
Basal Intestinal Morphology, Immunolocalization of Leptin and Ghrelin and Their Receptors in Newborn Wistar Rats after Prenatal Exposure to Fumonisins
 , ,
, ,  , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Fumonisins Stock Solution
2.2. Animals and Experimental Design
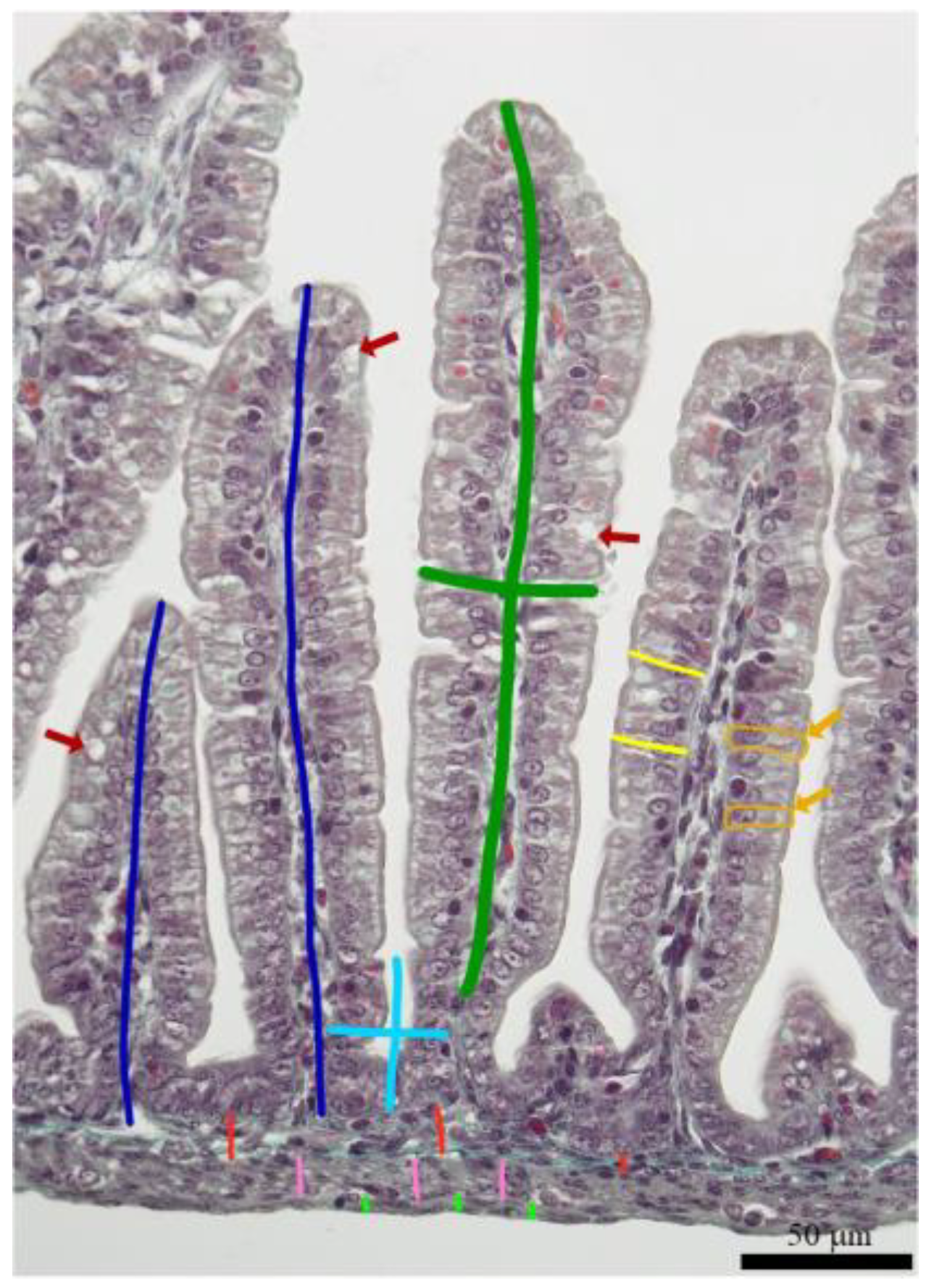
2.3. Tissue Sampling and Morphometrical Analysis
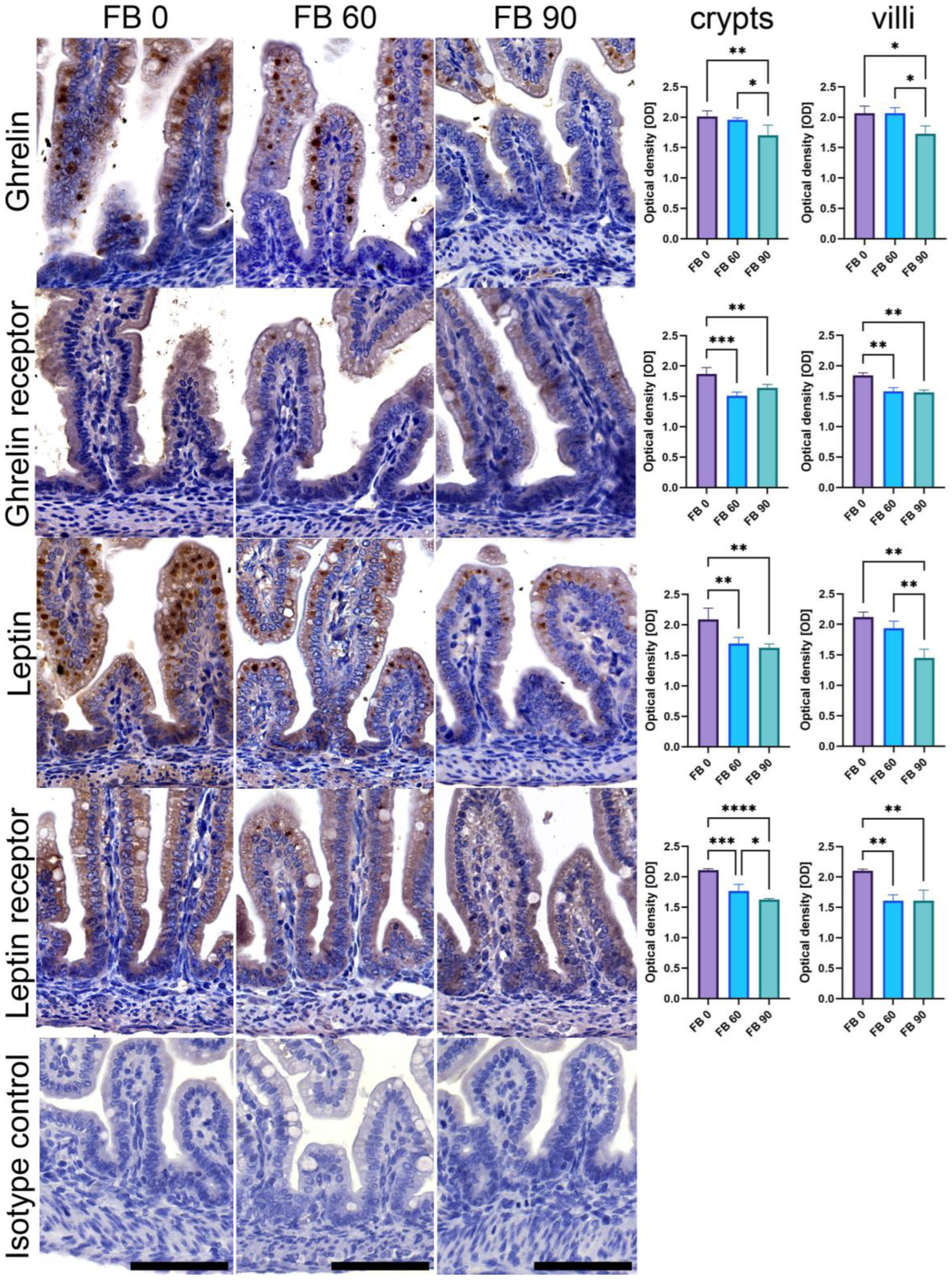
2.4. Leptin, Ghrelin and Their Receptors Immunostaining
2.5. Statistical Analysis
3. Results
3.1. Duodenal Basal Morphology
3.2. Ghrelin, Leptin, and Their Receptors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kosicki, R.; Twarużek, M.; Kannenberg, K.; Grajewski, J. Contamination of Acorns of Pedunculate Oak (Quercus robur L.), as Feed Material, by Moulds and Mycotoxins. Ann. Anim. Sci. 2021, 21, 977–990. [Google Scholar] [CrossRef]
- Placinta, C.M.; D’Mello, J.P.F.; Macdonald, A.M.C. A Review of Worldwide Contamination of Cereal Grains and Animal Feed with Fusarium Mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin Occurrence in Feed and Feed Raw Materials Worldwide: Long-Term Analysis with Special Focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef]
- Stockmann-Juvala, H.; Savolainen, K. A Review of the Toxic Effects and Mechanisms of Action of Fumonisin B1. Hum. Exp. Toxicol. 2008, 27, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Shima, H.; Vesonder, R.F.; Tokuda, H.; Nishino, H.; Katoh, S.; Tamura, S.; Sugimura, T.; Nagao, M. Inhibition of Protein Serine/Threonine Phosphatases by Fumonisin B1, a Mycotoxin. Biochem. Biophys. Res. Commun. 1996, 220, 160–165. [Google Scholar] [CrossRef]
- Riley, R.T.; Enongene, E.; Voss, K.A.; Norred, W.P.; Meredith, F.I.; Sharma, R.P.; Spitsbergen, J.; Williams, D.E.; Carlson, D.B.; Merrill, A.H. Sphingolipid Perturbations as Mechanisms for Fumonisin Carcinogenesis. Environ. Health Perspect. 2001, 109, 301–308. [Google Scholar] [CrossRef]
- Li, Y.; Fan, Y.; Xia, B.; Xiao, Q.; Wang, Q.; Sun, W.; Zhang, H.; He, C. The Immunosuppressive Characteristics of FB1 by Inhibition of Maturation and Function of BMDCs. Int. Immunopharm. 2017, 47, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Kang, S.C. Endoplasmic Reticulum Stress-Mediated Autophagy Activation Attenuates Fumonisin B1 Induced Hepatotoxicity in Vitro and in Vivo. Food Chem. Toxicol. 2017, 110, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, W.C.; Abel, S.; Smuts, C.M.; Marnewick, J.; Marasas, W.F.; Lemmer, E.R.; Ramljak, D. Fumonisin-Induced Hepatocarcinogenesis: Mechanisms Related to Cancer Initiation and Promotion. Environ. Health Perspect. 2001, 109, 291–300. [Google Scholar] [CrossRef]
- Chen, J.; Wei, Z.; Wang, Y.; Long, M.; Wu, W.; Kuca, K. Fumonisin B1: Mechanisms of Toxicity and Biological Detoxification Progress in Animals. Food Chem. Toxicol. 2021, 149, 111977. [Google Scholar] [CrossRef]
- Smith, G.W. Fumonisins. In Veterinary Toxicology: Basic and Clinical Principles; Elsevier: London, UK; Academic Press: London, UK, 2018; pp. 1003–1018. ISBN 978-0-12-811410-0. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Leblanc, J.; Nielsen, E.; et al. Assessment of Information as Regards the Toxicity of Fumonisins for Pigs, Poultry and Horses. EFSA J. 2022, 20, e07534. [Google Scholar] [CrossRef]
- Loh, Z.H.; Ouwerkerk, D.; Klieve, A.V.; Hungerford, N.L.; Fletcher, M.T. Toxin Degradation by Rumen Microorganisms: A Review. Toxins 2020, 12, 664. [Google Scholar] [CrossRef] [PubMed]
- Bryła, M.; Roszko, M.; Szymczyk, K.; Jędrzejczak, R.; Obiedziński, M.W.; Sękul, J. Fumonisins in Plant-Origin Food and Fodder—A Review. Food Addit. Contam. Part A 2013, 30, 1626–1640. [Google Scholar] [CrossRef] [PubMed]
- The Commission of the European Communities. EC Commission Regulation (EC) No. 856/2005 of 6 June 2005 Amending Regulation (EC) No. 466/2001 as Regards Fusarium Toxins. OJEU 2005, L 143, 3–8. [Google Scholar]
- The Commission of the European Communities. EC Commission Regulation (EC) No 401/2006 of 23 February 2006 Laying down the Methods of Sampling and Analysis for the Official Control of the Levels of Mycotoxins in Foodstuffs. OJEU 2006, L 70, 12–34. [Google Scholar]
- U.S. Food & Drug Administration. FDA Guidance for Industry: Fumonisin Levels in Human Foods and Animal Feeds. Available online: www.fda.gov/RegulatoryInformation/Guidances/ucm109231.htm (accessed on 21 April 2023).
- Tomaszewska, E.; Rudyk, H.; Świetlicka, I.; Hułas-Stasiak, M.; Donaldson, J.; Arczewska, M.; Muszyński, S.; Dobrowolski, P.; Puzio, I.; Kushnir, V.; et al. The Influence of Prenatal Fumonisin Exposure on Bone Properties, as Well as OPG and RANKL Expression and Immunolocalization, in Newborn Offspring Is Sex and Dose Dependent. Int. J. Mol. Sci. 2021, 22, 13234. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Rudyk, H.; Wojtysiak, D.; Donaldson, J.; Muszyński, S.; Arciszewski, M.B.; Lisova, N.; Brezvyn, O.; Puzio, I.; Abramowicz, B.; et al. Basal Blood Morphology, Serum Biochemistry, and the Liver and Muscle Structure of Weaned Wistar Rats Prenatally Exposed to Fumonisins. Animals 2022, 12, 2353. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Rudyk, H.; Świetlicka, I.; Hułas-Stasiak, M.; Donaldson, J.; Arczewska, M.; Muszyński, S.; Dobrowolski, P.; Mielnik-Błaszczak, M.; Arciszewski, M.B.; et al. Trabecular Bone Parameters, TIMP-2, MMP-8, MMP-13, VEGF Expression and Immunolocalization in Bone and Cartilage in Newborn Offspring Prenatally Exposed to Fumonisins. Int. J. Mol. Sci. 2021, 22, 12528. [Google Scholar] [CrossRef] [PubMed]
- Kras, K.; Rudyk, H.; Muszyński, S.; Tomaszewska, E.; Dobrowolski, P.; Kushnir, V.; Muzyka, V.; Brezvyn, O.; Arciszewski, M.B.; Kotsyumbas, I. Morphology and Chemical Coding of Rat Duodenal Enteric Neurons Following Prenatal Exposure to Fumonisins. Animals 2022, 12, 1055. [Google Scholar] [CrossRef]
- Kyei, N.N.A.; Boakye, D.; Gabrysch, S. Maternal Mycotoxin Exposure and Adverse Pregnancy Outcomes: A Systematic Review. Mycotoxin Res. 2020, 36, 243–255. [Google Scholar] [CrossRef]
- Gelineau-van Waes, J.; Voss, K.A.; Stevens, V.L.; Speer, M.C.; Riley, R.T. Maternal Fumonisin Exposure as a Risk Factor for Neural Tube Defects. Adv. Food Nutr. Res. 2009, 56, 145–181. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Barker, D.J. Fetal Programming and Adult Health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of In Utero and Early-Life Conditions on Adult Health and Disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, E.; Rudyk, H.; Dobrowolski, P.; Donaldson, J.; Świetlicka, I.; Puzio, I.; Kamiński, D.; Wiącek, D.; Kushnir, V.; Brezvyn, O.; et al. Changes in the Intestinal Histomorphometry, the Expression of Intestinal Tight Junction Proteins, and the Bone Structure and Liver of Pre-Laying Hens Following Oral Administration of Fumonisins for 21 Days. Toxins 2021, 13, 375. [Google Scholar] [CrossRef]
- Rudyk, H.; Tomaszewska, E.; Kotsyumbas, I.; Muszyński, S.; Tomczyk-Warunek, A.; Szymańczyk, S.; Dobrowolski, P.; Wiącek, D.; Kamiński, D.; Brezvyn, O. Bone Homeostasis in Experimental Fumonisins Intoxication of Rats. Ann. Anim. Sci. 2019, 19, 403–419. [Google Scholar] [CrossRef]
- Rudyk, H.; Tomaszewska, E.; Arciszewski, M.B.; Muszyński, S.; Tomczyk-Warunek, A.; Dobrowolski, P.; Donaldson, J.; Brezvyn, O.; Kotsyumbas, I. Histomorphometrical Changes in Intestine Structure and Innervation Following Experimental Fumonisins Intoxication in Male Wistar Rats. Pol. J. Vet. Sci. 2020, 23, 77–88. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Layton, C. Connective and Other Mesenchymal Tissues with Their Stains. In Bancroft’s Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2019; pp. 153–175. ISBN 978-0-7020-6887-4. [Google Scholar]
- Iwaniak, P.; Tomaszewska, E.; Muszyński, S.; Marszałek-Grabska, M.; Pierzynowski, S.G.; Dobrowolski, P. Dietary Alpha-Ketoglutarate Partially Abolishes Adverse Changes in the Small Intestine after Gastric Bypass Surgery in a Rat Model. Nutrients 2022, 14, 2062. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Prost, Ł.; Dobrowolski, P.; Chand, D.K.P.; Donaldson, J.; Czech, A.; Klebaniuk, R.; Fabjanowska, J.; Muszyński, S. Prenatal Programming of the Small Intestine in Piglets: The Effect of Supplementation with 3-Hydroxy-3-Methylbutyric Acid (HMB) in Pregnant Sows on the Structure of Jejunum of Their Offspring. Ann. Anim. Sci. 2022, 22, 613–623. [Google Scholar] [CrossRef]
- Sakamoto, K.; Hirose, H.; Onizuka, A.; Hayashi, M.; Futamura, N.; Kawamura, Y.; Ezaki, T. Quantitative Study of Changes in Intestinal Morphology and Mucus Gel on Total Parenteral Nutrition in Rats. J. Surg. Res. 2000, 94, 99–106. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Prost, Ł.; Chand, D.K.P.; Donaldson, J.; Winiarczyk, D.; Jarosz, Ł.; Ciszewski, A.; Czech, A.; Muszyński, S. The Effect of Supplementation with β-Hydroxy-β-Methylbutyric Acid (HMB) to Pregnant Sows on the Mucosal Structure, Immunolocalization of Intestinal Barrier Proteins, VIP and Leptin in the Large Intestine in Their Offspring. Ann. Anim. Sci. 2023, 23, 87–96. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An Open Source Plugin for the Quantitative Evaluation and Automated Scoring of Immunohistochemistry Images of Human Tissue Samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [PubMed]
- Seyed Jafari, S.M.; Hunger, R.E. IHC Optical Density Score: A New Practical Method for Quantitative Immunohistochemistry Image Analysis. Appl. Immunohistochem. Mol. Morphol. 2017, 25, e12–e13. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.-H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, Mechanism of Action and Toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Śliwa, E.; Dobrowolski, P.; Tatara, M.R.; Piersiak, T.; Siwicki, A.; Rokita, E.; Pierzynowski, S.G. Alpha-Ketoglutarate Protects the Liver of Piglets Exposed during Prenatal Life to Chronic Excess of Dexamethasone from Metabolic and Structural Changes. J. Anim. Physiol. Anim. Nutr. 2009, 93, 192–202. [Google Scholar] [CrossRef]
- Collins, T.F.X.; Shackelford, M.E.; Sprando, R.L.; Black, T.N.; Láborde, J.B.; Hansen, D.K.; Eppley, R.M.; Trucksess, M.W.; Howard, P.C.; Bryant, M.A.; et al. Effects of Fumonisin B1 in Pregnant Rats. Food Chem. Toxicol. 1998, 36, 397–408. [Google Scholar] [CrossRef]
- Collins, T.F.X.; Sprando, R.L.; Black, T.N.; Shackelford, M.E.; Laborde, J.B.; Hansen, D.K.; Eppley, R.M.; Trucksess, M.W.; Howard, P.C.; Bryant, M.A.; et al. Effects of Fumonisin B1 in Pregnant Rats. Part 2. Food Chem. Toxicol. 1998, 36, 673–685. [Google Scholar] [CrossRef]
- Reusens-Billen, B.; Remacle, C.; Hoet, J.J. The Development of the Fetal Rat Intestine and Its Reaction to Maternal Diabetes. Diabetes Res. Clin. Pract. 1989, 6, 199–211. [Google Scholar] [CrossRef]
- Miller, D.L. Rat Small Intestine: Development, Composition and Effects of Perfusion. Digest. Dis. Sci. 1971, 16, 247–254. [Google Scholar] [CrossRef]
- Shames, B. Anatomy and Physiology of the Duodenum. In Shackelford’s Surgery of the Alimentary Tract; Yeo, C.J., Ed.; Elsevier: Philadelphia, PA, USA, 2019; pp. 786–803. ISBN 978-0-323-40232-3. [Google Scholar]
- Brandtzaeg, P.; Kiyono, H.; Pabst, R.; Russell, M.W. Terminology: Nomenclature of Mucosa-Associated Lymphoid Tissue. Mucosal Immunol. 2008, 1, 31–37. [Google Scholar] [CrossRef] [PubMed]
- MacNaughton, W.K. Mechanisms and Consequences of Intestinal Inflammation. In Physiology of the Gastrointestinal Tract; Elsevier: Amsterdam, The Netherlands, 2006; pp. 1115–1135. ISBN 978-0-12-088394-3. [Google Scholar]
- Dickson, I. Enterocyte Zonation along the Intestinal Villus Reveals a Division of Labour. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 717. [Google Scholar] [CrossRef] [PubMed]
- Cera, K.R.; Mahan, D.C.; Cross, R.F.; Reinhart, G.A.; Whitmoyer, R.E. Effect of Age, Weaning and Postweaning Diet on Small Intestinal Growth and Jejunal Morphology in Young Swine. J. Anim. Sci. 1988, 66, 574. [Google Scholar] [CrossRef] [PubMed]
- Piva, A.; Casadei, G.; Pagliuca, G.; Cabassi, E.; Galvano, F.; Solfrizzo, M.; Riley, R.T.; Diaz, D.E. Activated Carbon Does Not Prevent the Toxicity of Culture Material Containing Fumonisin B1 When Fed to Weanling Piglets. J. Anim. Sci. 2005, 83, 1939–1947. [Google Scholar] [CrossRef]
- Javed, T.; Bunte, R.M.; Dombrink-Kurtzman, M.A.; Richard, J.L.; Bennett, G.A.; Côté, L.M.; Buck, W.B. Comparative Pathologic Changes in Broiler Chicks on Feed Amendedwith Fusarium Proliferatum Culture Material or Purified FumonisinB1 and Moniliformin. Mycopathologia 2005, 159, 553–564. [Google Scholar] [CrossRef]
- Grenier, B.; Bracarense, A.-P.F.L.; Schwartz, H.E.; Trumel, C.; Cossalter, A.-M.; Schatzmayr, G.; Kolf-Clauw, M.; Moll, W.-D.; Oswald, I.P. The Low Intestinal and Hepatic Toxicity of Hydrolyzed Fumonisin B1 Correlates with Its Inability to Alter the Metabolism of Sphingolipids. Biochem. Pharmacol. 2012, 83, 1465–1473. [Google Scholar] [CrossRef]
- Rauber, R.H.; Oliveira, M.S.; Mallmann, A.O.; Dilkin, P.; Mallmann, C.A.; Giacomini, L.Z.; Nascimento, V.P. Effects of Fumonisin B1 on Selected Biological Responses and Performance of Broiler Chickens. Pesq. Vet. Bras. 2013, 33, 1081–1086. [Google Scholar] [CrossRef]
- Antonissen, G.; Croubels, S.; Pasmans, F.; Ducatelle, R.; Eeckhaut, V.; Devreese, M.; Verlinden, M.; Haesebrouck, F.; Eeckhout, M.; De Saeger, S.; et al. Fumonisins Affect the Intestinal Microbial Homeostasis in Broiler Chickens, Predisposing to Necrotic Enteritis. Vet. Res. 2015, 46, 98. [Google Scholar] [CrossRef]
- Matsuda, K.; Ono, S.; Tanaka, I.; Inoue, M.; Kinowaki, S.; Ishikawa, M.; Tsuda, M.; Yamamoto, K.; Shimizu, Y.; Takahashi, S.; et al. Histological and Magnified Endoscopic Evaluation of Villous Atrophy in Gastrointestinal Graft-versus-Host Disease. Ann. Hematol. 2020, 99, 1121–1128. [Google Scholar] [CrossRef]
- Robert, H.; Payros, D.; Pinton, P.; Théodorou, V.; Mercier-Bonin, M.; Oswald, I.P. Impact of Mycotoxins on the Intestine: Are Mucus and Microbiota New Targets? J. Toxicol. Environ. Health B Crit. Rev. 2017, 20, 249–275. [Google Scholar] [CrossRef]
- Hooper, L.V. Epithelial Cell Contributions to Intestinal Immunity. Adv. Immunol. 2015, 126, 129–172. [Google Scholar] [CrossRef] [PubMed]
- Vancamelbeke, M.; Vermeire, S. The Intestinal Barrier: A Fundamental Role in Health and Disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Han, T.-S.; Son, M.-Y. Intestinal Morphogenesis in Development, Regeneration, and Disease: The Potential Utility of Intestinal Organoids for Studying Compartmentalization of the Crypt-Villus Structure. Front. Cell Dev. Biol. 2020, 8, 593969. [Google Scholar] [CrossRef] [PubMed]
- Hartsock, A.; Nelson, W.J. Adherens and Tight Junctions: Structure, Function and Connections to the Actin Cytoskeleton. Biochim. Biophys. Acta 2008, 1778, 660–669. [Google Scholar] [CrossRef]
- Dixit, V.D.; Schaffer, E.M.; Pyle, R.S.; Collins, G.D.; Sakthivel, S.K.; Palaniappan, R.; Lillard, J.W.; Taub, D.D. Ghrelin Inhibits Leptin- and Activation-Induced Proinflammatory Cytokine Expression by Human Monocytes and T Cells. J. Clin. Investig. 2004, 114, 57–66. [Google Scholar] [CrossRef]
- Spiridon, I.; Ciobanu, D.; Giușcă, S.; Căruntu, I. Ghrelin and Its Role in Gastrointestinal Tract Tumors (Review). Mol. Med. Rep. 2021, 24, 663. [Google Scholar] [CrossRef]
- Mani, B.K.; Shankar, K.; Zigman, J.M. Ghrelin’s Relationship to Blood Glucose. Endocrinology 2019, 160, 1247–1261. [Google Scholar] [CrossRef]
- Williams, J.; Mobarhan, S. A Critical Interaction: Leptin and Ghrelin. Nutr. Rev. 2003, 61, 391–393. [Google Scholar] [CrossRef]
- O’Connor, A.; O’Moráin, C. Digestive Function of the Stomach. Dig. Dis. 2014, 32, 186–191. [Google Scholar] [CrossRef]
- Konturek, S.J.; Konturek, J.W.; Pawlik, T.; Brzozowski, T. Brain-Gut Axis and Its Role in the Control of Food Intake. J. Physiol. Pharmacol. 2004, 55, 137–154. [Google Scholar]
- Gomes, D.; Le, L.; Perschbacher, S.; Haas, N.A.; Netz, H.; Hasbargen, U.; Delius, M.; Lange, K.; Nennstiel, U.; Roscher, A.A.; et al. Predicting the Earliest Deviation in Weight Gain in the Course towards Manifest Overweight in Offspring Exposed to Obesity in Pregnancy: A Longitudinal Cohort Study. BMC Med. 2022, 20, 156. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Dependent Variable | FB (mg/kg b.w.) | SEM | p-Value | p-Level | |||
|---|---|---|---|---|---|---|---|
| 0 | 60 | 90 | Linear | Quadratic | |||
| Longitudinal m. lamina thickness, μm | 10.27 b | 5.72 a | 7.07 a | 0.653 | <0.001 | <0.001 | 0.002 |
| Circular m. lamina thickness, μm | 20.06 b | 12.68 a | 12.95 a | 1.823 | 0.018 | 0.008 | 0.107 |
| Submucosa thickness, μm | 12.63 | 10.89 | 9.54 | 1.199 | 0.223 | 0.090 | 0.898 |
| Mucosa thickness, μm | 246.78 | 210.66 | 235.79 | 24.661 | 0.572 | 0.622 | 0.320 |
| Crypt depth, μm | 37.1 | 34.87 | 39.41 | 3.350 | 0.640 | 0.751 | 0.422 |
| Crypt thickness, µm | 38.69 | 38.01 | 33.61 | 2.385 | 0.292 | 0.195 | 0.532 |
| Villus length, µm | 250.94 | 175.33 | 196.38 | 22.122 | 0.074 | 0.058 | 0.094 |
| Villus thickness, µm | 84.63 b | 60.52 a | 56.98 a | 5.264 | 0.004 | 0.001 | 0.131 |
| Total number of villi, /mm | 11.21 | 13.02 | 11.79 | 0.788 | 0.283 | 0.431 | 0.136 |
| Total crypt number, /mm | 13.73 b | 11.44 a | 14.45 b | 0.407 | <0.001 | 0.826 | <0.001 |
| Villus length/crypt depth ratio | 6.29 | 5.58 | 5.13 | 0.986 | 0.778 | 0.415 | 0.917 |
| Enterocyte, per 100 µm of villus | 17.04 | 17.06 | 15.82 | 0.622 | 0.298 | 0.244 | 0.419 |
| Goblet cells, per 100 µm of villus | 3.923 | 3.752 | 3.635 | 0.344 | 0.840 | 0.563 | 0.950 |
| Villus epithelium thickness, µm | 17.16 b | 13.52 a | 12.40 a | 0.824 | 0.003 | <0.001 | 0.231 |
| Dependent Variable | FB (mg/kg b.w.) | SEM | p-Value | p-Level | |||
|---|---|---|---|---|---|---|---|
| 0 | 60 | 90 | Linear | Quadratic | |||
| Longitudinal m. lamina thickness, μm | 12.15 | 9.87 | 8.26 | 1.319 | <0.001 | 0.054 | 0.837 |
| Circular m. lamina thickness, μm | 14.95 | 16.50 | 13.72 | 1.500 | 0.441 | 0.722 | 0.256 |
| Submucosa thickness, μm | 12.03 | 9.70 | 12.33 | 1.011 | 0.166 | 0.866 | 0.063 |
| Mucosa thickness, μm | 338.85 b | 216.27 a | 190.09 a | 13.236 | <0.001 | <0.001 | 0.009 |
| Crypt depth, μm | 37.16 b | 28.34 a | 29.65 a | 2.409 | 0.043 | 0.025 | 0.106 |
| Crypt thickness, µm | 40.75 b | 30.28 a | 34.13 ab | 2.336 | 0.020 | 0.028 | 0.024 |
| Villus length, µm | 301.69 b | 187.94 a | 160.44 a | 11.507 | <0.001 | <0.001 | 0.008 |
| Villus thickness, µm | 66.25 | 59.20 | 57.90 | 2.930 | 0.129 | 0.051 | 0.436 |
| Total number of villi, /mm | 11.80 | 9.92 | 10.74 | 0.707 | 0.202 | 0.200 | 0.140 |
| Total crypt number, /mm | 12.86 | 11.34 | 11.47 | 0.899 | 0.436 | 0.243 | 0.463 |
| Villus length/crypt depth ratio | 8.55 b | 6.92 a | 5.58 a | 0.424 | <0.001 | <0.001 | 0.784 |
| Enterocyte, per 100 µm of villus | 16.99 | 16.55 | 17.39 | 0.658 | 0.671 | 0.272 | 0.797 |
| Goblet cells, per 100 µm of villus | 6.08 b | 3.50 a | 2.86 a | 0.418 | <0.001 | <0.001 | 0.077 |
| Villus epithelium thickness, µm | 16.25 b | 13.12 a | 12.23 a | 0.561 | <0.001 | <0.001 | 0.125 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomaszewska, E.; Rudyk, H.; Dobrowolski, P.; Arciszewski, M.B.; Donaldson, J.; Kras, K.; Abramowicz, B.; Kuc, D.; Muszyński, S. Basal Intestinal Morphology, Immunolocalization of Leptin and Ghrelin and Their Receptors in Newborn Wistar Rats after Prenatal Exposure to Fumonisins. Animals 2023, 13, 1538. https://doi.org/10.3390/ani13091538
Tomaszewska E, Rudyk H, Dobrowolski P, Arciszewski MB, Donaldson J, Kras K, Abramowicz B, Kuc D, Muszyński S. Basal Intestinal Morphology, Immunolocalization of Leptin and Ghrelin and Their Receptors in Newborn Wistar Rats after Prenatal Exposure to Fumonisins. Animals. 2023; 13(9):1538. https://doi.org/10.3390/ani13091538
Chicago/Turabian StyleTomaszewska, Ewa, Halyna Rudyk, Piotr Dobrowolski, Marcin B. Arciszewski, Janine Donaldson, Katarzyna Kras, Beata Abramowicz, Damian Kuc, and Siemowit Muszyński. 2023. "Basal Intestinal Morphology, Immunolocalization of Leptin and Ghrelin and Their Receptors in Newborn Wistar Rats after Prenatal Exposure to Fumonisins" Animals 13, no. 9: 1538. https://doi.org/10.3390/ani13091538
APA StyleTomaszewska, E., Rudyk, H., Dobrowolski, P., Arciszewski, M. B., Donaldson, J., Kras, K., Abramowicz, B., Kuc, D., & Muszyński, S. (2023). Basal Intestinal Morphology, Immunolocalization of Leptin and Ghrelin and Their Receptors in Newborn Wistar Rats after Prenatal Exposure to Fumonisins. Animals, 13(9), 1538. https://doi.org/10.3390/ani13091538