
Temperament Behaviours in Individually Tested Sheep Are Not Related to Behaviours Expressed in the Presence of Conspecifics


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Housing
2.3. Behavioural Testing
2.3.1. Stationary Human (H) Test
2.3.2. Startle (S)/Novel Object Test
2.3.3. Isolation Box (IB)/Temperament Test
2.4. Statistical Analysis
2.4.1. Determining the Repeatability of Behaviours across Social Conditions
2.4.2. Exploring the Relationship between Behaviours across Social Situations
3. Results
3.1. Repeatability of Behaviours across Social Conditions
3.2. Exploring the Relationship between Behaviours across Social Situations by Comparing Rank Order
3.3. Exploring the Relationship between Behaviours across Social Situations by Comparing Behavioural Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dodd, C.L.; Pitchford, W.S.; Edwards, J.E.H.; Hazel, S.J. Measures of behavioural reactivity and their relationships with production traits in sheep: A review. Appl. Anim. Behav. Sci. 2012, 140, 1–15. [Google Scholar] [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Bell, A.M.; Johnson, J.C.; Ziemba, R.E. Behavioral syndromes: An integrative overview. Q. Rev. Biol. 2004, 79, 241–277. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.M. Maternal Behaviour and Rearing Ability of Merino Ewes Can Be Improved by Strategic Feed Supplementation during Late Pregnancy and Selection for Calm Temperament. Ph.D. Thesis, University of Western Australia, Perth, Australia, 1999. [Google Scholar]
- Putu, I.I.G. Maternal Behaviour in Merino Ewes during the First Two Days after Parturition and Survival of Lambs. Ph.D. Thesis, University of Western Australia, Perth, Australia, 1988. [Google Scholar]
- Schiller, K.; McVey, C.; Doyle, S.; Horback, K. Chute scoring as a potential method for assessing individual differences in arousal among ewes. Appl. Anim. Behav. Sci. 2020, 230, 105073. [Google Scholar] [CrossRef]
- Doyle, R.E.; Broster, J.C.; Barnes, K.; Browne, W.J. Temperament, age and weather predict social interaction in the sheep flock. Behav. Process. 2016, 131, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Kilgour, R.; Szantar-Coddington, M. Arena behaviour of ewes selected for superior mothering ability differs from that of unselected ewes. Anim. Reprod. Sci. 1995, 37, 133–141. [Google Scholar] [CrossRef]
- Çakmakçı, C. Estimating the repeatability of behavioral traits in Norduz sheep subjected to an arena test. Appl. Anim. Behav. Sci. 2022, 254, 105704. [Google Scholar] [CrossRef]
- Atkinson, L.; Doyle, R.E.; Woodward, A.; Jongman, E.C. Behavioural reactivity testing in sheep indicates the presence of multiple temperament traits. Behav. Process. 2022, 201, 104711. [Google Scholar] [CrossRef]
- Cockram, M.S.; Ranson, M.; Imlah, P.; Goddard, P.J.; Burrells, C.; Harkiss, G.D. The behavioural, endocrine and immune responses of sheep to isolation. Anim. Prod. 1994, 58, 389–399. [Google Scholar] [CrossRef]
- Rushen, J. Use of aversion-learning techniques to measure distress in sheep. Appl. Anim. Behav. Sci. 1990, 28, 3–14. [Google Scholar] [CrossRef]
- González, M.; Averós, X.; de Heredia, I.B.; Ruiz, R.; Arranz, J.; Estevez, I. The effect of social buffering on fear responses in sheep (Ovis aries). Appl. Anim. Behav. Sci. 2013, 149, 13–20. [Google Scholar] [CrossRef]
- Kilgour, R.J.; Szantar-Coddington, M.R. The arena test and cortisol response of sheep as indirect selection criteria for the improvement of lamb survival. Anim. Reprod. Sci. 1997, 46, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.C.; Canali, E.; Jones, R.B. A critical review of fear tests used on cattle, pigs, sheep, poultry and horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef] [PubMed]
- Vandenheede, M.; Bouissou, M. Sex differences in fear reactions in sheep. Appl. Anim. Behav. Sci. 1993, 37, 39–55. [Google Scholar] [CrossRef]
- Boissy, A.; Bouix, J.; Orgeur, P.; Poindron, P.; Bibé, B.; Le Neindre, P. Genetic analysis of emotional reactivity in sheep: Effects of the genotypes of the lambs and of their dams. Genet. Sel. Evol. 2005, 37, 381–401. [Google Scholar] [CrossRef] [PubMed]
- Viérin, M.; Bouissou, M.F. Responses of weaned lambs to fear-eliciting situations: Origin of individual differences. Dev. Psychobiol. J. Int. Soc. Dev. Psychobiol. 2003, 42, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Rice, M.; Borg, S.; Edwards, L.E.; Ponnampalam, E.N.; Coleman, G.J. Relationships between handling, behaviour and stress in lambs at abattoirs. Animal 2019, 13, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Christensen, R. Ordinal—Regression Models for Ordinal Data (R Package Version 2019.12-10); CRAN Team: Vienna, Austria, 2019. [Google Scholar]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research (R Package Version 2.1.9); Northwestern University: Evanston, IL, USA, 2021. [Google Scholar]
- Tabachnick, B.; Fidell, L. Using Multivariate Statistics, 5th ed.; Pearson Education: Boston, MA, USA, 2007. [Google Scholar]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions (R Package Version 2.1.2); CRAN Team: Vienna, Austria, 2021. [Google Scholar]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R package for determining the relevant number of clusters in a data set. J. Stat. Softw. 2014, 61, 1–36. [Google Scholar] [CrossRef]
- Marini, D.; Monk, J.E.; Campbell, D.L.M.; Lee, C.; Belson, S.; Small, A. Sex impacts pain behaviour but not emotional reactivity of lambs following ring tail docking. PeerJ 2023, 11, e15092. [Google Scholar] [CrossRef]
- Price, E.O.; Thos, J. Behavioral responses to short-term social isolation in sheep and goats. Appl. Anim. Ethol. 1980, 6, 331–339. [Google Scholar] [CrossRef]
- Murphy, P.M.; Purvis, I.W.; Lindsay, D.R.; Leneindre, P.; Orgeur, P.; Poindron, P. Measures of temperament are highly repeatable in merino sheep and some are related to maternal behaviour. Proc. Aust. Soc. Anim. Prod. 1994, 20, 247–248. [Google Scholar]
- Monk, J.E.; Colditz, I.G.; Clark, S.; Lee, C. Repeatability of an attention bias test for sheep suggests variable influence of state and trait affect on behaviour. PeerJ 2023, 11, e14730. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, G. What can vigilance tell us about fear? Anim. Sentience 2017, 2, 1. [Google Scholar] [CrossRef]
- Beausoleil, N.J.; Blache, D.; Stafford, K.J.; Mellor, D.J.; Noble, A.D.L. Selection for temperament in sheep: Domain-general and context-specific traits. Appl. Anim. Behav. Sci. 2012, 139, 74–85. [Google Scholar] [CrossRef]
- Koolhaas, J.; Van Reenen, C. Animal behavior and well-being symposium: Interaction between coping style/personality, stress, and welfare: Relevance for domestic farm animals. J. Anim. Sci. 2016, 94, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Rice, M.; Karlen, M.G.; Calleja, L.; Barnett, J.L.; Nash, J.; Coleman, G.J. Human–animal interactions at abattoirs: Relationships between handling and animal stress in sheep and cattle. Appl. Anim. Behav. Sci. 2011, 135, 24–33. [Google Scholar] [CrossRef]
- Benus, R.; Bohus, B.; Koolhaas, J.; Van Oortmerssen, G. Heritable variation for aggression as a reflection of individual coping strategies. Experientia 1991, 47, 1008–1019. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Volchan, E.; Rocha-Rego, V.; Bastos, A.F.; Oliveira, J.M.; Franklin, C.; Gleiser, S.; Berger, W.; Souza, G.G.L.; Oliveira, L.; David, I.A.; et al. Immobility reactions under threat: A contribution to human defensive cascade and PTSD. Neurosci. Biobehav. Rev. 2017, 76, 29–38. [Google Scholar] [CrossRef]
- Kozlowska, K.; Walker, P.; McLean, L.; Carrive, P. Fear and the defense cascade: Clinical implications and management. Harv. Rev. Psychiatry 2015, 23, 263. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Davis, M.; Ohman, A. Fear and anxiety: Animal models and human cognitive psychophysiology. J. Affect. Disord. 2000, 61, 137–159. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviour | Description | H Test | S Test | IB Test |
|---|---|---|---|---|
| Vocalisations | Number of vocalisations (low- and high-pitched bleats). | IND GRP | IND GRP | IND |
| Escape attempts | Number of jumps against the arena or isolation box walls (two feet or less remain on the ground). | IND GRP | IND GRP | IND |
| Locomotion | Number of single front-foot steps. | IND GRP | IND GRP | IND |
| Vigilance | Time spent with head at or above shoulder height. | IND GRP | IND GRP | |
| Attention to stimulus | Time spent with head oriented towards the stimulus (human/umbrella/ball). | IND GRP | IND GRP | |
| Proximity to stimulus | Closest zone to stimulus the lamb entered: 1 = came within 0–1 m 2 = came within 1–2 m 3 = came within 2–3 m 4 = stayed 3 m + away | IND GRP | IND GRP | |
| Interactions with the stimulus | If contact was made with the stimulus (human/umbrella/ball) using any body part (yes/no). | IND | IND | |
| Startle response | Magnitude of startle response: 0 = did not startle 1 = jumped/startled but took no steps 2 = took steps but moved <1 square 3 = ran or moved >1 square 4 = fled and may have attempted escape | IND GRP | ||
| Turns | Number of times lamb turned body 180 degrees to face the opposite door of the isolation box | IND |
| Test | Behaviour | Unit | Number of Lambs to Perform Behaviour | Mean per Individual | Range (min, max) | |||
|---|---|---|---|---|---|---|---|---|
| IND | GRP | IND | GRP | IND | GRP | |||
| Human | Vocalisations | count | 179 (81%) | 7 (3%) | 18.5 | 0.2 | (0, 75) | (0, 13) |
| Escape Attempts | count | 112 (51%) | 4 (2%) | 1.4 | 0.02 | (0, 19) | (0, 1) | |
| Locomotion | count | 218 (99%) | 215 (98%) | 117 | 34.6 | (0, 381) | (1, 101) | |
| Vigilance | seconds | 220 (100%) | 220 (100%) | 223 | 170 | (150.8, 240) | (1.1, 240) | |
| Attention to Human | seconds | 220 (100%) | 215 (98%) | 86.4 | 31.9 | (18.2, 181.5) | (0, 156.2) | |
| Proximity to Human | 1–4 | 220 (100%) | 220 (100%) | 2.3 | 3.6 | (1, 4) | (2, 4) | |
| Startle | Vocalisations | count | 170 (77%) | 8 (4%) | 18.7 | 0.2 | (0, 87) | (0, 16) |
| Escape Attempts | count | 74 (34%) | 7 (3%) | 0.8 | 0.03 | (0, 9) | (0, 1) | |
| Locomotion | count | 220 (100%) | 217 (99%) | 112 | 32.3 | (4, 359) | (0, 94) | |
| Vigilance | seconds | 220 (100%) | 220 (100%) | 216 | 176 | (124.8, 240) | (6.3, 240) | |
| Attention to Object | seconds | 220 (100%) | 209 (95%) | 63.4 | 32.8 | (7.8, 175.2) | (0, 180.1) | |
| Proximity to Object | 1–4 | 220 (100%) | 220 (100%) | 2.1 | 3.3 | (1, 4) | (1, 4) | |
| Startle Response | 0–4 | 211 (96%) | 157 (71%) | 2.4 | 1.6 | (0, 4) | (0, 4) | |
| Test | Behaviour | IND Condition | Test Day Order | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Estimate | p-Value | 2.5% CI | 97.5% CI | Estimate | p-Value | 2.5% CI | 97.5% CI | ||
| Human | Locomotion * | 0.05 | 0.288 | −0.05 | 0.15 | - | - | - | - |
| Vigilance | 0.31 | <0.001 | 0.2 | 0.42 | 44.6 | <0.001 | 24.49 | 64.64 | |
| Attention to Human | 0.2 | <0.001 | 0.09 | 0.31 | 52.7 | <0.001 | 31.34 | 74.13 | |
| Proximity to Human | 0.05 | 0.152 | −0.02 | 0.12 | - | - | - | - | |
| Startle | Locomotion | 0.1 | 0.048 | 0.002 | 0.2 | - | - | - | - |
| Vigilance | 0.13 | 0.018 | 0.03 | 0.24 | 57.2 | <0.001 | 36.71 | 77.63 | |
| Attention to Object | 0.12 | 0.065 | −0.01 | 0.24 | 46.4 | <0.001 | 25.61 | 67.1 | |
| Proximity to Object | 0.01 | 0.639 | −0.03 | 0.05 | 1.4 | <0.001 | 1.02 | 1.69 | |
| Startle Response | 0.05 | 0.736 | −0.23 | 0.33 | 4.2 | <0.001 | 2.3 | 6.08 | |
| GRP Behaviour | IND Behaviour/Fixed Effect | Estimate | p-Value | 2.5% CI | 97.5% CI |
|---|---|---|---|---|---|
| Vigilance (H) | Vocalisations (S) * | −0.16 | 0.01 | −0.27 | −0.05 |
| Vigilance (S) | 0.14 | 0.03 | 0.02 | 0.26 | |
| Test Day Order | 48.19 | <0.001 | 28.36 | 68.05 | |
| Attention to human | Vocalisations (S) * | −0.12 | 0.04 | −0.23 | −0.01 |
| Escape Attempts (H) * | 0.16 | 0.01 | 0.04 | 0.28 | |
| Test Day Order | 59.61 | <0.001 | 38.51 | 80.66 | |
| Proximity to human | Interactions (H) | −0.002 | 0.03 | −0.003 | −0.0001 |
| IND S Steps | −0.001 | 0.01 | −0.002 | −0.0003 | |
| Vigilance (S) | Vigilance (H) | 0.2 | <0.001 | 0.09 | 0.3 |
| Test Day Order | 55.81 | <0.001 | 35.35 | 76.25 | |
| Attention to S object | Locomotion (IB) * | 0.16 | 0.01 | 0.04 | 0.29 |
| Test Day Order | 39.98 | <0.001 | 20.9 | 59.07 | |
| Sex | −19.77 | 0.03 | −36.73 | −2.79 | |
| Weight at weaning | 2.29 | 0.02 | 0.33 | 4.3 | |
| Proximity to S object | Test Day Order | 1.35 | <0.001 | 1.02 | 1.69 |
| Locomotion | Proximity to S Object | 8.11 | 0.04 | 0.59 | 15.63 |
| S response | Locomotion (H) * | −0.01 | 0.04 | −0.01 | −0.0004 |
| Test Day Order | 4.14 | <0.001 | 2.57 | 5.71 |
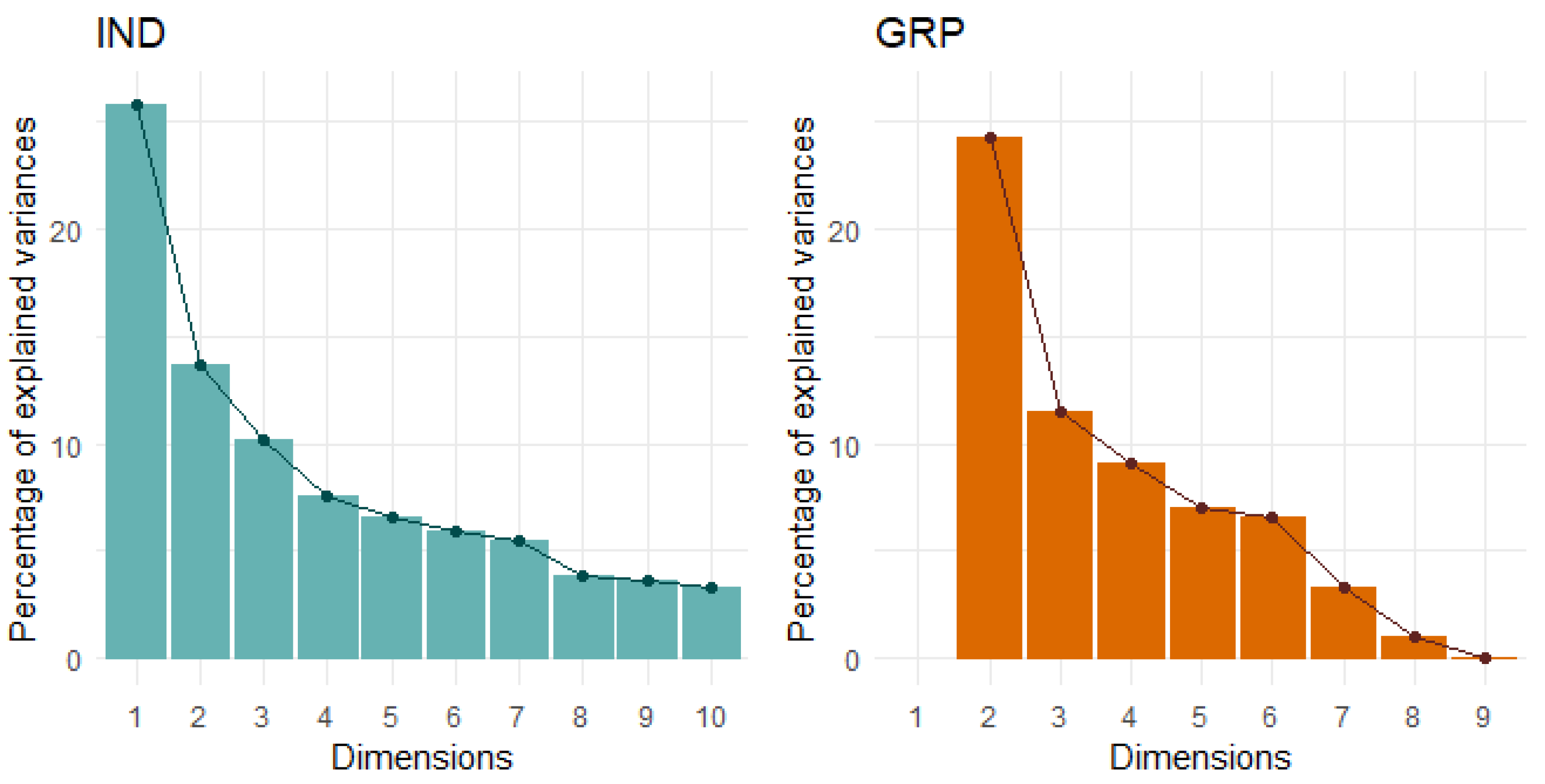
| IND Behaviours | PC1 Sociability/Explore–Avoid | PC2 General Activity | PC3 Bold–Shy | PC4 Vigilance | h2 |
|---|---|---|---|---|---|
| Vocalisations (S) | 0.85 | −0.09 | 0.07 | 0.04 | 0.75 |
| Vocalisations (H) | 0.84 | −0.18 | 0.11 | 0.07 | 0.75 |
| Proximity to Human | −0.74 | −0.12 | −0.08 | 0.18 | 0.6 |
| Interactions (H) | 0.71 | 0.05 | 0.04 | 0.04 | 0.5 |
| Interactions (S) | 0.61 | 0.41 | −0.03 | −0.22 | 0.58 |
| Proximity to S Object | −0.6 | −0.35 | −0.07 | 0.38 | 0.63 |
| Vocalisations (IB) | 0.6 | −0.09 | −0.02 | 0.29 | 0.45 |
| Locomotion (S) | 0.49 | 0.42 | 0.38 | −0.22 | 0.6 |
| Locomotion (IB) | 0.09 | 0.79 | 0.13 | 0.1 | 0.66 |
| Turns (IB) | −0.03 | 0.79 | 0.05 | 0.04 | 0.63 |
| Escape Attempts (IB) | −0.17 | 0.36 | 0.11 | 0.32 | 0.27 |
| Escape Attempts (S) | −0.02 | 0.15 | 0.76 | 0.17 | 0.63 |
| Escape Attempts (H) | −0.2 | 0.2 | 0.68 | 0.3 | 0.63 |
| Locomotion (H) | 0.38 | 0.39 | 0.58 | −0.04 | 0.64 |
| Attention to Human | −0.21 | 0.08 | −0.58 | 0.35 | 0.5 |
| Attention to S Object | −0.14 | 0.06 | −0.43 | 0.32 | 0.31 |
| Vigilance (S) | 0.07 | 0 | 0 | 0.77 | 0.6 |
| Vigilance (H) | 0.06 | 0.09 | 0.01 | 0.74 | 0.55 |
| Proportion Variance | 0.23 | 0.12 | 0.12 | 0.11 | |
| Cumulative Variance | 0.23 | 0.35 | 0.46 | 0.57 |
| IND Behaviour | Cluster IND A: Exploratory | Cluster IND B: Head Down | Cluster IND C: Active | Cluster IND D: Freeze | |
|---|---|---|---|---|---|
| PC1 | Vocalisations (S) | 1.33 | 0.16 | −0.11 | −0.53 |
| Vocalisations (H) | 1.36 | 0.13 | −0.30 | −0.49 | |
| Proximity to Human | −1.38 | −0.28 | 0.14 | 0.63 | |
| Interactions (H) | 2.23 | −0.41 | −0.21 | −0.39 | |
| Interactions (S) | 1.00 | 0.47 | 0.09 | −0.68 | |
| Proximity to S Object | −0.72 | −0.67 | −0.24 | 0.76 | |
| Vocalisations (IB) | 1.06 | −0.15 | −0.15 | −0.21 | |
| Locomotion (S) | 0.67 | 0.54 | 0.91 | −0.74 | |
| PC2 | Locomotion (IB) | 0.37 | 0.18 | 0.92 | −0.38 |
| Turns (IB) | 0.02 | 0.18 | 0.76 | −0.25 | |
| Escape Attempts (IB) | 0.05 | −0.21 | 0.68 | 0.04 | |
| PC3 | Escape Attempts (S) | −0.02 | −0.17 | 2.88 | −0.28 |
| Escape Attempts (H) | −0.20 | −0.25 | 2.54 | −0.11 | |
| Locomotion (H) | 0.67 | 0.26 | 1.37 | −0.60 | |
| Attention to Human | −0.36 | −0.28 | −0.34 | 0.37 | |
| Attention to S Object | −0.22 | −0.17 | −0.41 | 0.26 | |
| PC4 | Vigilance (S) | 0.10 | −0.30 | 0.50 | 0.12 |
| Vigilance (H) | 0.01 | −0.24 | 0.53 | 0.10 | |
| Males | 16 | 30 | 8 | 52 | |
| Females | 16 | 43 | 6 | 47 | |
| Total | 32 | 73 | 14 | 99 |
| GRP Behaviours | PC1 Attention/ Vigilance | PC2 Bold–Shy/ Activity | PC3 Response to S Object | h2 |
|---|---|---|---|---|
| Vigilance (H) | 0.77 | 0.2 | 0.19 | 0.67 |
| Attention to Human | 0.75 | −0.11 | 0.06 | 0.58 |
| Vigilance (S) | 0.73 | 0.18 | 0.28 | 0.64 |
| Attention to S Object | 0.66 | −0.02 | 0.05 | 0.44 |
| Locomotion (H) | 0.04 | 0.84 | 0.03 | 0.71 |
| Proximity to Human | 0.11 | −0.75 | 0.18 | 0.61 |
| Locomotion (S) | 0.18 | 0.6 | 0.11 | 0.41 |
| Startle Response (S) | 0.09 | 0.01 | 0.85 | 0.72 |
| Proximity to S Object | 0.22 | −0.03 | 0.71 | 0.55 |
| Proportion Variance | 0.25 | 0.19 | 0.15 | |
| Cumulative Variance | 0.25 | 0.44 | 0.96 |
| GRP Behaviour | Cluster GRP A: Attentive/Avoiding | Cluster GRP B: Active H approaching | Cluster GRP C: Wary/Inactive | Cluster GRP D: Hiding | |
|---|---|---|---|---|---|
| PC1 | Vigilance (H) | 0.68 | 0.49 | 0.37 | −1.27 |
| Attention to Human | 1.58 | −0.03 | −0.16 | −0.62 | |
| Vigilance (S) | 0.57 | 0.56 | 0.43 | −1.35 | |
| Attention to S Object | 1.35 | 0.17 | −0.24 | −0.56 | |
| PC2 | Locomotion (H) | −0.21 | 1.09 | −0.46 | −0.21 |
| Proximity to Human | 0.61 | −1.06 | 0.42 | 0.02 | |
| Locomotion (S) | 0.52 | 0.61 | −0.38 | −0.31 | |
| PC3 | Startle Response (S) | 0.61 | 0.01 | 0.12 | −0.50 |
| Proximity to S Object | 0.61 | 0.24 | 0.05 | −0.60 | |
| Males | 18 | 31 | 37 | 26 | |
| Females | 14 | 19 | 40 | 33 | |
| Total | 32 | 50 | 77 | 59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atkinson, L.; Doyle, R.E.; Jongman, E.C. Temperament Behaviours in Individually Tested Sheep Are Not Related to Behaviours Expressed in the Presence of Conspecifics. Animals 2024, 14, 155. https://doi.org/10.3390/ani14010155
Atkinson L, Doyle RE, Jongman EC. Temperament Behaviours in Individually Tested Sheep Are Not Related to Behaviours Expressed in the Presence of Conspecifics. Animals. 2024; 14(1):155. https://doi.org/10.3390/ani14010155
Chicago/Turabian StyleAtkinson, Leigh, Rebecca E. Doyle, and Ellen C. Jongman. 2024. "Temperament Behaviours in Individually Tested Sheep Are Not Related to Behaviours Expressed in the Presence of Conspecifics" Animals 14, no. 1: 155. https://doi.org/10.3390/ani14010155
APA StyleAtkinson, L., Doyle, R. E., & Jongman, E. C. (2024). Temperament Behaviours in Individually Tested Sheep Are Not Related to Behaviours Expressed in the Presence of Conspecifics. Animals, 14(1), 155. https://doi.org/10.3390/ani14010155