
Analysis of Rumen Degradation Characteristics, Attached Microbial Community, and Cellulase Activity Changes of Garlic Skin and Artemisia argyi Stalk

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Chemical Composition
2.2. Animals Feeding
2.3. Ruminal Degradability
2.4. Rumen Microbiome Analysis
2.5. Cellulase Activity Assay
2.6. Statistical Analysis Method
3. Results
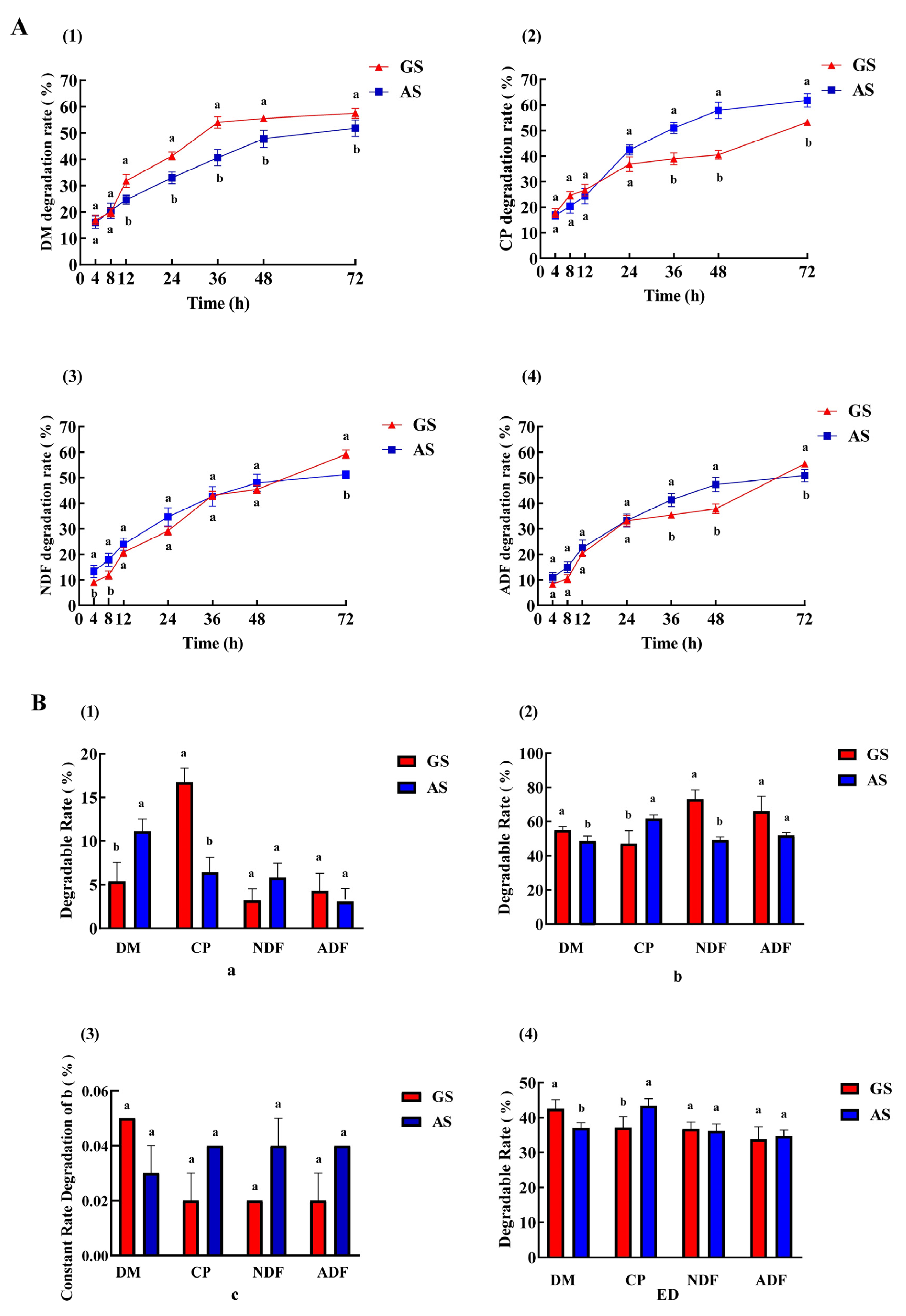
3.1. Ruminal Degradability
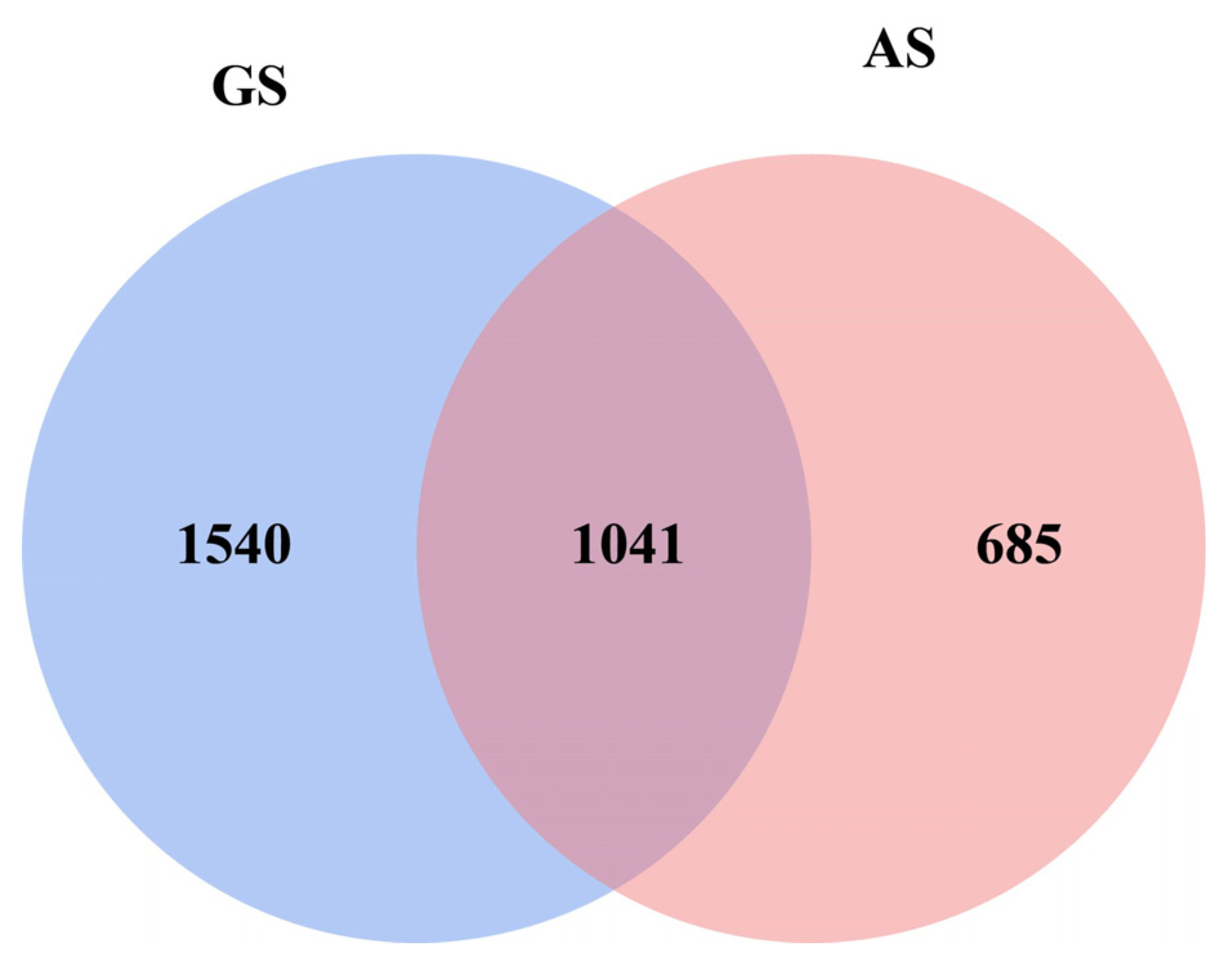
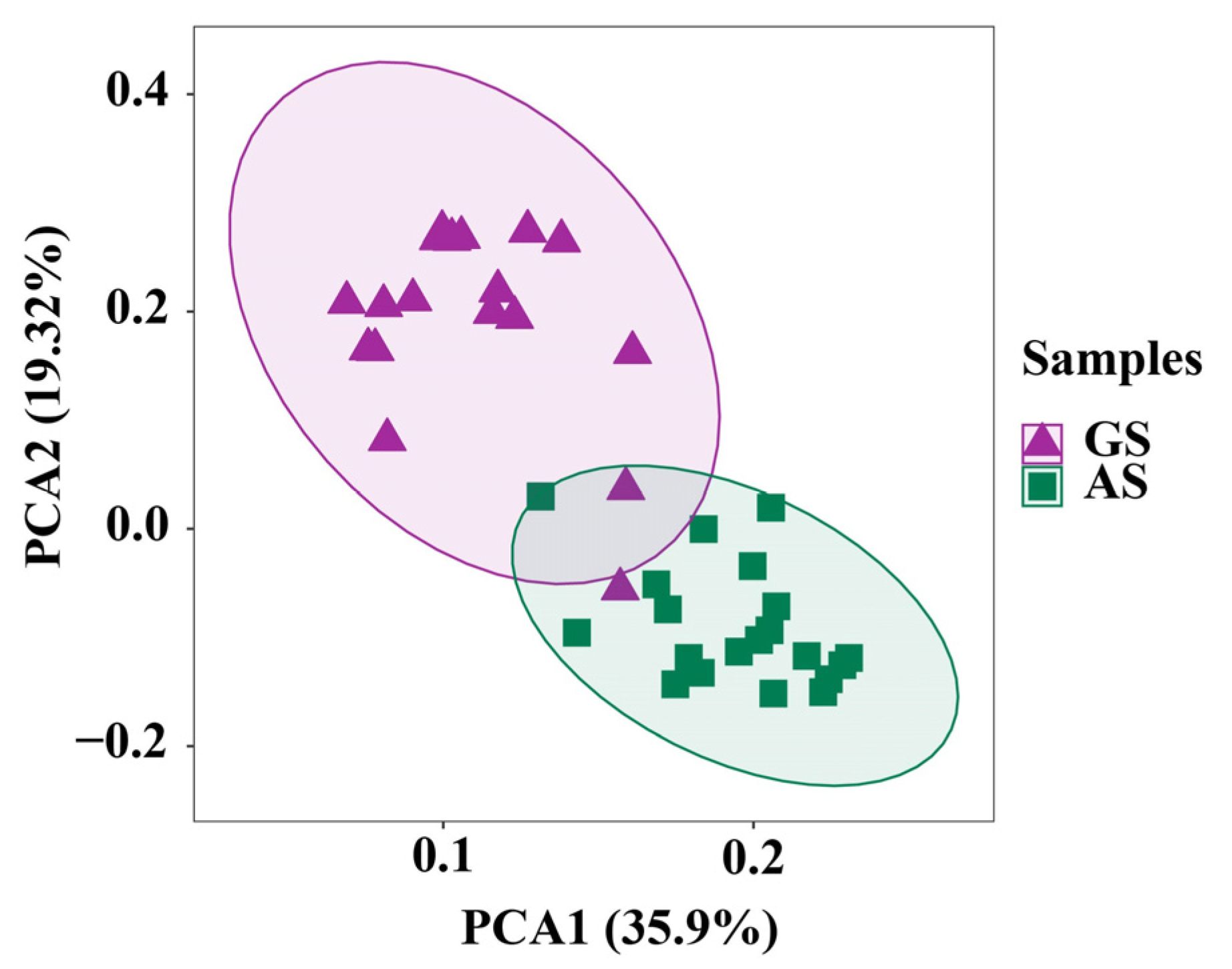
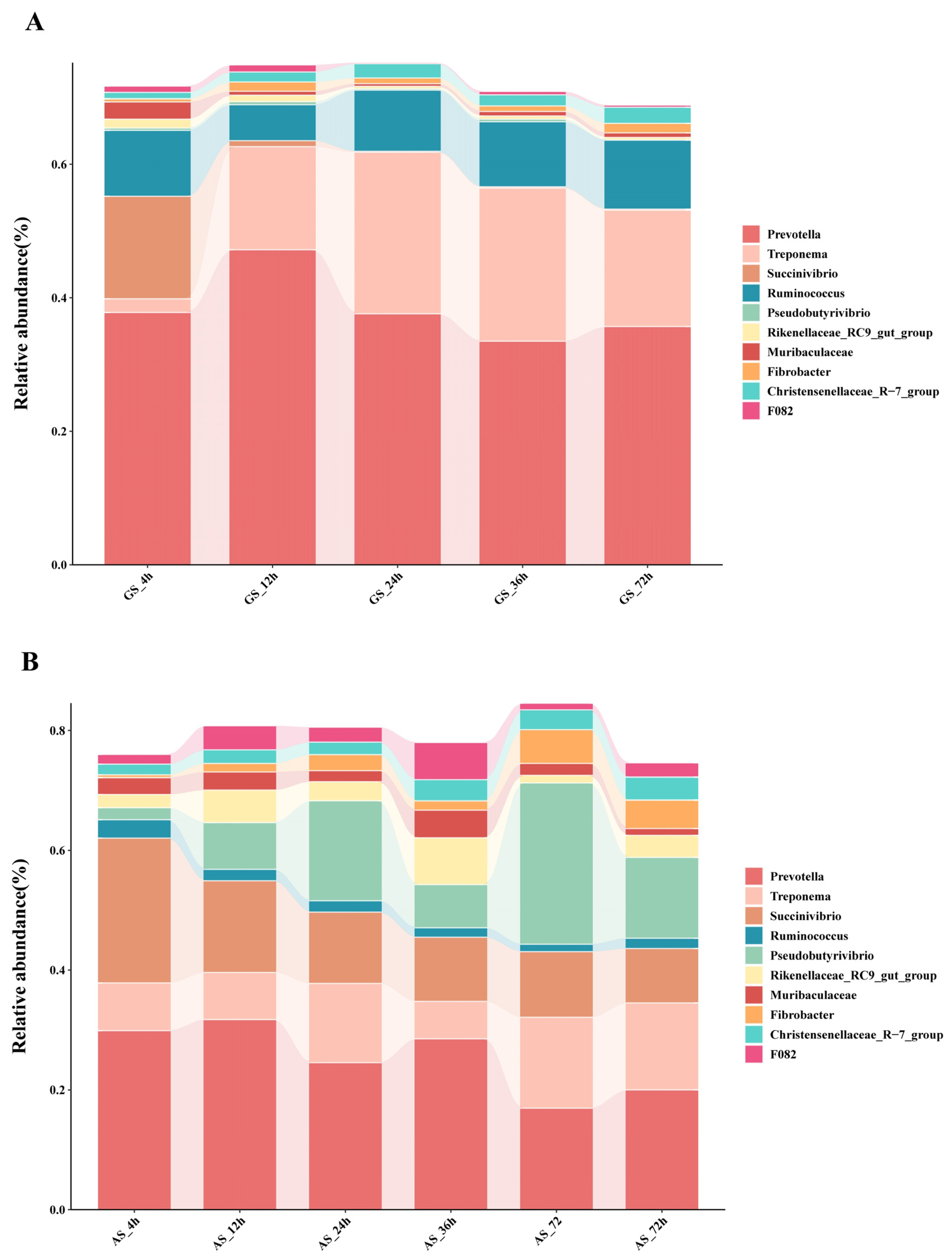
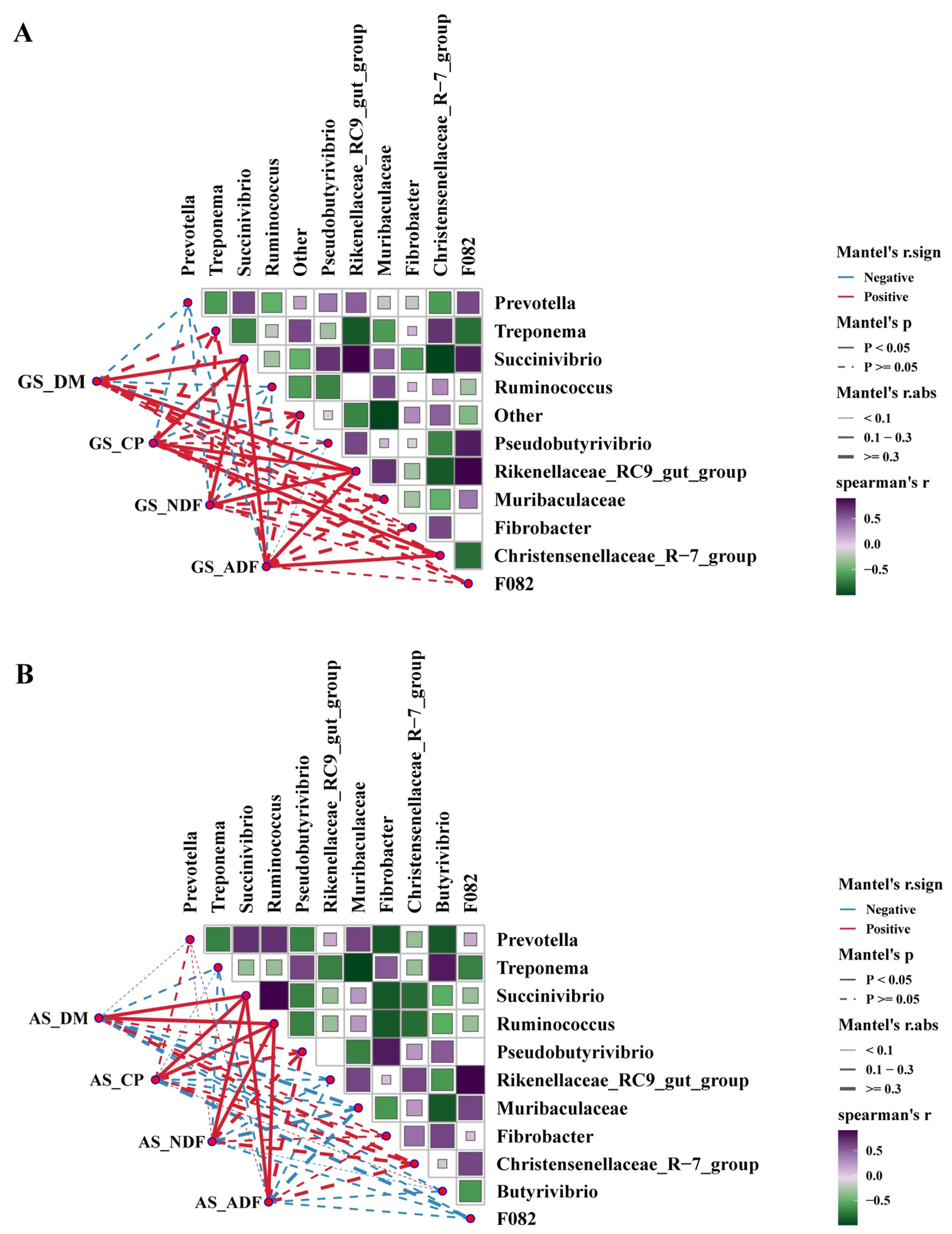
3.2. Rumen Microbiome Analysis
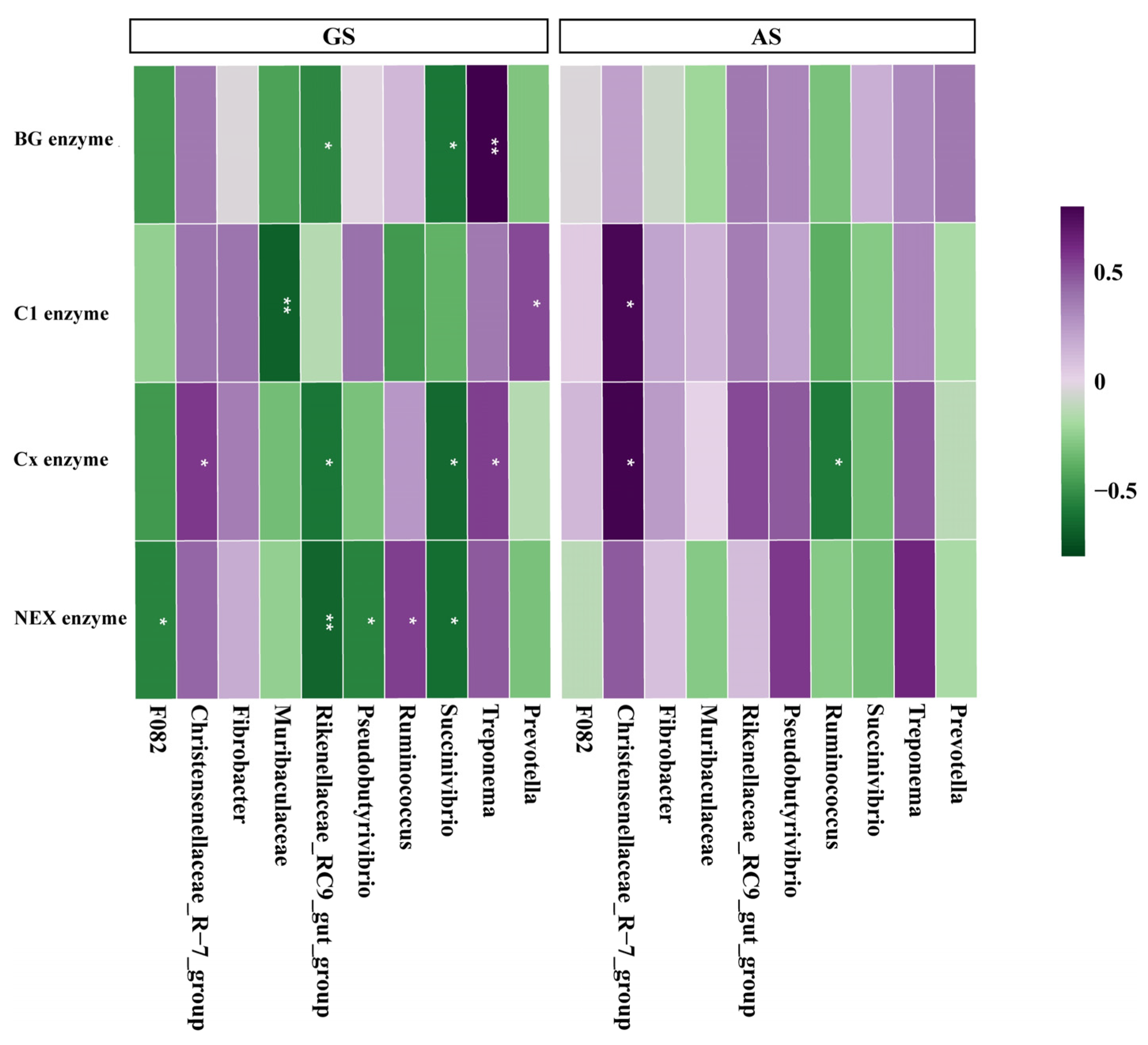
3.3. Cellulase Activity
3.4. The Correlation between Nutrients Degradation Rate, Cellulase Activity and Rumen Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alagawany, M.; Farag, M.R.; Sahfi, M.E.; Elnesr, S.S.; Alqaisi, O.; El-Kassas, S.; Al-Wajeeh, A.S.; Taha, A.E.; Abd E-Hack, M.E. Phytochemical characteristics of Paulownia trees wastes and its use as unconventional feedstuff in animal feed. Anim. Biotechnol. 2022, 33, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Salim, M.H.; Kassab, Z.; Abdellaoui, Y.; García-Cruz, A.; Soumare, A.; Ablouh, E.-H.; El Achaby, M. Exploration of multifunctional properties of garlic skin derived cellulose nanocrystals and extracts incorporated chitosan biocomposite films for active packaging application. Int. J. Biol. Macromol. 2022, 210, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Petropoulos, S.; Ferreira, I.C. Chemical composition and bioactive compounds of garlic (Allium sativum L.) as affected by pre- and post-harvest conditions: A review. Food Chem. 2016, 211, 41–50. [Google Scholar] [CrossRef]
- Sobenin, I.A.; Andrianova, I.V.; Lakunin, K.Y.; Karagodin, V.P.; Bobryshev, Y.V.; Orekhov, A.N. Anti-atherosclerotic effects of garlic preparation in freeze injury model of atherosclerosis in cholesterol-fed rabbits. Phytomedicine 2016, 23, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Carreón-Delgado, D.F.; Hernández-Montesinos, I.Y.; Rivera-Hernández, K.N.; Del Sugeyrol Villa-Ramírez, M.; Ochoa-Velasco, C.E.; Ramírez-López, C. Evaluation of Pretreatments and Extraction Conditions on the Antifungal and Antioxidant Effects of Garlic (Allium sativum) Peel Extracts. Plants 2023, 12, 217. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Su, Z.; Xu, W.; Sun, H.X.; Gao, J.F.; Tu, D.F.; Ren, C.H.; Zhang, Z.J.; Cao, H.G. Garlic skin induces shifts in the rumen microbiome and metabolome of fattening lambs. Animal 2021, 15, 100216. [Google Scholar] [CrossRef]
- Rahayu, T.; Prayitno, C.; Munasik, M. Consumption and Digestibility Value of Crude Fiber of Dairy Goat Feed with Supplementation Garlic Husk Flour and Organic Minerals (Cr, Se, and Zn-Lysinat). In International Conference on Improving Tropical Animal Production for Food Security (ITAPS 2021); Atlantis Press: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Tian, F.; Zhou, Z.; Wang, X.; Zhang, K.; Han, S. Changes in Soil Microbial Community along a Chronosequence of Perennial Mugwort Cropping in Northern China Plain. Agronomy 2022, 12, 1568. [Google Scholar] [CrossRef]
- Wen, W.; Xu, P.; Xiang, H.; Wen, M.; Ye, X.; Chu, C.; Tong, S. Comprehensive two-dimensional countercurrent chromatography × gas chromatography characterization of Artemisia argyi essential oil. Anal. Chim. Acta 2023, 1237, 340614. [Google Scholar] [CrossRef]
- Yu, D.; Huang, N.N.; Du, X.W. Review of the Chemical Composition and Biological Activities of Essential Oils from Artemisia Argyi, Artemisia Princeps, and Artemisia Montana. Curr. Top. Med. Chem. 2023, 23, 1522–1541. [Google Scholar] [CrossRef]
- Du, Z.; Risu, N.; Gentu, G.; Jia, Y.; Cai, Y. Growth performance, apparent digestibility, and N balance in Mongolian lambs and hoggs fed diets supplemented with a Chinese traditional herbal medicine complex. Anim. Sci. J. 2018, 89, 1451–1458. [Google Scholar] [CrossRef]
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The rumen microbiome: Balancing food security and environmental impacts. Nat. Rev. Microbiol. 2021, 19, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Hagen, L.H.; Brooke, C.G.; Shaw, C.A.; Norbeck, A.D.; Piao, H.; Arntzen, M.Ø.; Olson, H.M.; Copeland, A.; Isern, N.; Shukla, A.; et al. Proteome specialization of anaerobic fungi during ruminal degradation of recalcitrant plant fiber. ISME J. 2021, 15, 421–434. [Google Scholar] [CrossRef]
- Xu, Q.; Qiao, Q.; Gao, Y.; Hou, J.; Hu, M.; Du, Y.; Zhao, K.; Li, X. Gut Microbiota and Their Role in Health and Metabolic Disease of Dairy Cow. Front. Nutr. 2021, 8, 701511. [Google Scholar] [CrossRef] [PubMed]
- Selinger, L.B.; Forsberg, C.W.; Cheng, K.J. The Rumen: A Unique Source of Enzymes for Enhancing Livestock Production. Anaerobe 1996, 2, 263–284. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, N.R.; Kumar, S.; Kumari, R.; Bharti, V.K. A Review on Rumen Anaerobic Fungi: Current Understanding on Carbohydrate Fermentation and Roughages Digestion in Ruminants. Appl. Biochem. Microbiol. 2023, 59, 231–249. [Google Scholar] [CrossRef]
- Hasan, M.T.; AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Artington, VA, USA, 2015. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Ling, W.; Zhang, L.; Feng, Q.; Degen, A.A.; Li, J.; Qi, Y.; Li, Y.; Zhou, Y.; Liu, Y.; Yang, F.; et al. Effects of Different Additives on Fermentation Quality, Microbial Communities, and Rumen Degradation of Alfalfa Silage. Fermentation 2022, 8, 660. [Google Scholar] [CrossRef]
- Huang, X.; Xu, Y.; Wu, X.; Ding, Y.; Fan, C.; Xue, Y.; Zhuo, Z.; Cheng, J. Mixed Fermentation of Lactiplantibacillus plantarum and Bacillus licheniformis Changed the Chemical Composition, Bacterial Community, and Rumen Degradation Rate of Tea Residue. Fermentation 2022, 8, 380. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Wang, W.-K.; Wu, Q.-C.; Zhang, F.; Li, W.-J.; Li, S.-L.; Wang, W.; Cao, Z.-J.; Yang, H.-J. In Situ Rumen Degradation Characteristics and Bacterial Colonization of Corn Silages Differing in Ferulic and p-Coumaric Acid Contents. Microorganisms 2022, 10, 2269. [Google Scholar] [CrossRef]
- Ørskov, E.R.; McDonald, I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 1979, 92, 499–503. [Google Scholar] [CrossRef]
- Kennedy, P.M.; Murphy, M.R. The nutritional implications of differential passage of particles through the ruminant alimentary tract. Nutr. Res. Rev. 1988, 1, 189–208. [Google Scholar] [CrossRef] [PubMed]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH assembly of TALENs for high-throughput genome editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef]
- McCarthy, K.M.; Lynch, M.B.; Pierce, K.M.; Fahey, A.G.; Gath, V.P.; McDonald, M.; Boland, T.M.; Sheridan, H.; Markiewicz-Keszycka, M.; Mulligan, F.J. Rumen fermentation and forage degradability in dairy cows offered perennial ryegrass, perennial ryegrass and white clover, or a multispecies forage. Livest. Sci. 2023, 269, 105185. [Google Scholar] [CrossRef]
- Luo, C.; Wang, D.; Lu, N.; Li, H.; Liu, G.; Cao, Z.; Yang, H.; Li, S.; Yu, X.; Shao, W.; et al. Analysis of Chemical Composition, Amino Acid Content, and Rumen Degradation Characteristics of Six Organic Feeds. Animals 2022, 12, 682. [Google Scholar] [CrossRef] [PubMed]
- Das, L.; Kundu, S.s.; Datt, C.; Kumar, D.; Tariq, H. In Situ Ruminal Degradation Kinetics of Dry Matter, Crude Protein and Neutral Detergent Fiber of Tropical Ruminant Feedstuffs. Indian J. Anim. Nutr. 2015, 32, 45–51. [Google Scholar]
- Li, Q.; Xue, B.; Zhao, Y.; Wu, T.; Liu, H.; Yi, X.; Sun, C.; Wang, Z.; Zou, H.; Yan, T. In situ degradation kinetics of 6 roughages and the intestinal digestibility of the rumen undegradable protein1. J. Anim. Sci. 2018, 96, 4835–4844. [Google Scholar] [CrossRef] [PubMed]
- Broderick, G.A. Effects of varying dietary protein and energy levels on the production of lactating dairy cows. J. Dairy Sci. 2003, 86, 1370–1381. [Google Scholar] [CrossRef]
- Mekuriaw, Y.; Asmare, B. Nutrient intake, digestibility and growth performance of Washera lambs fed natural pasture hay supplemented with graded levels of Ficus thonningii(Chibha) leaves as replacement for concentrate mixture. Agric. Food Secur. 2018, 7, 30. [Google Scholar] [CrossRef]
- Zhao, S.; Zhou, S.; Zhao, Y.; Yang, J.; Lv, L.; Zheng, Z.; Lu, H.; Ren, Y. Comparative Study of the Nutritional Value and Degradation Characteristics of Amaranth Hay in the Rumen of Goats at Different Growth Stages. Animals 2022, 13, 25. [Google Scholar] [CrossRef]
- Rehemujiang, H.; Yusuf, H.A.; Ma, T.; Diao, Q.; Kong, L.; Kang, L.; Tu, Y. Evaluating Fermentation Quality, Aerobic Stability, and Rumen-Degradation (In Situ) Characteristics of Various Protein-Based Total Mixed Rations. Animals 2023, 13, 2730. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Sievert, S.J.; Shaver, R.D.; Welch, D.A.; Combs, D.K. In situ dry matter, protein, and fiber degradation of perennial forages. J. Dairy Sci. 1993, 76, 2632–2643. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.X.; Zhang, X.M.; Li, Q.S.; Wang, R.; Zhang, M.; Zhang, S.Z.; Lin, B.; Tan, B.; Tan, Z.L.; Wang, M. Comparison of in situ ruminal straw fiber degradation and bacterial community between buffalo and Holstein fed with high-roughage diet. Front. Microbiol. 2022, 13, 1079056. [Google Scholar] [CrossRef] [PubMed]
- Ransom-Jones, E.; Jones, D.L.; McCarthy, A.J.; McDonald, J.E. The Fibrobacteres: An important phylum of cellulose-degrading bacteria. Microb. Ecol. 2012, 63, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Yu, Y.; Lv, Y.; Wang, D.; Li, H.; Li, Z.; Zhang, Y.; Chen, L.; Leng, J. Metagenomic Sequencing Reveals the Taxonomic and Functional Characteristics of Rumen Micro-organisms in Gayals. Microorganisms 2023, 11, 1098. [Google Scholar] [CrossRef] [PubMed]
- Gharechahi, J.; Vahidi, M.F.; Sharifi, G.; Ariaeenejad, S.; Ding, X.-Z.; Han, J.-L.; Salekdeh, G.H. Lignocellulose degradation by rumen bacterial communities: New insights from metagenome analyses. Environ. Res. 2023, 229, 115925. [Google Scholar] [CrossRef] [PubMed]
- Pinnell, L.J.; Reyes, A.A.; Wolfe, C.A.; Weinroth, M.D.; Metcalf, J.L.; Delmore, R.J.; Belk, K.E.; Morley, P.S.; Engle, T.E. Bacteroidetes and Firmicutes Drive Differing Microbial Diversity and Community Composition Among Micro-Environments in the Bovine Rumen. Front. Vet. Sci. 2022, 9, 897996. [Google Scholar] [CrossRef]
- Fernando, S.C.; Purvis, H.T., 2nd; Najar, F.Z.; Sukharnikov, L.O.; Krehbiel, C.R.; Nagaraja, T.G.; Roe, B.A.; Desilva, U. Rumen microbial population dynamics during adaptation to a high-grain diet. Appl. Environ. Microbiol. 2010, 76, 7482–7490. [Google Scholar] [CrossRef]
- Wang, M.; Wu, L.; Guo, Y.; Sun, J.; Deng, M.; Liu, G.; Li, Y.; Sun, B. Effects of fermented herbal tea residue on meat quality, rumen fermentation parameters and microbes of black goats. AMB Express 2023, 13, 106. [Google Scholar] [CrossRef]
- Greiner, L.; Graham, A.; Goncalves, M.; Orlando, U.; Touchette, K.J. Evaluation of the optimal standardized ileal digestible valine:lysine ratio in lactating sow diets1. J. Anim. Sci. 2019, 97, 2965–2971. [Google Scholar] [CrossRef]
- Paster, B.J.; Canale-Parola, E. Physiological diversity of rumen spirochetes. Appl. Environ. Microbiol. 1982, 43, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Stanton, T.B.; Canale-Parola, E. Enumeration and selective isolation of rumen spirochetes. Appl. Environ. Microbiol. 1979, 38, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.P.; McCormick, C.A.; Suen, G. Fibrobacter communities in the gastrointestinal tracts of diverse hindgut-fermenting herbivores are distinct from those of the rumen. Environ. Microbiol. 2017, 19, 3768–3783. [Google Scholar] [CrossRef] [PubMed]
- Senés-Guerrero, C.; Colón-Contreras, F.A.; Reynoso-Lobo, J.F.; Tinoco-Pérez, B.; Siller-Cepeda, J.H.; Pacheco, A. Biogas-producing microbial composition of an anaerobic digester and associated bovine residues. Microbiologyopen 2019, 8, e00854. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, M.; Xue, C.; Zhu, W.; Mao, S. Characterization and comparison of the temporal dynamics of ruminal bacterial microbiota colonizing rice straw and alfalfa hay within ruminants. J. Dairy Sci. 2016, 99, 9668–9681. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, Y.; Li, Y.; Zhang, Y.; Liu, T.; Wang, Y.; Sharpton, T.J.; Zhu, W. Progressive Colonization of Bacteria and Degradation of Rice Straw in the Rumen by Illumina Sequencing. Front. Microbiol. 2017, 8, 2165. [Google Scholar] [CrossRef]
- Piao, H.; Lachman, M.; Malfatti, S.; Sczyrba, A.; Knierim, B.; Auer, M.; Tringe, S.G.; Mackie, R.I.; Yeoman, C.J.; Hess, M. Temporal dynamics of fibrolytic and methanogenic rumen microorganisms during in situ incubation of switchgrass determined by 16S rRNA gene profiling. Front. Microbiol. 2014, 5, 307. [Google Scholar] [CrossRef]
- Betancur-Murillo, C.L.; Aguilar-Marín, S.B.; Jovel, J. Prevotella: A Key Player in Ruminal Metabolism. Microorganisms 2023, 11, 1. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, K.; Li, M.; Fan, H.; Wang, M.; Xia, S.; Chen, Y.; Bai, X.; Liu, Z.; Ni, J.; et al. A Preliminary Study of the Potential Molecular Mechanisms of Individual Growth and Rumen Development in Calves with Different Feeding Patterns. Microorganisms 2023, 11, 2423. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Z.; Tan, Y.; Chang, S.; Zheng, H.; Wang, H.; Yan, T.; Guru, T.; Hou, F. Selenium Yeast Dietary Supplement Affects Rumen Bacterial Population Dynamics and Fermentation Parameters of Tibetan Sheep (Ovis aries) in Alpine Meadow. Front. Microbiol. 2021, 12, 663945. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, T.; Wang, X.; Geng, Y.; Zhao, N.; Hu, L.; Liu, H.; Kang, S.; Xu, S. Effect of Dietary Protein Levels on Dynamic Changes and Interactions of Ruminal Microbiota and Metabolites in Yaks on the Qinghai-Tibetan Plateau. Front. Microbiol. 2021, 12, 684340. [Google Scholar] [CrossRef] [PubMed]
- Vahidi, M.F.; Gharechahi, J.; Behmanesh, M.; Ding, X.Z.; Han, J.L.; Hosseini Salekdeh, G. Diversity of microbes colonizing forages of varying lignocellulose properties in the sheep rumen. PeerJ 2021, 9, e10463. [Google Scholar] [CrossRef] [PubMed]
- Ozbayram, E.G.; Kleinsteuber, S.; Nikolausz, M. Biotechnological utilization of animal gut microbiota for valorization of lignocellulosic biomass. Appl. Microbiol. Biotechnol. 2020, 104, 489–508. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Liu, B.; La, S.; Li, D.; Zhu, X.; Sun, H.; Ma, S.; Cui, Y.; Shi, Y. Alfalfa hay substitution for wheat straw improves beef quality via rumen microflora alteration. Heliyon 2023, 9, e20803. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Ahmed, J.; Gavande, P.V.; Fontes, C.M.G.A.; Goyal, A. Structural and functional insights into the glycoside hydrolase family 30 xylanase of the rumen bacterium Ruminococcus flavefaciens. J. Mol. Struct. 2023, 1272, 134155. [Google Scholar] [CrossRef]
- Dixit, S.; Kumar, S.; Sharma, R.; Banakar, P.; Singh, M.; Keshri, A.; Tyagi, A. Rumen multi-omics addressing diet–host–microbiome interplay in farm animals: A review. Anim. Biotechnol. 2023, 34, 3187–3205. [Google Scholar] [CrossRef] [PubMed]
- Arriola, K.G.; Oliveira, A.S.; Ma, Z.X.; Lean, I.J.; Giurcanu, M.C.; Adesogan, A.T. A meta-analysis on the effect of dietary application of exogenous fibrolytic enzymes on the performance of dairy cows. J. Dairy Sci. 2017, 100, 4513–4527. [Google Scholar] [CrossRef]
- Hua, D.; Hendriks, W.H.; Xiong, B.; Pellikaan, W.F. Starch and Cellulose Degradation in the Rumen and Applications of Metagenomics on Ruminal Microorganisms. Animals 2022, 12, 3020. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, J.; Gui, H.; Wang, H.; Li, Y.; Zhang, J.; Cao, S.; Zhong, J.; Qian, Y.; Meng, C. Effect of Garlic Straw with Silage Corn Stalks on Hu Sheep Rumen Fermentation and Microbial Community In Vitro. Metabolites 2023, 13, 1201. [Google Scholar] [CrossRef]
- Zhang, F.; Xu, J.; Hu, Y.; Fang, J.; Yang, M.; Huang, K.; Xu, W.; He, X. Diallyl trisulfide ameliorates bone loss and alters specific gut microbiota and serum metabolites in natural aging mice. Food Funct. 2023, 14, 7642–7653. [Google Scholar] [CrossRef]
- Xu, Y.; Aung, M.; Sun, Z.; Zhou, Y.; Xue, T.; Cheng, X.; Cheng, Y.; Hao, L.; Zhu, W.; Degen, A. Ensiling of rice straw enhances the nutritive quality, improves average daily gain, reduces in vitro methane production and increases ruminal bacterial diversity in growing Hu lambs. Anim. Feed. Sci. Technol. 2023, 295, 115513. [Google Scholar] [CrossRef]
- Han, Y.; Chang, X.; Xiang, H.; Fang, Y.; Hao, L.; Gu, Y.; Han, X.; Zhao, B.; Zhao, Z.; Zhao, C.; et al. Exploring biomimetic potential of ruminant digestion strategies for lignocellulosic biomass utilization: A comprehensive review. Renew. Sustain. Energy Rev. 2023, 188, 113887. [Google Scholar] [CrossRef]
- Won, M.-Y.; Oyama, L.B.; Courtney, S.J.; Creevey, C.J.; Huws, S.A. Can rumen bacteria communicate to each other? Microbiome 2020, 8, 23. [Google Scholar] [CrossRef]
- Hao, Y.; Guo, C.; Gong, Y.; Sun, X.; Wang, W.; Wang, Y.; Yang, H.; Cao, Z.; Li, S. Rumen Fermentation, Digestive Enzyme Activity, and Bacteria Composition between Pre-Weaning and Post-Weaning Dairy Calves. Animals 2021, 11, 2527. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Nutrient Composition | ||||
|---|---|---|---|---|---|
| DM | CP | NDF | ADF | HC | |
| GS | 87.36 ± 0.83 b | 12.40 ± 1.02 a | 34.17 ± 1.80 b | 29.92 ± 1.93 b | 4.25 ± 0.69 b |
| AS | 91.27 ± 1.26 a | 8.19 ± 0.50 b | 83.62 ± 0.39 a | 59.54 ± 1.59 a | 24.08 ± 1.38 a |
| Items | Content |
|---|---|
| Ingredients | |
| Leymus chinensis | 55.00 |
| Corn | 31.00 |
| Soybean meal | 12.20 |
| Salt | 0.50 |
| Stone powder | 0.10 |
| Calcium bicarbonate | 0.20 |
| Premix 1 | 1.00 |
| Total | 100 |
| Nutrient composition 2 | |
| Crude protein | 10.94 |
| Neutral detergent fiber | 48.66 |
| Acid detergent fiber | 24.83 |
| Ether extract | 2.36 |
| Calcium | 0.62 |
| Phosphorus | 0.35 |
| Net energy for lactation (MJ/kg DM) | 11.67 |
| Items | Time | GS | AS |
|---|---|---|---|
| Chao1 | 4 h | 304.37 ± 16.80 | 342.71 ± 35.74 |
| 12 h | 366.42 ± 24.18 B | 499.70 ± 53.82 A | |
| 24 h | 313.86 ± 65.66 B | 509.87 ± 14.93 A | |
| 36 h | 373.45 ± 38.49 B | 623.76 ± 61.37 A | |
| 72 h | 357.94 ± 59.91 B | 584.29 ± 15.51 A | |
| ACE | 4 h | 340.04 ± 40.60 | 305.68 ± 18.87 |
| 12 h | 362.45 ± 25.67 B | 492.93 ± 51.84 A | |
| 24 h | 311.99 ± 63.55 B | 528.23 ± 24.60 A | |
| 36 h | 411.32 ± 34.96 B | 620.92 ± 68.00 A | |
| 72 h | 354.32 ± 56.36 B | 579.46 ± 17.52 A | |
| Shannon | 4 h | 5.98 ± 0.39 | 5.59 ± 0.29 |
| 12 h | 6.38 ± 0.11 | 6.73 ± 0.61 | |
| 24 h | 6.06 ± 0.17 | 6.47 ± 0.48 | |
| 36 h | 6.36 ± 0.29 | 7.18 ± 0.49 | |
| 72 h | 6.17 ± 0.25 B | 6.98 ± 0.27 A | |
| Simpson | 4 h | 0.94 ± 0.03 | 0.94 ± 0.05 |
| 12 h | 0.96 ± 0.01 | 0.97 ± 0.02 | |
| 24 h | 0.96 ± 0.00 | 0.96 ± 0.01 | |
| 36 h | 0.97 ± 0.00 | 0.98 ± 0.01 | |
| 72 h | 0.97 ± 0.00 | 0.97 ± 0.02 | |
| Coverage | 4 h | 0.99 | 0.99 |
| 12 h | 0.99 | 0.99 | |
| 24 h | 0.99 | 0.99 | |
| 36 h | 0.99 | 0.99 | |
| 72 h | 0.99 | 0.99 |
| Items | Time | GS | AS |
|---|---|---|---|
| Bacteroidetes | 4 h | 49.58 ± 5.27 | 42.90 ± 5.04 ab |
| 12 h | 54.83 ± 3.79 | 50.91 ± 5.57 a | |
| 24 h | 43.99 ± 5.84 | 38.32 ± 8.43 ab | |
| 36 h | 40.97 ± 4.11 B | 54.07 ± 3.70 Aa | |
| 72 h | 44.48 ± 6.62 | 31.09 ± 7.31 b | |
| Firmicutes | 4 h | 31.59 ± 5.92 | 23.07 ± 6.38 ab |
| 12 h | 26.54 ± 6.81 | 22.90 ± 4.95 b | |
| 24 h | 30.32 ± 9.75 | 32.32 ± 4.40 ab | |
| 36 h | 34.42 ± 7.05 | 25.30 ± 3.94 ab | |
| 72 h | 36.10 ± 3.65 | 38.04 ± 7.46 a | |
| Spirochaetes | 4 h | 2.07 ± 2.37 b | 7.99 ± 10.73 |
| 12 h | 15.41 ± 2.94 a | 7.97 ± 4.30 | |
| 24 h | 24.23 ± 4.59 Aa | 13.36 ± 2.80 B | |
| 36 h | 22.93 ± 3.00 Aa | 6.37 ± 3.63 B | |
| 72 h | 17.43 ± 6.46 a | 14.91 ± 0.87 | |
| Proteobacteria | 4 h | 16.04 ± 9.48 a | 24.95 ± 14.52 |
| 12 h | 1.69 ± 0.69 b | 16.08 ± 13.49 | |
| 24 h | 0.53 ± 0.56 b | 12.66 ± 9.68 | |
| 36 h | 0.68 ± 0.22 b | 11.56 ± 8.36 | |
| 72 h | 0.42 ± 0.19 Bb | 10.15 ± 5.78 A |
| Items | Time | GS | AS |
|---|---|---|---|
| BG enzyme (nmol/min/g) | 4 h | 118.01 ± 10.2 Bd | 141.97 ± 3.7 Ab |
| 12 h | 135.81 ± 1.58 Bcd | 185.33 ± 5.18 Aa | |
| 24 h | 179.35 ± 10.08 b | 190.08 ± 13.73 a | |
| 36 h | 247.90 ± 15.18 Aa | 173.45 ± 19.19 Bab | |
| 72 h | 152.40 ± 8.38 bc | 165.38 ± 22.46 ab | |
| C1 enzyme (nmol/min/g) | 4 h | 1113.06 ± 52.41 Bc | 2696.35 ± 104.63 Ab |
| 12 h | 3135.52 ± 170.74 Aa | 1914.42 ± 204.05 Bc | |
| 24 h | 2931.15 ± 131.95 ab | 3100.23 ± 88.85 b | |
| 36 h | 2693.80 ± 152.47 Bb | 3697.37 ± 225.66 Aa | |
| 72 h | 2762.01 ± 149.84 b | 3058.61 ± 201.28 b | |
| Cx enzyme (nmol/min/g) | 4 h | 1563.18 ± 105.90 d | 1573.46 ± 67.00 c |
| 12 h | 1901.04 ± 89.95 Bc | 2396.90 ± 125.45 Ab | |
| 24 h | 2081.71 ± 133.63 bc | 3131.45 ± 230.17 a | |
| 36 h | 2222.04 ± 116.64 Bb | 3291.45 ± 103.87 Aa | |
| 72 h | 3644.36 ± 133.21 Aa | 3001.92 ± 136.63 Ba | |
| NEX enzyme (nmol/min/g) | 4 h | 213.59 ± 20.57 d | 211.70 ± 12.71 d |
| 12 h | 189.97 ± 8.74 Bd | 204.64 ± 0.95 Ad | |
| 24 h | 321.35 ± 4.32 Bb | 438.95 ± 21.32 Aa | |
| 36 h | 250.24 ± 5.51 Bc | 360.70 ± 27.67 Ab | |
| 72 h | 425.62 ± 17.49 Aa | 273.22 ± 35.00 Bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, M.; Liu, H.; Jiang, X.; Qiu, S.; Li, K.; Lu, J.; Zhang, M.; Qiu, Y.; Wang, B.; Ma, Z.; et al. Analysis of Rumen Degradation Characteristics, Attached Microbial Community, and Cellulase Activity Changes of Garlic Skin and Artemisia argyi Stalk. Animals 2024, 14, 169. https://doi.org/10.3390/ani14010169
Gu M, Liu H, Jiang X, Qiu S, Li K, Lu J, Zhang M, Qiu Y, Wang B, Ma Z, et al. Analysis of Rumen Degradation Characteristics, Attached Microbial Community, and Cellulase Activity Changes of Garlic Skin and Artemisia argyi Stalk. Animals. 2024; 14(1):169. https://doi.org/10.3390/ani14010169
Chicago/Turabian StyleGu, Mingming, Haoyu Liu, Xinghui Jiang, Shuiling Qiu, Keyao Li, Jianing Lu, Mingrui Zhang, Yujun Qiu, Benzhi Wang, Zhiyi Ma, and et al. 2024. "Analysis of Rumen Degradation Characteristics, Attached Microbial Community, and Cellulase Activity Changes of Garlic Skin and Artemisia argyi Stalk" Animals 14, no. 1: 169. https://doi.org/10.3390/ani14010169
APA StyleGu, M., Liu, H., Jiang, X., Qiu, S., Li, K., Lu, J., Zhang, M., Qiu, Y., Wang, B., Ma, Z., & Gan, Q. (2024). Analysis of Rumen Degradation Characteristics, Attached Microbial Community, and Cellulase Activity Changes of Garlic Skin and Artemisia argyi Stalk. Animals, 14(1), 169. https://doi.org/10.3390/ani14010169