


Effects of a High-Concentrate Diet on the Blood Parameters and Liver Transcriptome of Goats



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals, Design, and Diet
2.2. Sample Collection
2.3. Laboratory Analyses
2.4. Transcriptome Analysis of the Liver
2.5. Validating the Expression of DEGs by RT-qPCR
2.6. Data Analysis
3. Results
3.1. The Digestibility of Nutrients
3.2. Blood Parameters
3.3. Pathological Damage of Gastrointestinal Tract and Liver Tissues
3.4. Transcriptome Analysis of Goat Liver
3.4.1. Summary of RNA and Sequencing Quality Control
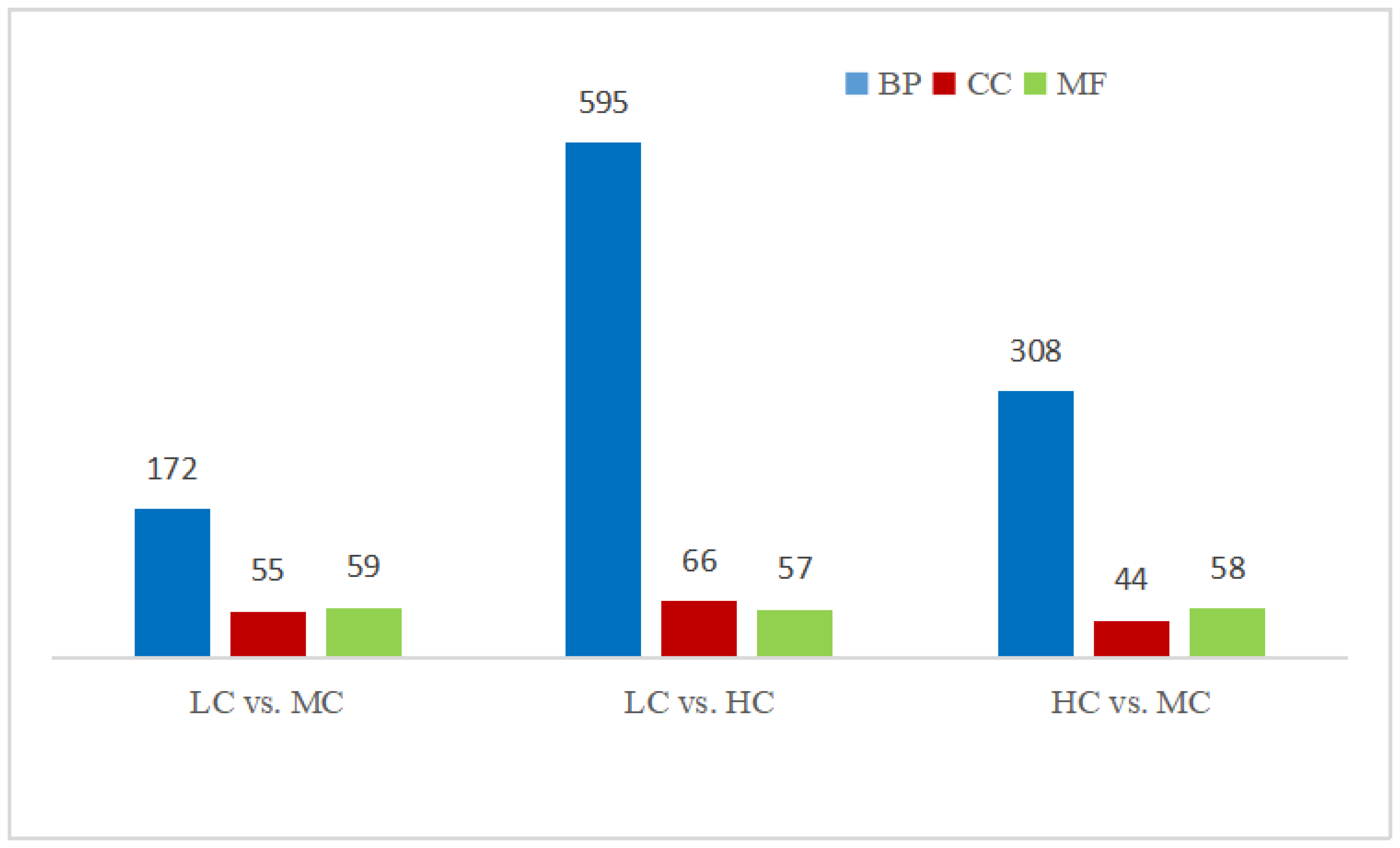
3.4.2. Differentially Expressed Genes among Groups
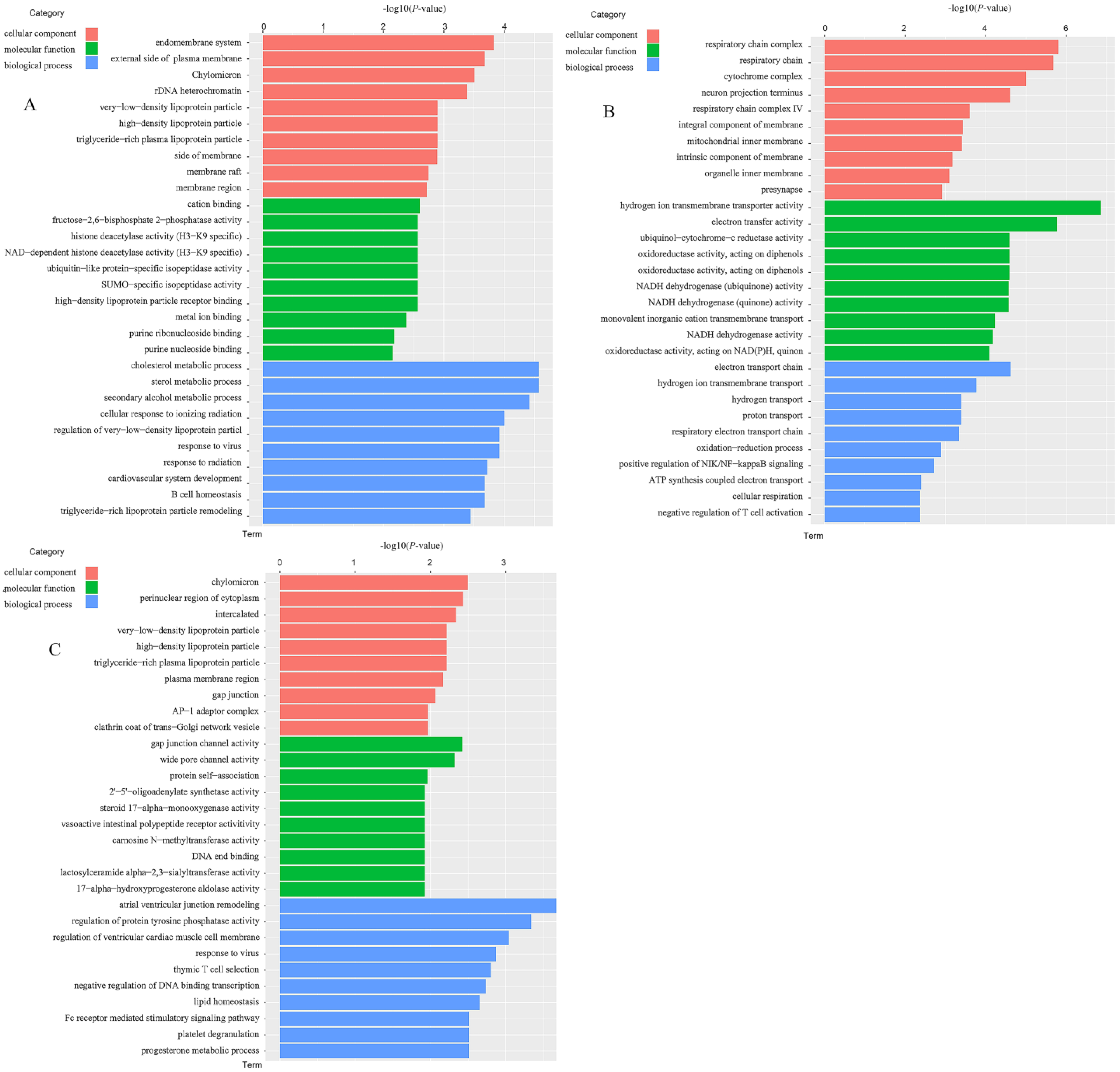
3.4.3. Analysis of GO Functional Enrichment of DEGs
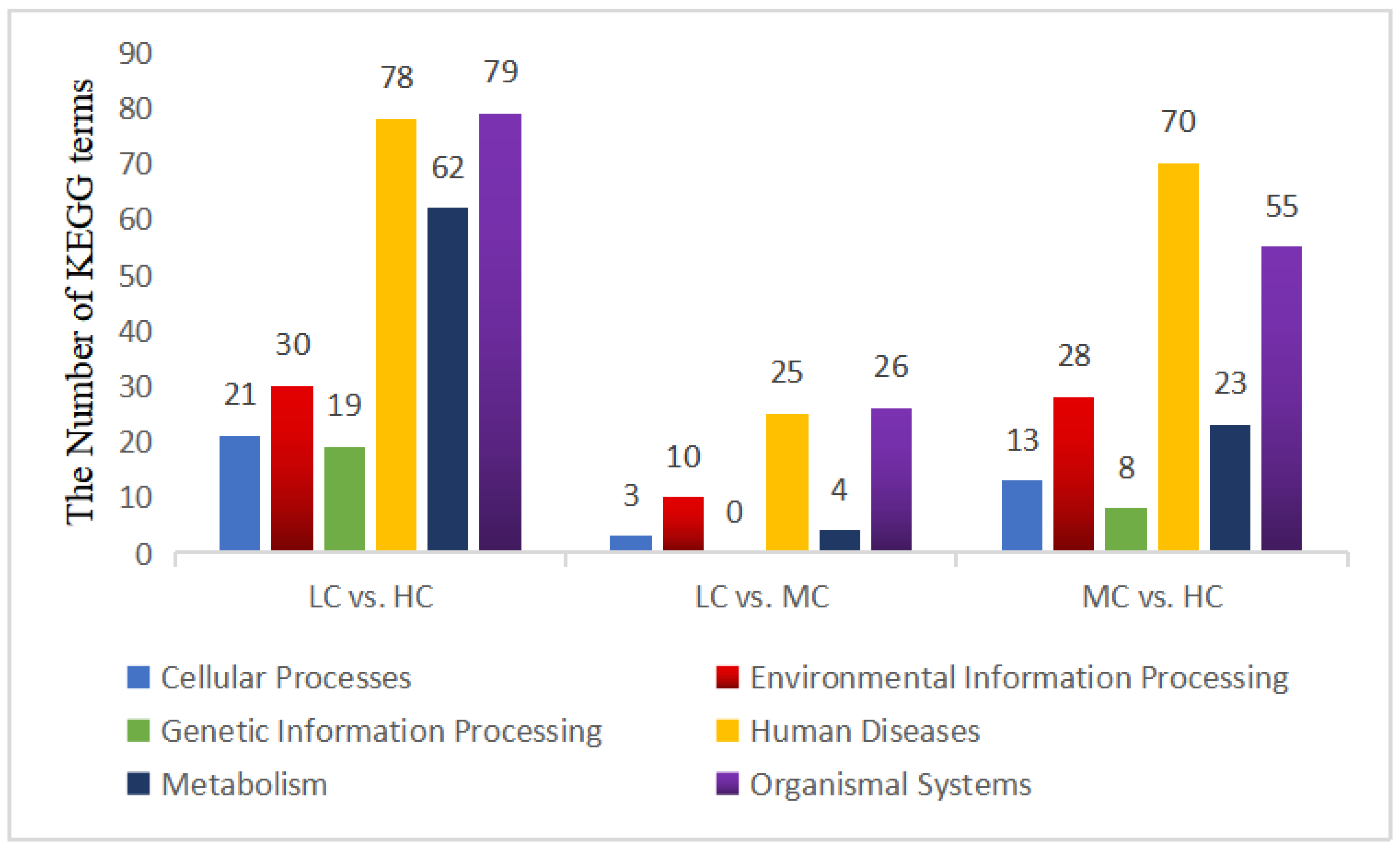
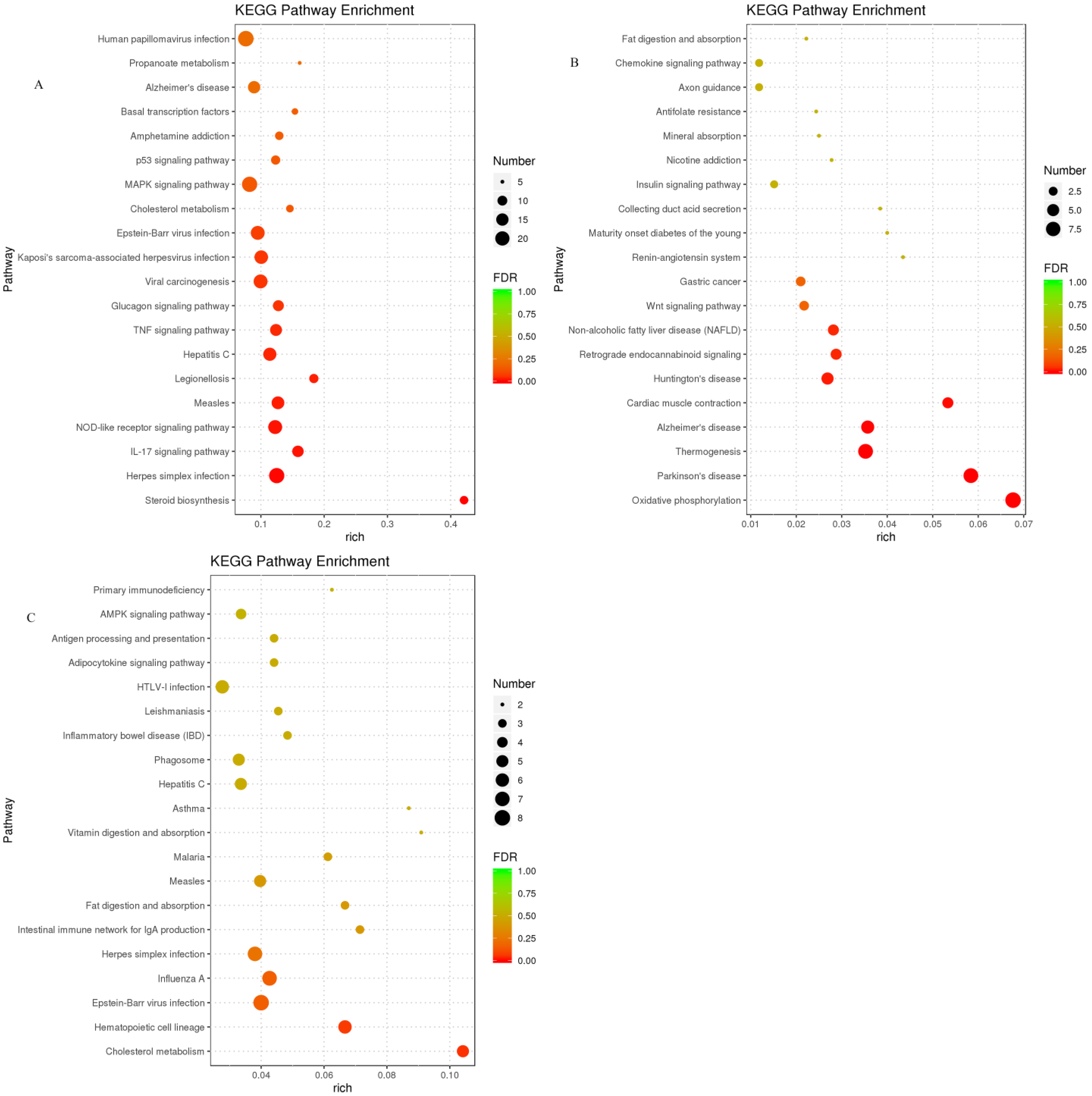
3.4.4. Analysis of the KEGG Functional Enrichment of DEGs
3.4.5. DEGs Related to Nutrient Metabolism and Toxin Detoxification
3.5. Validation of RNA-Seq Results by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cavallini, D.; Mammi, L.M.; Palmonari, A.; García-González, R.; Chapman, J.D.; McLean, D.J.; Formigoni, A. Effect of an immunomodulatory feed additive in mitigating the stress responses in lactating dairy cows to a high concentrate diet challenge. Animals 2022, 12, 2129. [Google Scholar] [CrossRef] [PubMed]
- Masebo, N.T.; Marliani, G.; Cavallini, D.; Accorsi, P.A.; Di Pietro, M.; Beltrame, A.; Gentile, A.; Jacinto, J.G.P. Health and welfare assessment of beef cattle during the adaptation period in a specialized commercial fattening unit. Res. Vet. Sci. 2023, 158, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Sanh, M.V.; Wiktorsson, H.; Ly, L.V. Effects of natural grass forage to concentrate ratios and feeding principles on milk production and performance of crossbred lactating cows. Asian-Australas. J. Anim. Sci. 2002, 15, 650–657. [Google Scholar] [CrossRef]
- Guthrie, M.J.; Galyean, M.L.; Malcolm-Callis, K.J.; Duff, G.C. Roughage source and level in beef cattle finishing diets. Prof. Anim. Sci. 1996, 12, 192–198. [Google Scholar] [CrossRef]
- Goad, D.W.; Goad, C.L.; Nagaraja, T.G. Ruminal microbial and fermentative changes associated with experimentally induced subacute acidosis in steers. J. Anim. Sci. 1998, 76, 234–241. [Google Scholar] [CrossRef]
- Clauss, M.; Hume, I.D.; Hummel, J. Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 2010, 4, 979–992. [Google Scholar] [CrossRef]
- Owens, F.N.; Secrist, D.S.; Hill, W.J.; Gill, D.R. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef]
- Drackley, J.K.; Donkin, S.S.; Reynolds, C.K. Major advances in fundamental dairy cattle nutrition. J. Dairy Sci. 2006, 89, 1324–1336. [Google Scholar] [CrossRef]
- Górka, P.; Schurmann, B.L.; Walpole, M.E.; Błońska, A.; Li, S.; Plaizier, J.C.; Kowalski, Z.M.; Penner, G.B. Effect of increasing the proportion of dietary concentrate on gastrointestinal tract measurements and brush border enzyme activity in holstein steers. J. Dairy Sci. 2017, 100, 4539–4551. [Google Scholar] [CrossRef]
- Gressley, T.F.; Hall, M.B.; Armentano, L.E. Ruminant nutrition symposium: Productivity, digestion, and health responses to hindgut acidosis in ruminants. J. Anim. Sci. 2011, 89, 1120–1130. [Google Scholar] [CrossRef]
- Ye, H.; Liu, J.; Feng, P.; Zhu, W.; Mao, S. Grain-rich diets altered the colonic fermentation and mucosa-associated bacterial communities and induced mucosal injuries in goats. Sci. Rep. 2016, 6, 20329. [Google Scholar] [CrossRef] [PubMed]
- Girolami, F.; Barbarossa, A.; Badino, P.; Ghadiri, S.; Cavallini, D.; Zaghini, A.; Nebbia, C. Effects of turmeric powder on aflatoxin m1 and aflatoxicol excretion in milk from dairy cows exposed to aflatoxin b1 at the eu maximum tolerable levels. Toxins 2022, 14, 430. [Google Scholar] [CrossRef]
- Cavallini, D.; Penazzi, L.; Valle, E.; Raspa, F.; Bergero, D.; Formigoni, A.; Fusaro, I. When changing the hay makes a difference: A series of case reports. J. Equine Vet. Sci. 2022, 113, 103940. [Google Scholar] [CrossRef] [PubMed]
- Buonaiuto, G.; Cavallini, D.; Mammi, L.M.E.; Ghiaccio, F.; Palmonari, A.; Formigoni, A.; Visentin, G. The accuracy of nirs in predicting chemical composition and fibre digestibility of hay-based total mixed rations. Ital. J. Anim. Sci. 2021, 20, 1730–1739. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official’s Analytical Chemists; Association of Official Analytical Chemists: Arlington, VA, USA, 2003. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Doyle, M.P.; Harmon, B.G.; Brown, C.A.; Mueller, P.E.; Parks, A.H. Reduction of carriage of enterohemorrhagic Escherichia coli O157:H7 in cattle by inoculation with probiotic bacteria. J. Clin. Microbiol. 1998, 36, 641–647. [Google Scholar] [CrossRef]
- Zhang, K.; Meng, M.; Gao, L.; Tu, Y.; Bai, Y. Sodium butyrate improves high-concentrate-diet-induced impairment of ruminal epithelium barrier function in goats. J. Agric. Food Chem. 2018, 66, 8729–8736. [Google Scholar] [CrossRef]
- Chevallier, M.; Guerret, S.; Chossegros, P.; Gerard, F.; Grimaud, J.A. A histological semiquantitative scoring system for evaluation of hepatic fibrosis in needle liver biopsy specimens: Comparison with morphometric studies. Hepatology 1994, 20, 349–355. [Google Scholar] [CrossRef]
- Fan, H.; Qiu, M.; Mei, J.; Shen, G.; Liu, S.; Chen, R. Effects of four regulating-intestine prescriptions on pathology and ultrastructure of colon tissue in rats with ulcerative colitis. World J. Gastroenterol. WJG 2005, 11, 4800. [Google Scholar] [CrossRef]
- Wang, W.; Li, C.; Li, F.; Wang, X.; Zhang, X.; Liu, T.; Nian, F.; Yue, X.; Li, F.; Pan, F. Effects of early feeding on the host rumen transcriptome and bacterial diversity in lambs. Sci. Rep. 2016, 6, 32479. [Google Scholar] [CrossRef]
- Pollesel, M.; Tassinari, M.; Frabetti, A.; Fornasini, D.; Cavallini, D. Effect of does parity order on litter homogeneity parameters. Ital. J. Anim. Sci. 2020, 19, 1188–1194. [Google Scholar] [CrossRef]
- Dong, G.; Liu, S.; Wu, Y.; Lei, C.; Zhou, J.; Zhang, S. Diet-induced bacterial immunogens in the gastrointestinal tract of dairy cows: Impacts on immunity and metabolism. Acta Vet. Scand. 2011, 53, 48. [Google Scholar] [CrossRef]
- Dong, H.; Wang, S.; Jia, Y.; Ni, Y.; Zhang, Y.; Zhuang, S.; Shen, X.; Zhao, R. Long-term effects of subacute ruminal acidosis (sara) on milk quality and hepatic gene expression in lactating goats fed a high-concentrate diet. PLoS ONE 2013, 8, e82850. [Google Scholar] [CrossRef]
- Monteiro, H.F.; Faciola, A.P. Ruminal acidosis, bacterial changes, and lipopolysaccharides. J. Anim. Sci. 2020, 98, a248. [Google Scholar] [CrossRef]
- Penner, G.B.; Steele, M.A.; Aschenbach, J.R.; McBride, B.W. Ruminant nutrition symposium: Molecular adaptation of ruminal epithelia to highly fermentable diets. J. Anim. Sci. 2011, 89, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zhang, J.; Li, S.; Ji, S.; Cao, Z.; Zhang, H.; Wang, Y. Effects of a wide range of dietary forage-to-concentrate ratios on nutrient utilization and hepatic transcriptional profiles in limit-fed holstein heifers. BMC Genom. 2018, 19, 148. [Google Scholar] [CrossRef] [PubMed]
- Ceciliani, F.; Lecchi, C.; Urh, C.; Sauerwein, H. Proteomics and metabolomics characterizing the pathophysiology of adaptive reactions to the metabolic challenges during the transition from late pregnancy to early lactation in dairy cows. J. Proteom. 2018, 178, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef] [PubMed]
- Klingenspor, M. Cold-induced recruitment of brown adipose tissue thermogenesis. Exp. Physiol. 2003, 88, 141–148. [Google Scholar] [CrossRef]
- Bortvedt, S.F.; Lund, P.K. Insulin-like growth factor 1: Common mediator of multiple enterotrophic hormones and growth factors. Curr. Opin. Gastroenterol. 2012, 28, 89. [Google Scholar] [CrossRef] [PubMed]
- Duanmu, Y.; Cong, R.; Tao, S.; Tian, J.; Dong, H.; Zhang, Y.; Ni, Y.; Zhao, R. Comparative proteomic analysis of the effects of high-concentrate diet on the hepatic metabolism and inflammatory response in lactating dairy goats. J. Anim. Sci. Biotechnol. 2016, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Liu, X.; Yan, J.; Aabdin, Z.U.; Bilal, M.S.; Shen, X. Sodium butyrate ameliorates high-concentrate diet-induced inflammation in the rumen epithelium of dairy goats. J. Agric. Food Chem. 2017, 65, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef]
- Emmanuel, D.; Dunn, S.M.; Ametaj, B.N. Feeding high proportions of barley grain stimulates an inflammatory response in dairy cows. J. Dairy Sci. 2008, 91, 606–614. [Google Scholar] [CrossRef]
- Andersen, P.H.; Jarløv, N.; Hesselholt, M.; Baek, L. Studies on in vivo endotoxin plasma disappearance times in cattle. J. Vet. Med. Ser. A 1996, 43, 93–101. [Google Scholar] [CrossRef]
- Zlydaszyk, J.C.; Moon, R.J. Fate of 51cr-labeled lipopolysaccharide in tissue culture cells and livers of normal mice. Infect. Immun. 1976, 14, 100–105. [Google Scholar] [CrossRef]
- Heinrich, P.C.; Castell, J.V.; Andus, T. Interleukin-6 and the acute phase response. Biochem. J. 1990, 265, 621. [Google Scholar] [CrossRef]
- Taguchi, Y.; Yamamoto, M.; Yamate, T.; Lin, S.; Mocharla, H.; DeTogni, P.; Nakayama, N.; Boyce, B.F.; Abe, E.; Manolagas, S.C. Interleukin-6-type cytokines stimulate mesenchymal progenitor differentiation toward the osteoblastic lineage. Proc. Assoc. Am. Physicians 1998, 110, 559–574. [Google Scholar]
- Tao, S.; Duanmu, Y.; Dong, H.; Tian, J.; Ni, Y.; Zhao, R. A high-concentrate diet induced colonic epithelial barrier disruption is associated with the activating of cell apoptosis in lactating goats. BMC Vet. Res. 2014, 10, 235. [Google Scholar] [CrossRef]
- Van der Vorst, E.P.C. High-density lipoproteins and apolipoprotein a1. Vertebrate and invertebrate respiratory proteins, lipoproteins and other body fluid proteins. Subcell. Biochem. 2020, 94, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Schletter, J.; Heine, H.; Ulmer, A.J.; Rietschel, E.T. Molecular mechanisms of endotoxin activity. Arch. Microbiol. 1995, 164, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S.; Hirschfeld, M.J.; Perera, P. Signal integration in lipopolysaccharide (lps)-stimulated murine macrophages. J. Endotoxin Res. 2001, 7, 237–241. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Group 2 | ||
|---|---|---|---|
| LC | MC | HC | |
| Ingredients | |||
| Corn | 4.00 | 28.67 | 49.25 |
| Wheat bran | 7.00 | 2.71 | 7.30 |
| Soybean meal | 9.00 | 6.78 | 10.50 |
| Rapeseed meal | 1.00 | 6.32 | 1.00 |
| Cottonseed meal | 1.50 | 2.26 | 8.00 |
| Oat grass | 43.80 | 24.00 | 5.00 |
| Wheat straw | 8.00 | 6.00 | 1.00 |
| Alfalfa hay | 23.00 | 20.00 | 14.00 |
| CaCO3 | 0.15 | 0.50 | 1.00 |
| CaHPO4 | 0.05 | 0.26 | 0.45 |
| NaCl | 0.50 | 0.50 | 0.50 |
| NaHCO3 | 1.00 | 1.00 | 1.00 |
| Premix 1 | 1.00 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Chemical composition | |||
| Dry matter | 88.84 | 89.05 | 90.14 |
| Crude protein | 12.06 | 13.77 | 15.93 |
| Ether extract | 3.08 | 4.23 | 5.23 |
| Crude fiber | 23.19 | 16.42 | 8.71 |
| Nitrogen free extract | 35.24 | 42.11 | 50.89 |
| Metabolizable energy, MJ/kg | 2.17 | 2.49 | 2.88 |
| Neutral detergent fiber | 43.54 | 31.65 | 19.55 |
| Acid detergent fiber | 21.05 | 15.04 | 7.34 |
| Calcium | 0.59 | 0.67 | 0.76 |
| Phosphorus | 0.37 | 0.43 | 0.49 |
| Concentrate:roughage | 25:75 | 50:50 | 80:20 |
| Item | Group 1 | ||
|---|---|---|---|
| LC | MC | HC | |
| Dry matter | 64.30 ± 2.99 b | 62.70 ± 4.84 b | 79.44 ± 3.59 a |
| Crude protein | 70.30 ± 5.94 b | 67.10 ± 5.56 b | 80.48 ± 3.80 a |
| Gross energy | 63.24 ± 2.94 b | 61.45 ± 4.69 b | 79.08 ± 3.54 a |
| Acid detergent fiber | 49.43 ± 5.19 ab | 34.74 ± 9.36 b | 51.94 ± 8.84 a |
| Neutral detergent fiber | 54.86 ± 6.15 b | 47.68 ± 8.72 b | 64.24 ± 6.12 a |
| Ether extract | 59.59 ± 3.60 b | 58.73 ± 4.82 b | 79.65 ± 3.33 a |
| Item | Group 1 | ||
|---|---|---|---|
| LC | MC | HC | |
| Glucose, g/L | 6.70 ± 1.66 a | 5.04 ± 0.50 b | 5.36 ± 0.55 b |
| Triglyceride, g/L | 0.69 ± 0.08 | 0.69 ± 0.08 | 0.67 ± 0.09 |
| Non-esterified fatty acid, mmol/L | 0.11 ± 0.03 | 0.09 ± 0.03 | 0.12 ± 0.01 |
| Lipopolysaccharide, EU/L | 203.65 ± 31.76 b | 193.43 ± 10.41 b | 247.07 ± 27.70 a |
| Lactic acid, μg/L | 496.67 ± 422.93 b | 841.25 ± 668.99 b | 2093.75 ± 759.75 a |
| Acetate, μmol/L | 701.66 ± 146.24 b | 918.13 ± 119.80 b | 1227.25 ± 313.51 a |
| Propionic, μmol/L | 72.95 ± 10.54 b | 95.92 ± 10.55 b | 114.33 ± 12.52 a |
| Butyrate, μmol/L | 46.37 ± 6.21 b | 52.38 ± 6.58 b | 65.68 ± 15.53 a |
| Item | Group 1 | ||
|---|---|---|---|
| LC | MC | HC | |
| Glucose, g/L | 3.33 ± 0.75 | 3.08 ± 0.97 | 3.73 ± 1.08 |
| Acetate, μmol/L | 60.68 ± 18.50 | 46.77 ± 16.72 | 47.91 ± 10.42 |
| Propionic, μmol/L | 10.73 ± 2.12 | 9.20 ± 1.25 | 9.36 ± 2.45 |
| Butyrate, μmol/L | 3.91 ± 2.82 | 5.13 ± 1.91 | 6.09 ± 1.21 |
| Alanine transaminase, g/L | 24.33 ± 10.07 | 18.17 ± 5.49 | 23.50 ± 7.15 |
| Blood urea nitrogen, g/L | 7.32 ± 3.08 | 6.60 ± 1.40 | 8.58 ± 1.87 |
| Triglyceride, g/L | 0.57 ± 0.09 | 0.51 ± 0.06 | 0.50 ± 0.04 |
| Total protein, g/L | 70.92 ± 6.22 | 73.48 ± 9.31 | 65.37 ± 5.58 |
| Albumin, g/L | 34.32 ± 2.56 | 35.52 ± 3.50 | 34.57 ± 1.56 |
| Low-density lipoprotein-C, mmol/L | 0.34 ± 0.13 | 0.25 ± 0.10 | 0.25 ± 0.10 |
| High-density lipoprotein-C, mmol/L | 1.04 ± 0.21 | 1.21 ± 0.34 | 1.16 ± 0.30 |
| Lipopolysaccharide, EU/L | 202.71 ± 10.02 | 189.01 ± 13.09 | 201.09 ± 21.91 |
| Lactic acid, μg/L | 2.94 ± 0.67 | 4.38 ± 0.21 | 2.67 ± 0.12 |
| Interleukin-1β, ng/L | 234.44 ± 24.94 b | 247.71 ± 28.03 b | 288.29 ± 62.02 a |
| Interleukin-6, ng/L | 104.72 ± 40.93 | 128.19 ± 40.18 | 115.64 ± 19.25 |
| Tumor Necrosis Factor-α, ng/L | 1757.60 ± 230.23 | 1758.60 ± 531.31 | 1796.74 ± 116.66 |
| Interleukin-4, ng/L | 94.15 ± 23.25 | 127.74 ± 69.16 | 155.48 ± 36.20 |
| Interleukin-10, ng/L | 1279.65 ± 186.63 b | 2217.24 ± 767.27 a | 2319.94 ± 423.92 a |
| Diamineoxidase, pg/ml | 204.89 ± 18.52 b | 210.38 ± 15.88 b | 273.99 ± 11.94 a |
| Lipopolysaccharide-binding protein, pg/mL | 10.07 ± 0.42 | 9.63 ± 0.47 | 10.09 ± 0.51 |
| Haptoglobin, pg/mL | 14.60 ± 1.04 b | 16.93 ± 0.60 b | 18.53 ± 0.85 a |
| Serum amyloid A protein, pg/mL | 439.20 ± 13.46 b | 530.35 ± 31.67 a | 548.79 ± 22.35 a |
| Tissue 2 | Group 1 | ||
|---|---|---|---|
| LC | MC | HC | |
| Liver | 1.33 ± 0.52 | 1.83 ± 0.75 | 1.50 ± 0.84 |
| Jejunum | 1.00 ± 0.63 b | 0.83 ± 0.75 b | 2.17 ± 0.98 a |
| Colon | 1.80 ± 0.45 | 2.00 ± 0.00 | 1.80 ± 0.45 |
| Rumen | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Pathway ID | Pathway Name | Groups 1 | |||||
|---|---|---|---|---|---|---|---|
| LC vs. HC | LC vs. MC | MC vs. HC | |||||
| Up-Regulated Gene | Down-Regulated Gene | Up-Regulated Gene | Down-Regulated Gene | Up-Regulated Gene | Down-Regulated Gene | ||
| chx04152 | AMPK signaling pathway | CCNA1, CREB3L4, CPT1 | CREB3L1, PFKFB1, LOC108637886, PRKAB1, HMGCR, MAP3K7, SIRT1, ACACA | CCNA1 | PFKFB1, LOC108633303, ACACA | ||
| chx03320 | PPAR signaling pathway | EHHADH | APOA5, CPT1B, APOA1, APOA2, APOC3 | SORBS1 | LOC108633303 | APOA1, APOC3 | |
| chx04910 | Insulin signaling pathway | PRKAR1B, CALML4 | SOCS3, SOCS2, SOS1, PRKAB1, ACACA, MAPK8, PPP1R3C, PPP1CB, EIF4E, PRKACA | SHC2, SORBS1 | SOCS3, ACACA, SHC2, EIF4E | ||
| chx04714 | Thermogenesis signaling pathways | CREB3L1, KLB, SOS1, PRKAB1, SIRT6, PRKACA | ATP6, COX7A1, CREB3L4, SLC25A29, ND6, CPT1B, COX2, LOC102173562, ATP5I, NDUFB9, LOC102168533 | ND1, ATP6, ND5, LOC102168533, CYTB, ND6, COX3, COX2 | COX7A1 | ||
| chx04064 | NF-κB signaling pathway | TRAF3 | CHUK, LYN, TAB2, MAP3K7, NFRSF13C, ICAM1, IL1R1 | TLR4, LYN, TRAF5 | |||
| chx04620 | Toll-like receptor signaling pathway | IRF7, TRAF3 | TLR4, TLR7, CHUK, TAB2, MAP3K7, MAPK8, IFNAR2, FADD | TLR7 | IRF7 | TLR4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Q.; Wang, L.; Liu, Y.; Yan, T. Effects of a High-Concentrate Diet on the Blood Parameters and Liver Transcriptome of Goats. Animals 2023, 13, 1559. https://doi.org/10.3390/ani13091559
Wang Y, Li Q, Wang L, Liu Y, Yan T. Effects of a High-Concentrate Diet on the Blood Parameters and Liver Transcriptome of Goats. Animals. 2023; 13(9):1559. https://doi.org/10.3390/ani13091559
Chicago/Turabian StyleWang, Yusu, Qiong Li, Lizhi Wang, Yuehui Liu, and Tianhai Yan. 2023. "Effects of a High-Concentrate Diet on the Blood Parameters and Liver Transcriptome of Goats" Animals 13, no. 9: 1559. https://doi.org/10.3390/ani13091559
APA StyleWang, Y., Li, Q., Wang, L., Liu, Y., & Yan, T. (2023). Effects of a High-Concentrate Diet on the Blood Parameters and Liver Transcriptome of Goats. Animals, 13(9), 1559. https://doi.org/10.3390/ani13091559