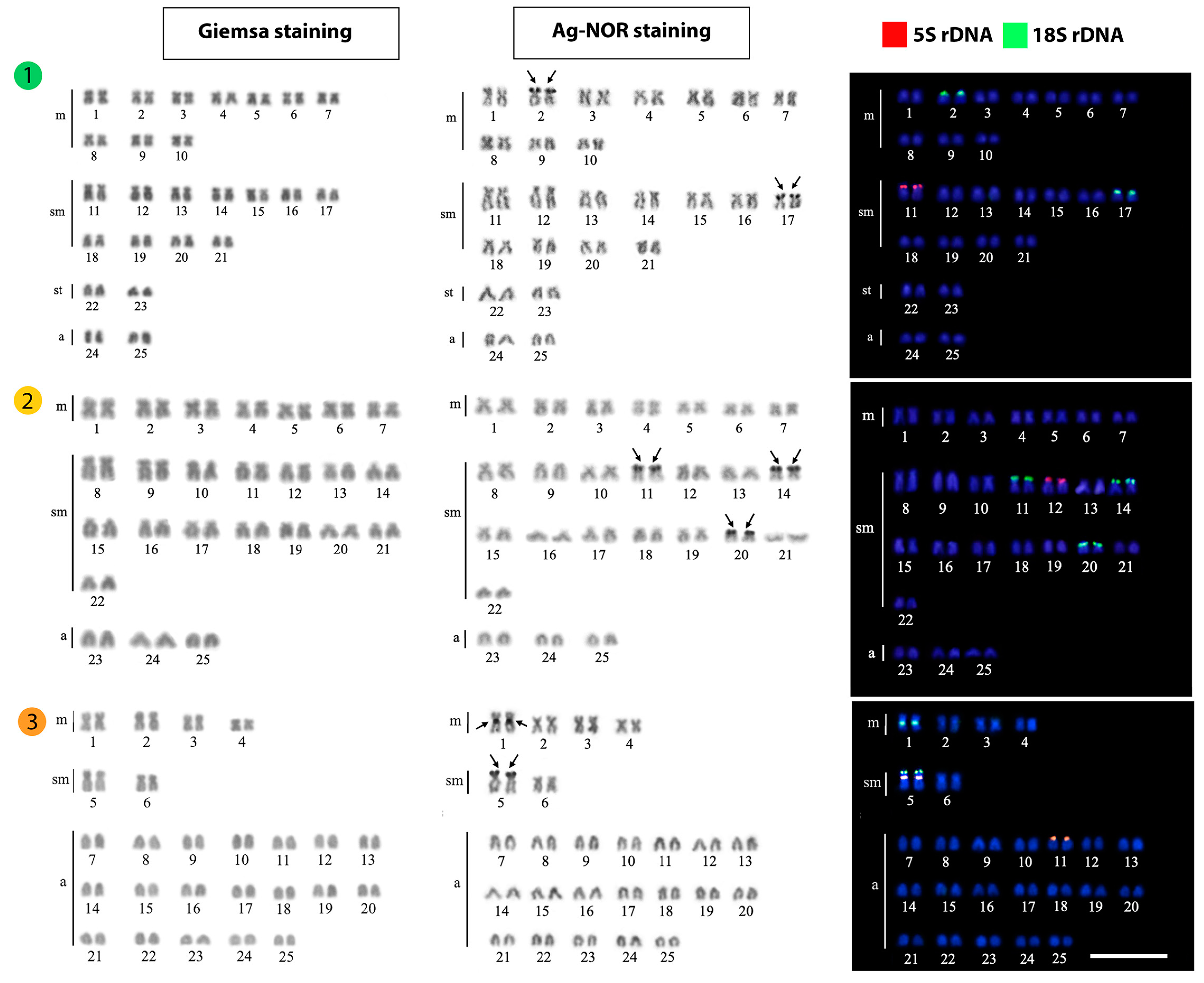
| Cosmochilus harmandi | 50 | 82 | 22m + 10sm + 10st + 8a | - | [34] |
| Cosmochilus harmandi | 50 | 84 | 12m + 16sm + 6st + 16a | 8 | [26] |
| Cosmochilus harmandi | 50 | 92 | 20m + 22sm + 4st + 4a | 2, 17 | Present work |
| Cyclocheilichthys apogon | 50 | 70 | 12m + 8sm + 6st + 24a | - | [28] |
| Cyclocheilichthys apogon | 50 | 76 | 18m + 8sm + 4st + 20a | - | [31] |
| Cyclocheilichthys apogon | 50 | 86 | 10m + 16sm + 10st + 14a | 6 | [26] |
| Cyclocheilichthys apogon | 50 | 74 | 14m + 30sm + 6a | 11, 14, 20 | Present work |
| Cyclocheilichthys armatus | 50 | 94 | 12m + 18sm + 14st + 6a | 3, 7 | [35] |
| Cyclocheilichthys repasson | 50 | 78 | 12m + 16sm + 6st + 16a | - | [36] |
| Cyclocheilichthys repasson | 50 | 84 | 6m + 6sm + 22st + 16a | - | [27] |
| Cyclocheilos enoplos | 50 | 90 | 10m + 30sm + 4st + 6a | two pairs (sm, a) | [37] |
| Cyclocheilos enoplos | 50 | 72 | 14m + 8sm + 10st + 18a | - | [38] |
| Cyclocheilos enoplos | 50 | 78 | 16m + 12sm + 6st + 16a | - | [31] |
| Hypsibarbus lagleri | 50 | 74 | 4m + 20sm + 26a | - | [39] |
| Hypsibarbus malcolmi | 50 | 64 | 10m + 4sm + 36a | - | [36] |
| Hypsibarbus malcolmi | 50 | 62 | 8m + 4sm + 38a | 1, 5 | Present work |
| Hypsibarbus vernayi | 50 | 58 | 6m + 2sm + 4st + 38a | - | [39] |
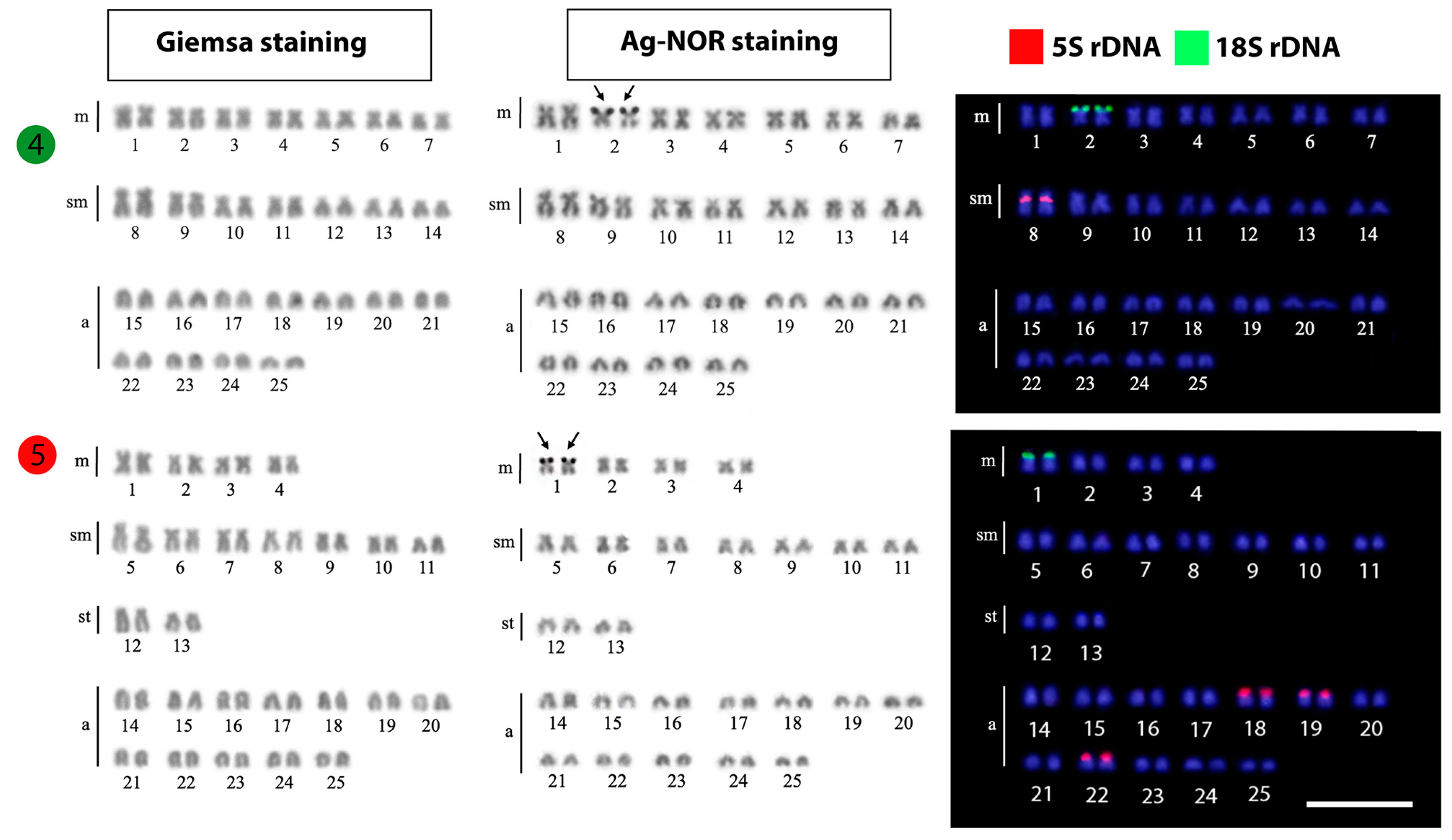
| Hypsibarbus wetmorei | 50 | 70 | 12m + 8sm + 6st + 24a | - | [28] |
| Hypsibarbus wetmorei | 50 | 74 | 12m + 12sm + 4st + 22a | 2 | [40] |
| Hypsibarbus wetmorei | 50 | 74 | 12m + 12sm + 2st + 24a | - | [39] |
| Hypsibarbus wetmorei | 50 | 82 | 10m + 14sm + 8st + 18a | 6 | [26] |
| Hypsibarbus wetmorei | 50 | 78 | 14m + 14sm + 22a | 2 | Present work |
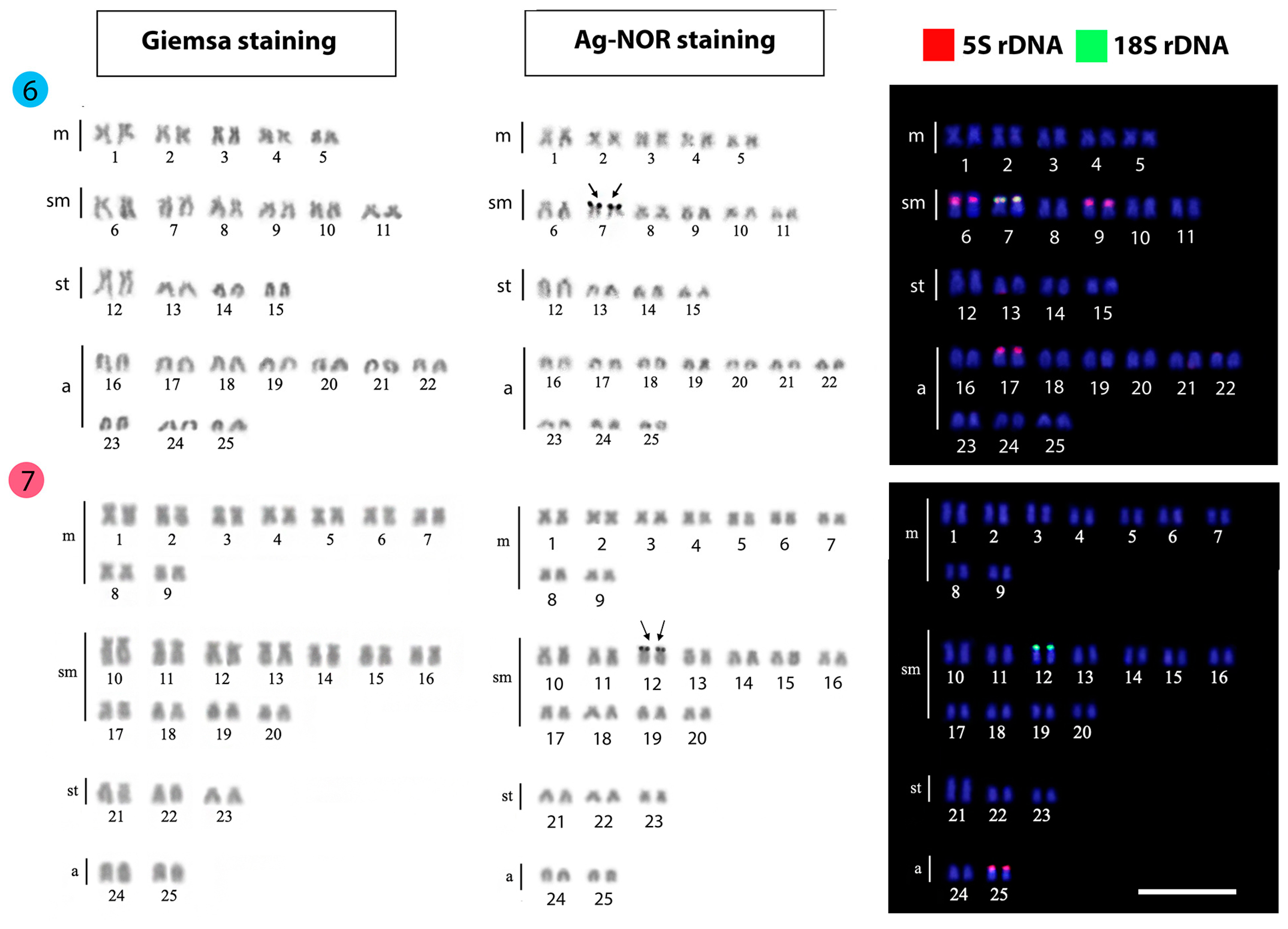
| Mystacoleucus argenteus | 50 | 76 | 6m + 20sm + 2st + 22a | - | [25] |
| Mystacoleucus chilopterus | 50 | 72 | 8m + 14sm + 4st + 24a | 1 | Present work |
| Mystacoleucus ectypus | 50 | 72 | 10m + 12sm + 8st + 20a | 7 | Present work |
| Mystacoleucus marginatus | 50 | 76 | 16m + 10sm + 24a | - | [41] |
| Mystacoleucus marginatus | 50 | 68 | 14m + 4sm + 2st + 30a | - | [31] |
| Poropuntius chonglingchungi | 50 | 80 | 12m + 18sm + 20a | - | [42] |
| Poropuntius deauratus | 50 | 74 | 14m + 10sm + 26a | - | [34] |
| Poropuntius laoensis | 50 | 74 | 14m + 10sm + 10st + 16a | - | [43] |
| Poropuntius normani | 50 | 72 | 10m + 12sm + 28a | - | [36] |
| Poropuntius sinensis | 50 | 82 | 10m + 22sm + 18a | [44] | |
| Puntioplties falcifer | 50 | 80 | 14m + 16sm + 2st + 18a | - | [36] |
| Puntioplties falcifer | 50 | 92 | 16m + 10sm + 16st + 8a | [27] | |
| Puntioplties proctozysron | 50 | 76 | 20m + 6sm + 6st + 18a | - | [38] |
| Puntioplties proctozysron | 50 | 76 | 16m + 10sm + 24a | - | [37] |
| Puntioplties proctozysron | 50 | 82 | 6m + 14sm + 12st + 18a | 2 | [45] |
| Puntioplties proctozysron | 50 | 90 | 18m + 22sm + 6st + 4a | 12 | Present work |
| Scaphognathops bandanensis | 50 | 64 | 10m + 6sm + 34a | - | [36] |
| Scaphognathops bandanensis | 50 | 66 | 10m + 6sm + 34a | 2 | [16] |
| Sikukia gudgeri | 50 | 68 | 10m + 8sm + 4st + 28a | - | [34] |
2. Materials and Methods
2.1. Individuals, Mitotic Chromosome Preparation and Ag-NOR Banding
2.2. Fluorescence In Situ Hybridization (FISH)
2.3. Comparative Genomic Hybridization (CGH)
2.4. Karyotyping and Image Processing
3. Results
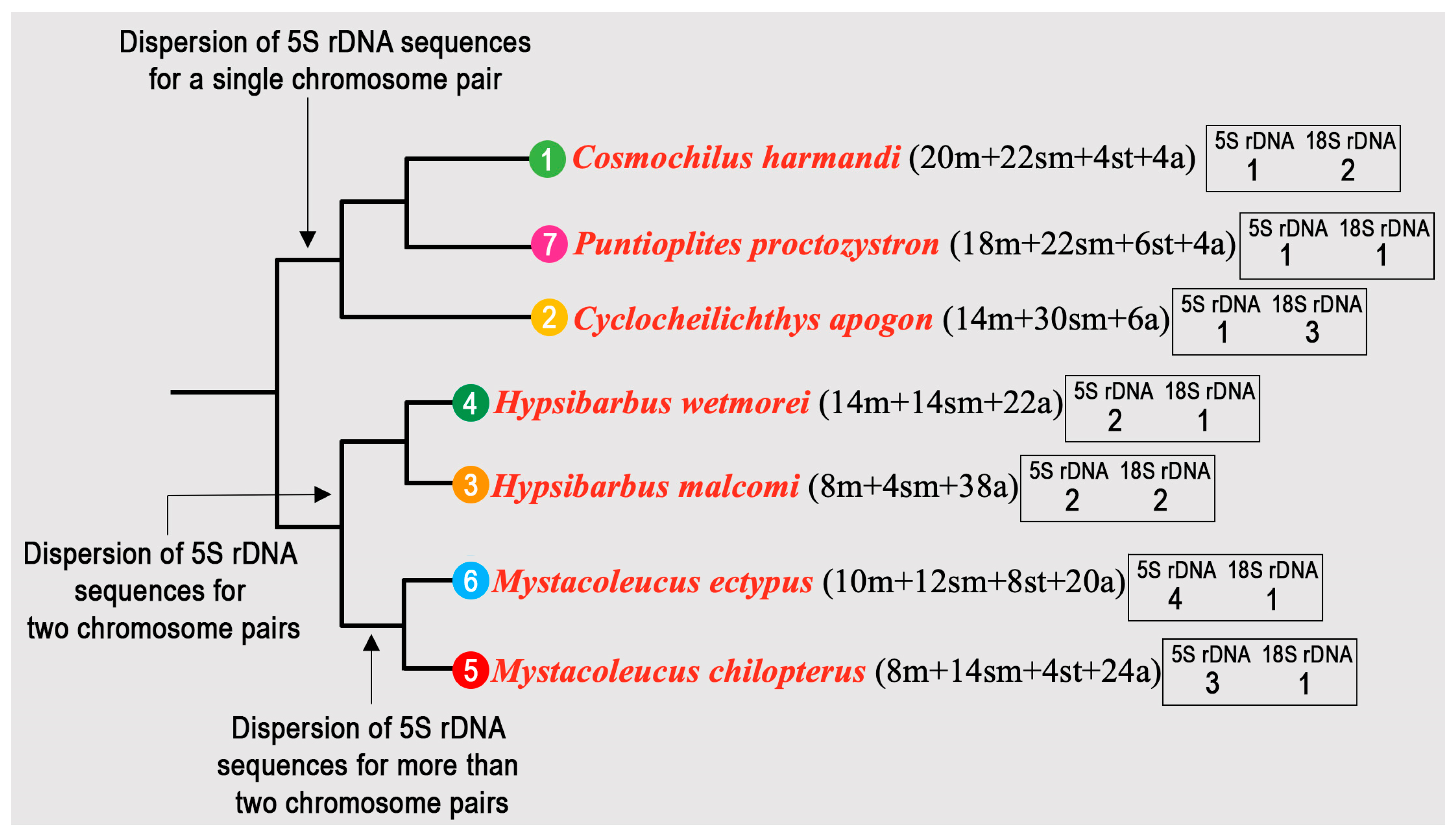
3.1. Karyotypes and Ag-NOR Phenotypes
3.2. FISH-Mapping
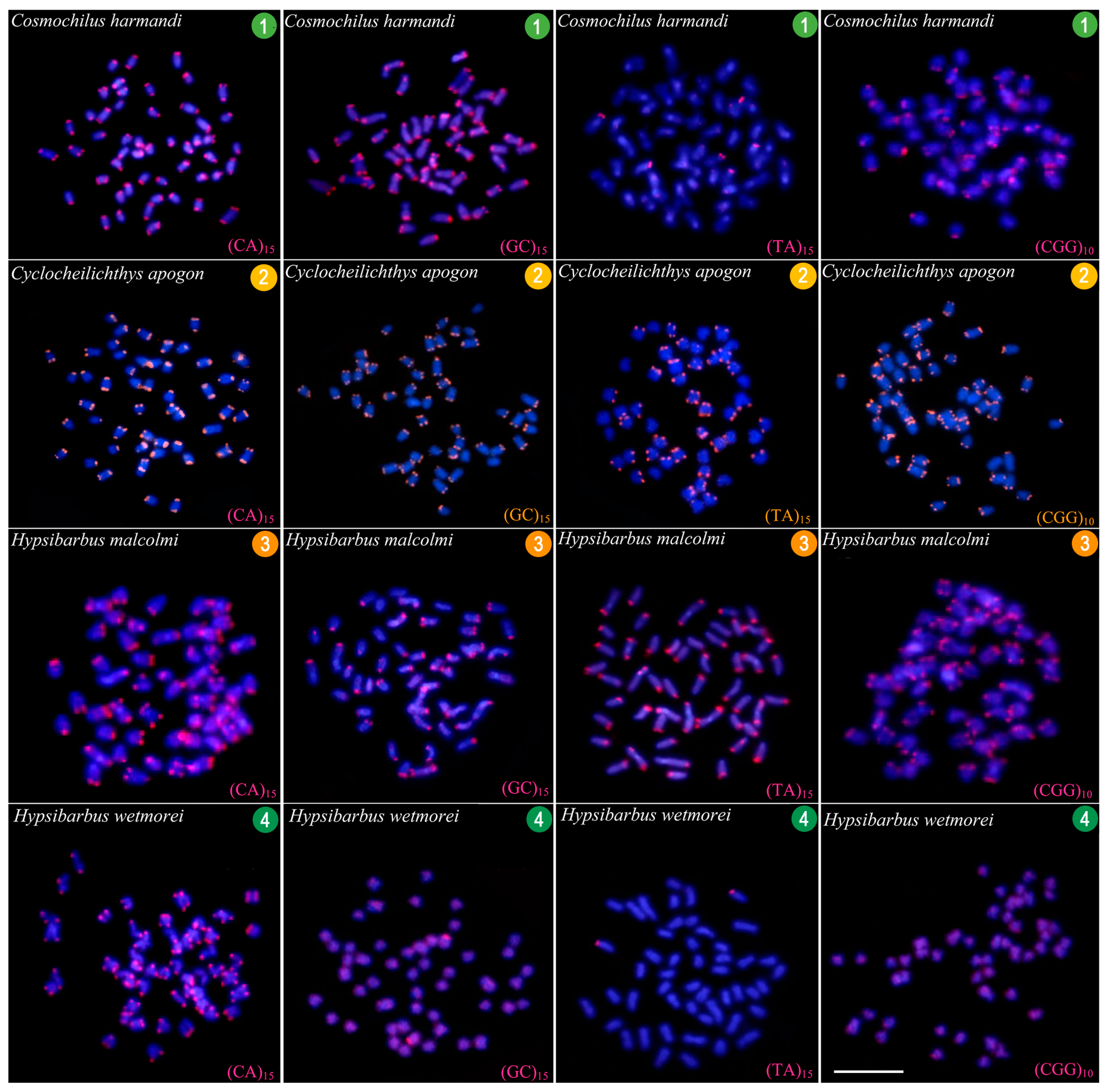
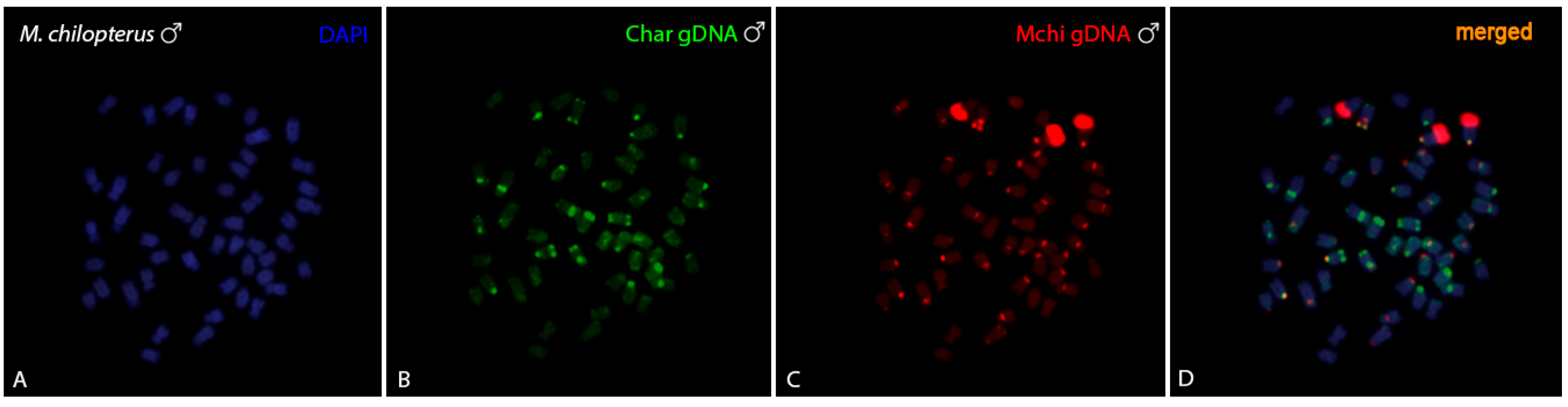
3.3. CGH-Studies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sado, T.; Hirt, M.V.; Pasco-Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenetics Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef]
- Yang, L.; Naylor, G.J.; Mayden, R.L. Deciphering reticulate evolution of the largest group of polyploid vertebrates, the subfamily cyprininae (Teleostei: Cypriniformes). Mol. Phylogenetics Evol. 2021, 166, 107323. [Google Scholar] [CrossRef]
- Wolf, U.; Ritter, H.; Atkin, N.B.; Ohno, S. Polyploidization in the fish family Cyprinidae, order Cypriniformes. Hum. Genet. 1969, 7, 240–244. [Google Scholar] [CrossRef]
- Sola, L.; Gornung, E. Classical and molecular cytogenetics of the zebrafish, Danio rerio (Cyprinidae, Cypriniformes): An overview. Genetica 2001, 111, 397–412. [Google Scholar] [CrossRef]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [PubMed]
- Šlechtová, V.; Bohlen, J.; Freyhof, J.; Ráb, P. Molecular phylogeny of the Southeast Asian freshwater fish family Botiidae (Teleostei: Cobitoidea) and the origin of polyploidy in their evolution. Mol. Phylogenetics Evol. 2006, 39, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Ráb, P.; Hnátková, E.; Majtánová, Z.; Šlechtová, V.B.; Bohlen, J. Karyotype Record for the Morphologically Derived, Rarely Collected, Freshwater Fish Ellopostoma mystax (Cypriniformes, Cobitoidea, Ellopostomatidae). Ichthyol. Herpetol. 2021, 109, 998–1001. [Google Scholar] [CrossRef]
- Bohlen, J.; Völker, M.; Rábová, M.; Ráb, P. Note on the banded karyotype of the enigmatic South Asian loach Vaillantella maassi (Cypriniformes). Ichthyol. Res. 2007, 55, 82–84. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. In Repetitive DNA Genome Dyn. Basel, 1st ed.; Garrido-Ramos, M.A., Ed.; Karger Publishers: Basel, Switzerland, 2012; Volume 7, pp. 197–221. [Google Scholar]
- Goes, C.A.G.; dos Santos, R.Z.; Aguiar, W.R.C.; Alves, D.C.V.; Silva, D.M.Z.D.A.; Foresti, F.; Oliveira, C.; Utsunomia, R.; Porto-Foresti, F. Revealing the Satellite DNA History in Psalidodon and Astyanax Characid Fish by Comparative Satellitomics. Front. Genet. 2022, 13, 884072. [Google Scholar] [CrossRef]
- Miura, I.; Shams, F.; Lin, S.M.; Cioffi, M.B.; Liehr, T.; Al-Rikabi, A.; Kuwana, C.; Srikulnath, K.; Higaki, Y.; Ezaz, T. Evolution of a multiple sex-chromosome system by three-sequential translocations among potential sex-chromosomes in the Taiwanese frog Odorrana swinhoana. Cells 2021, 10, 661. [Google Scholar] [CrossRef]
- Waters, P.D.; Patel, H.R.; Ruiz-Herrera, A.; Álvarez-González, L.; Lister, N.C.; Simakov, O.; Ezaz, T.; Kaur, P.; Frere, C.; Grützner, F.; et al. Microchromosomes are building blocks of bird, reptile, and mammal chromosomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2112494118. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, R.; de Souza, M.S.; Furo, I.d.O.; Romanov, M.N.; Gunski, R.J.; Garnero, A.d.V.; de Freitas, T.R.O.; de Oliveira, E.H.C.; O’connor, R.E.; Griffin, D.K. Interspecies Chromosome Mapping in Caprimulgiformes, Piciformes, Suliformes, and Trogoniformes (Aves): Cytogenomic Insight into Microchromosome Organization and Karyotype Evolution in Birds. Cells 2021, 10, 826. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.H.N.; Souza, F.H.S.; de Moraes, R.L.; Perez, M.F.; Sassi, F.D.M.C.; Viana, P.F.; Feldberg, E.; Ezaz, T.; Liehr, T.; Bertollo, L.A.C.; et al. The Genetic Differentiation of Pyrrhulina (Teleostei, Characiformes) Species is Likely Influenced by Both Geographical Distribution and Chromosomal Rearrangements. Front. Genet. 2022, 13, 869073. [Google Scholar] [CrossRef] [PubMed]
- Phimphan, S.; Chaiyasan, P.; Suwannapoom, C.; Reungsing, M.; Juntaree, S.; Tanomtong, A.; Supiwong, W. Comparative karyotype study of three Cyprinids (Cyprinidae, Cyprininae) in Thailand by classical cytogenetic and FISH techniques. Comp. Cytogenet. 2020, 14, 597–612. [Google Scholar] [CrossRef]
- Pereira, C.; Ráb, P.; Collares-Pereira, M. Chromosomes of Iberian Leuciscinae (Cyprinidae) Revisited: Evidence of Genome Restructuring in Homoploid Hybrids Using Dual-Color FISH and CGH. Cytogenet. Genome Res. 2013, 141, 143–152. [Google Scholar] [CrossRef]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Sember, A.; Majtánová, Z.; Ráb, P. Characterization of fish genomes by GISH and CGH. In Fish Cytogenetic Techniques, 1st ed.; Ozouf-Costaz, C., Pisano, E., Foresti, F., de Almeida, L., Eds.; CRC Press: London, UK, 2015; pp. 118–131. [Google Scholar]
- Doležálková, M.; Sember, A.; Marec, F.; Ráb, P.; Plötner, J.; Choleva, L. Is premeiotic genome elimination an exclusive mechanism for hemiclonal reproduction in hybrid males of the genus Pelophylax? BMC Genet. 2016, 17, 100. [Google Scholar] [CrossRef]
- Majtánová, Z.; Symonová, R.; Arias-Rodriguez, L.; Sallan, L.; Ráb, P. “Holostei versus Halecostomi” Problem: Insight from Cytogenetics of Ancient Nonteleost Actinopterygian Fish, Bowfin Amia calva: Molecular Cytogenetics of Amia calva. J. Exp. Zool. Part B Mol. Dev. Evol. 2017, 328, 620–628. [Google Scholar] [CrossRef]
- Majtánová, Z.; Unmack, P.J.; Prasongmaneerut, T.; Shams, F.; Srikulnath, K.; Ráb, P.; Ezaz, T. Evidence of Interspecific Chromosomal Diversification in Rainbowfishes (Melanotaeniidae, Teleostei). Genes 2020, 11, 818. [Google Scholar] [CrossRef]
- Shams, F.; Dyer, F.; Thompson, R.; Duncan, R.P.; Thiem, J.D.; Majtánová, Z.; Ezaz, T. Karyotypes and Sex Chromosomes in Two Australian Native Freshwater Fishes, Golden Perch (Macquaria ambigua) and Murray Cod (Maccullochella peelii) (Percichthyidae). Int. J. Mol. Sci. 2019, 20, 4244. [Google Scholar] [CrossRef]
- LeVan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for Centromeric Position on Chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Donsakul, T.; Magtoon, W.; Rangsiruji, A. Karyological studies of four Cyprinid fishes: Barbichthys nitidus, Mystacoleucus argenteus, Cychocheilichthys lagleri and Systomus sp. 1 from Thailand. In Proceedings of the 44rd Kasetsart University Annual Conference: Fisheries, Bangkok, Thailand, 30 January–2 February 2006; Kasetsart University: Bangkok, Thailand; pp. 469–476. [Google Scholar]
- Chantapan, T. Standardized karyotype and ideogram of cyprinid fishes (Subfamily: Cyprininae) in Thailand. Master’s Thesis, Khon Kaen University, Khon Kaen, Thailand, 2015. [Google Scholar]
- Seetapan, K. Karyotypes of sex fish species of the family Cyprinidae. In Proceedings of the 45th Kasetsart University Annual Conference: Fisheries, Bangkok, Thailand,, 30 January–2 February 2007; Kasetsart University: Bangkok, Thailand; pp. 749–758. [Google Scholar]
- Magtoon, W.; Arai, R. Karyotypes of five Puntius species and one Cyclocheilichtltys species (Pisces, Cyprinidae) from Thailand. Bull. Nat. Sci. Mus. Tokio 1989, 15, 167–175. [Google Scholar]
- Suzuki, A.; Taki, Y.; Mochizuki, M.; Hirata, J. Chromosomal speciation in Eurasian and Japanese Cyprinidae (Pisces, Cypriniformes). Cytobios 1995, 83, 171–186. [Google Scholar]
- Wu, G.M.; Zhu, X.P.; Hu, G.; Luo, J.R. The karyotype of Puntius gonionotus (Cyprinidae). Chin. J. Zool. 1991, 26, 20–21. [Google Scholar]
- Donsakul, T.; Poopitayasathaporn, A. Karyotype of Fifteen Species of Cyprinid Fishes (Family Cyprinidae) from Thailand. In Research Project; Srinakharinwirot University: Bangkok, Thailand, 2002. [Google Scholar]
- Khunda-Bukhsh, A.R.; Das, J.K. Cytogenetic analyses in eight species of teleostean fishes (Pisces): Karyotype, multiple Ag-NORs, sex chromosomes. Res. Rev. BioScience 2007, 1, 47–52. [Google Scholar]
- Arai, R. Fish Karyotypes: A Check List; Springer Science & Bussiness Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Donsakul, T.; Magtoon, W.; Rangsiruji, A. Karyotypes of four cyprinid fishes: Cyclocheilichthys repasson, Cosmochilus harmandi, Poropuntius deauratus and Sikukia gudgeri from Thailand. In Proceedings of the 43rd Kasetsart University Annual Conference: Fisheries, Kasetsart University, Bangkok, Thailand, 1–4 February 2005; pp. 344–351. [Google Scholar]
- Chaiyasan, P.; Supiwong, W.; Saenjundaeng, P.; Seetapan, K.; Pinmongkhonkul, S.; Tanomtong, A. A Report on Classical Cytogenetics of Hihgfin Barb Fish, Cyclocheilichthys armatus (Cypriniformes, Cyprinidae). Cytologia 2018, 83, 149–154. [Google Scholar] [CrossRef]
- Donsakul, T.; Rangsiruji, A.; Magtoon, W. Karyotypes of four cyprinid fishes: Poropuntius normani, Hypsibarbus malcolmi, Scaphognathops bandanensis and Henicorhynchus caudiguttatus from Thailand. In Proceedings of the 45th Kasetsart University Annual Conference: Fisheries, Kasetsart University, Bangkok, Thailand, 30 January–2 February 2007; pp. 740–748. [Google Scholar]
- Magtoon, W.; Arai, R. Karyotypes and distribution of nucleolus organizer regions in cyprinid fishes from Thailand. Jpn. J. Ichthyol. 1993, 40, 77–85. [Google Scholar] [CrossRef]
- Donsakul, T.; Magtoon, W. Karyotypes of four cyprinid fishes, Osteochilus melanopleura, Puntioplites proctozysron, Paralaubuca riveroi and Rasbora sumatrana, from Thailand. In Proceedings of the 33rd Kasetsart University Annual Conference: Fisheries, Kasetsart University, Bangkok, Thailand, 30 January–1 February 1995; pp. 128–138. [Google Scholar]
- Donsakul, T.; Magtoon, W. Karyotypes of two cyprinid fishes, Hypsibarbus wetmorei and Morulius chrysophekadion, from Thailand. In Proceedings of the 28th Congress on Science and Technology of Thailand, Queen Sirikit National Convention Centre, Bangkok, Thailand, 24–26 October 2002; p. 92. [Google Scholar]
- Piyapong, C. Karyotypes and Distribution of Nucleolus Organizer Regions in Four Cyprinid Species from Thailand. Master’s Thesis, Chulalongkorn University, Bangkok, Thailand, 1999. [Google Scholar]
- Arai, R.; Magtoon, W. Karyotypes of four cyprinid fishes from Thailand. Bull. Natl. Mus. Nat. Sci. 1991, 17, 183–188. [Google Scholar]
- Zan, R.G.; Song, Z.; Liu, W.G. Studies on karyotypes and nuclear DNA contents of some cyprinoid fishes, with notes on fish polyploids in China. In Proceedings of the 2nd International Conference Indo-Pacific Fishes, Tokyo, Japan, 1 October 1986; pp. 877–885. [Google Scholar]
- Donsakul, T.; Magtoon, W. Karyotypes of seven cyprinid fishes: Systomos binotatus, Puntius brevis, Poropuntius laoensis, Labiobarbus siamensis, Catlocarpiosiamensis, Tor tambroides and Probarbus jullieni from Thailand. Srinakharinwirot Sci. J. 2008, 24, 80–92. [Google Scholar]
- Zan, R.G.; Song, Z.; Liu, W.G. Studies of karyotypes of seven species of fish in Barbinae, with a discussion on identification of fish polyploids. Zool. Res. 1984, 5, 82–90. [Google Scholar]
- Supiwong, W.; Tanomtong, A.; Supanuam, P.; Jantarat, S.; Khakhong, S.; Sanoamuang, L. A discovery of nucleolar organizer regions (NORs) polymorphism and karyological analysis of Smith’ s Barb, Puntioplites proctozysron (Cypriniformes, Cyprinidae) in Thailand. Cytologia 2012, 77, 35–42. [Google Scholar] [CrossRef]
- Rainboth, W.J.; Vidthayanon, C.; Yen, M.D. Fishes of the Greater Mekong Ecosystem with Species List and Photographic Atlas, Miscellaneous Publications Museum of Zoology; University of Michigan: Ann Harbor, MI, USA, 2012; p. 314. [Google Scholar]
- Bertollo, L.A.C.; Cioffi, M.B.; Moreira-Filho, O. Direct chromosome preparation from freshwater teleost fishes. In Fish Cytogenetic Techniques: Ray-Fin Fishes and Chondrichthyans, 1st ed.; Ozouf-Costaz, C., Pisano, E., Foresti, F., Toledo, L.F.A., Eds.; RC Press: Boca Raton, FL, USA, 2015; pp. 21–26. [Google Scholar]
- Howell, W.M.; Black, D.A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Cioffi, M.B. Fish-FISH: Molecular cytogenetics in fish species. In Fluorescence In Situ Hybridization (FISH); Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 429–443. [Google Scholar] [CrossRef]
- Martins, C.; Ferreira, I.A.; Oliveira, C.; Foresti, F.; Galetti, P.M. A tandemly repetitive centromeric DNA sequence of the fish Hoplias malabaricus (Characiformes: Erythrinidae) is derived from 5S rDNA. Genetica 2006, 127, 133–141. [Google Scholar] [CrossRef]
- Cioffi, M.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L. Chromosomal Variability among Allopatric Populations of Erythrinidae Fish Hoplias malabaricus: Mapping of Three Classes of Repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite accumulation on the Y chromosome in Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Zwick, M.S.; Hanson, R.E.; Islam-Faridi, M.N.; Stelly, D.M.; Wing, R.A.; Price, H.J.; McKnight, T.D. A rapid procedure for the isolation of C0t-1 DNA from plants. Genome 1997, 40, 138–142. [Google Scholar] [CrossRef]
- Saenjundaeng, P.; Supiwong, W.; Sassi, F.M.C.; Bertollo, L.A.C.; Rab, P.; Kretschmer, R.; Tanomtong, A.; Suwannapoom, C.; Reungsing, M.; Cioffi, M.D.B. Chromosomes of Asian cyprinid fishes: Variable karyotype patterns and evolutionary trends in the genus Osteochilus (Cyprinidae, Labeoninae, “Osteochilini”). Genet. Mol. Biol. 2020, 43, e20200195. [Google Scholar] [CrossRef]
- Pereira, R.; Pereira, V.; Gomes, I.; Tomas, C.; Morling, N.; Amorim, A.; Prata, M.J.; Carracedo, Á.; Gusmão, L. A method for the analysis of 32 X chromosome insertion deletion polymorphisms in a single PCR. Int. J. Leg. Med. 2011, 126, 97–105. [Google Scholar] [CrossRef]
- Derenzini, M. The AgNORs. Micron 2000, 31, 117–120. [Google Scholar] [CrossRef]
- Gornung, E. Twenty Years of Physical Mapping of Major Ribosomal RNA Genes across the Teleosts: A Review of Research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Rebordinos, L.; Cross, I.; Merlo, A. High Evolutionary Dynamism in 5S rDNA of Fish: State of the Art. Cytogenet. Genome Res. 2013, 141, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.; Bertollo, L.A.C.; Villa, M.A.; de Oliveira, E.A.; Tanomtong, A.; Yano, C.F.; Supiwong, W.; Chaveerach, A. Genomic Organization of Repetitive DNA Elements and Its Implications for the Chromosomal Evolution of Channid Fishes (Actinopterygii, Perciformes). PLoS ONE 2015, 10, e0130199. [Google Scholar] [CrossRef]
- Symonová, R.; Howell, W.M. Vertebrate Genome Evolution in the Light of Fish Cytogenomics and rDNAomics. Genes 2018, 9, 96. [Google Scholar] [CrossRef]
- Salim, D.; Gerton, J.L. Ribosomal DNA instability and genome adaptability. Chromosom. Res. 2019, 27, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Roa, F.; Guerra, M. Non-Random Distribution of 5S rDNA Sites and Its Association with 45S rDNA in Plant Chromosomes. Cytogenet. Genome Res. 2015, 146, 243–249. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2017, 127, 141–150. [Google Scholar] [CrossRef]
- Fujiwara, A.; Abe, S.; Yamaha, E.; Yamazaki, F.; Yoshida, M.C. Uniparental chromosome elimination in the early embryogenesis of the inviable salmonid hybrids between masu salmon female and rainbow trout male. Chromosoma 1997, 106, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-P.; Gui, J.-F. Identification of genome organization in the unusual allotetraploid form of Carassius auratus gibelio. Aquaculture 2007, 265, 109–117. [Google Scholar] [CrossRef]
- Valente, G.T.; Schneider, C.H.; Gross, M.C.; Feldberg, E.; Martins, C. Comparative cytogenetics of cichlid fishes through genomic in-situ hybridization (GISH) with emphasis on Oreochromis niloticus. Chromosom. Res. 2009, 17, 791–799. [Google Scholar] [CrossRef]
- Ditcharoen, S.; Bertollo, L.A.C.; Ráb, P.; Hnátková, E.; Molina, W.F.; Liehr, T.; Tanomong, A.; Triantaphyllidis, C.; Ozouf-Costaz, C.; Tongnunui, S.; et al. Genomic organization of repetitive DNA elements and extensive karyotype diversity of silurid catfishes (Teleostei: Siluriformes): A comparative cytogenetic approach. Int. J. Mol. Sci. 2019, 20, 3545. [Google Scholar] [CrossRef]
- Rampin, M.; Bi, K.; Bogart, J.P.; Collares-Pereira, M.J. Identifying parental chromosomes and genomic rearrangements in animal hybrid complexes of species with small genome size using genomic in situ hybridization (GISH). Comp. Cytogenet. 2012, 6, 287–300. [Google Scholar] [CrossRef]
- Knytl, M.; Kalous, L.; Symonová, R.; Rylková, K.; Ráb, P. Chromosome studies of European cyprinid fishes: Cross-species painting reveals natural allotetraploid origin of a Carassius female with 206 chromosomes. Cytogenet. Genome Res. 2013, 139, 276–283. [Google Scholar] [CrossRef]
- Ráb, P.; Collares-Pereira, M.J. Chromosomes of European cyprinid fishes (Cyprinidae, Cypriniformes). A review. Folia Zool. 1995, 44, 193–214. [Google Scholar]
- Boroń, A.; Spoz, A.; Porycka, K.; Karolewska, M.; Ito, D.; Abe, S.; Kirtiklis, L.; Juchno, D. Molecular cytogenetic analysis of the crucian carp, Carassius carassius (Linnaeus, 1758) (Teleostei, Cyprinidae), using chromosome staining and fluorescence in situ hybridisation with rDNA probes. Comp. Cytogenet. 2014, 8, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Knytl, M.; Kalous, L.; Rylková, K.; Choleva, L.; Merilä, J.; Ráb, P. Morphologically indistinguishable hybrid Carassius female with 156 chromosomes: A threat for the threatened crucian carp, C. carassius, L. PLoS ONE 2018, 13, e0190924. [Google Scholar] [CrossRef]
- Saenjundaeng, P.; Cioffi, M.B.; de Oliveira, E.A.; Tanomtong, A.; Supiwong, W.; Phimphan, S.; Collares-Pereira, M.J.; Sember, A.; Bertollo, L.A.C.; Liehr, T.; et al. Chromosomes of Asian cyprinid fishes: Cytogenetic analysis of two representatives of small paleotetraploid tribe Probarbini. Mol. Cytogenet. 2018, 11, 1–9. [Google Scholar] [CrossRef]
- Nirchio, M.; Rossi, A.R.; Foresti, F.; Oliveira, C. Chromosome evolution in fishes: A new challenging proposal from Neotropical species. Neotrop. Ichtchyol. 2014, 12, 761–770. [Google Scholar] [CrossRef]
- Jacobina, U.P.; Martinez, P.A.; Cioffi, M.B.; Garcia, J.; Bertollo, L.A.C.; Molina, W.F. Morphological and karyotypic differentiation in Caranx lugubris (Perciformes: Carangidae) in the St. Peter and St. Paul Archipelago, mid-Atlantic Ridge. Helgol. Mar. Res. 2014, 68, 17–25. [Google Scholar] [CrossRef]
- Betancur-R, R.; Wiley, E.O.; Arratia, G.; Acero, A.; Bailly, N.; Miya, M.; Lecointre, G.; Ortí, G. Phylogenetic classification of bony fishes. BMC Evol. Biol. 2017, 17, 162. [Google Scholar] [CrossRef]








| Species | Hydrographic Basin | n |
|---|---|---|
| Cosmochilus harmandi | Chao Phraya (site 1) | 07♀; 06♂ |
| Cyclocheilichthys apogon | Mae Klong (site 2) | 08♀; 11♂ |
| Hypsibarbus malcolmi | Mekong (site 3) | 09♀; 09♂ |
| Hypsibarbus wetmorei | Mekong (site 4) | 07♀; 05♂ |
| Mystacoleucus chilopterus | Mae Klong (site 5) | 06♀; 08♂ |
| Mystacoleucus ectypus | Mae Klong (site 6) | 06♀; 06♂ |
| Puntioplites proctozysron | Mae Klong (site 7) | 07♀; 06♂ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khensuwan, S.; Sassi, F.d.M.C.; Moraes, R.L.R.; Jantarat, S.; Seetapan, K.; Phintong, K.; Thongnetr, W.; Kaewsri, S.; Jumrusthanasan, S.; Supiwong, W.; et al. Chromosomes of Asian Cyprinid Fishes: Genomic Differences in Conserved Karyotypes of ‘Poropuntiinae’ (Teleostei, Cyprinidae). Animals 2023, 13, 1415. https://doi.org/10.3390/ani13081415
Khensuwan S, Sassi FdMC, Moraes RLR, Jantarat S, Seetapan K, Phintong K, Thongnetr W, Kaewsri S, Jumrusthanasan S, Supiwong W, et al. Chromosomes of Asian Cyprinid Fishes: Genomic Differences in Conserved Karyotypes of ‘Poropuntiinae’ (Teleostei, Cyprinidae). Animals. 2023; 13(8):1415. https://doi.org/10.3390/ani13081415
Chicago/Turabian StyleKhensuwan, Sudarat, Francisco de M. C. Sassi, Renata L. R. Moraes, Sitthisak Jantarat, Kriengkrai Seetapan, Krit Phintong, Weera Thongnetr, Sarawut Kaewsri, Sarun Jumrusthanasan, Weerayuth Supiwong, and et al. 2023. "Chromosomes of Asian Cyprinid Fishes: Genomic Differences in Conserved Karyotypes of ‘Poropuntiinae’ (Teleostei, Cyprinidae)" Animals 13, no. 8: 1415. https://doi.org/10.3390/ani13081415
APA StyleKhensuwan, S., Sassi, F. d. M. C., Moraes, R. L. R., Jantarat, S., Seetapan, K., Phintong, K., Thongnetr, W., Kaewsri, S., Jumrusthanasan, S., Supiwong, W., Rab, P., Tanomtong, A., Liehr, T., & Cioffi, M. B. (2023). Chromosomes of Asian Cyprinid Fishes: Genomic Differences in Conserved Karyotypes of ‘Poropuntiinae’ (Teleostei, Cyprinidae). Animals, 13(8), 1415. https://doi.org/10.3390/ani13081415