

First Report of Sarcocystis pilosa from a Red Fox (Vulpes vulpes) Released for the Re-Introduction Project in South Korea

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Sample Collection
2.2. Fecal Flotation and Cyst Observation
2.3. DNA Extraction and PCR Sequencing
2.4. Sequence and Phylogenetic Analysis
3. Results
3.1. Morphological Observations
3.2. Molecular Characteristics
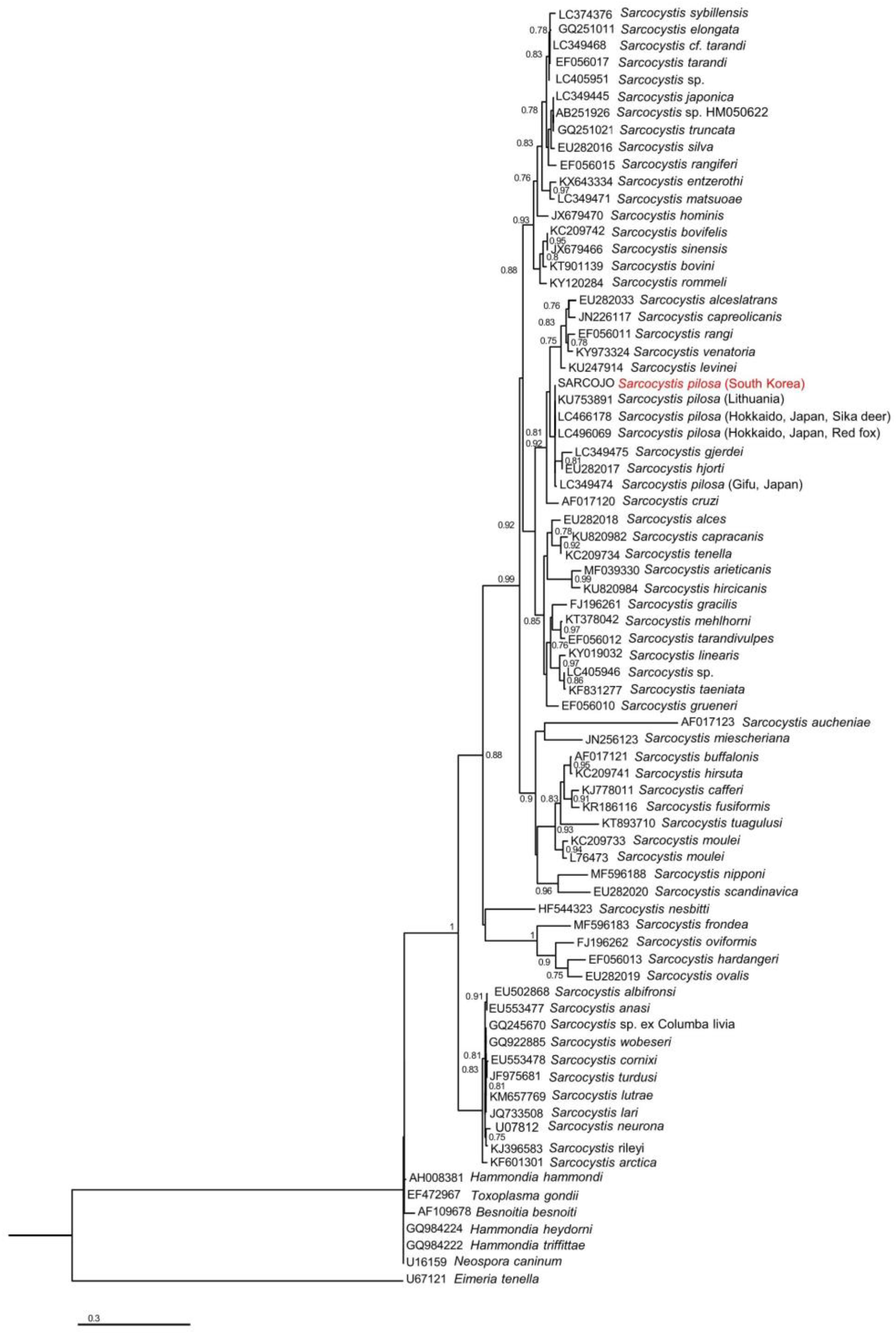
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, H.J.; Lee, B.K.; Kwon, G.H.; Chung, C.U. Release strategy for the Red fox (Vulpes vulpes) restoration project in Korea Based on Population Viability Analysis. Korean J. Environ. Ecol. 2013, 27, 417–428. [Google Scholar]
- Yu, J.N.; Han, S.H.; Kim, B.H.; Kryukov, A.P.; Kim, S.; Lee, B.Y.; Kwak, M. Insights into Korean red fox (Vulpes vulpes) based on mitochondrial cytochrome b sequence variation in East Asia. Zool. Sci. 2012, 29, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Garcês, A.; Pires, I. Secrets of the astute Red Fox (Vulpes vulpes, Linnaeus, 1758): An inside-ecosystem secret agent serving one health. Environments 2021, 8, 103. [Google Scholar] [CrossRef]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Chapter 1—General Biology. In Sarcocystosis of Animals and Humans; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–108. [Google Scholar]
- Moon, M.H. Sarcocystis infection and identification of Sarcocystis species in pigs in Korea. Korean J. Vet. Res. 1989, 29, 325–331. [Google Scholar]
- Wee, S.H.; Shin, S.S. Experimental induction of the two-host life cycle of Sarcocystis cruzi between dogs and Korean native calves. Korean J. Parasitol. 2001, 39, 227–232. [Google Scholar] [CrossRef]
- Son, H.; Kim, N.; Ryu, S.; Kim, H.; Rhee, J.; Cho, J.; Park, B. Ultrastructure of Sarcocystis grueneri-like sarcocysts from cardiac muscle of red deer (Cervus elaphus) in Korea. J. Vet. Clin. 2009, 26, 595–599. [Google Scholar]
- Kim, T.-H.; Han, J.-H.; Chang, S.-N.; Kim, D.-S.; Abdelkader, T.-S.; Seok, S.-H.; Park, J.-H.; Oh, H.-S.; Kim, J.-T.; Lee, B.-H.; et al. Detection of sarcocystic infection in a wild rodent (Apodemus agrarius chejuensis) captured on Jeju island. Lab. Anim. Res. 2011, 27, 357–359. [Google Scholar] [CrossRef]
- Hong, E.J.; Sim, C.; Chae, J.S.; Kim, H.C.; Park, J.; Choi, K.S.; Yu, D.H.; Park, C.H.; Yoo, J.G.; Park, B.K. Ultrastructural and molecular identification of Sarcocystis tenella (Protozoa, Apicomplexa) in naturally infected Korean native goats. Vet. Med. 2016, 61, 374–381. [Google Scholar] [CrossRef]
- Kim, H.-W.; Kim, H.-C.; Ryu, S.-Y.; Choi, K.-S.; Yu, D.-H.; Park, J.; Chae, J.-S.; Park, B.K. Molecular Identification of Sarcocystis grueneri in Wild Korean Water Deer (Hydropotes inermis argyropus). Korean J. Parasitol. 2018, 56, 129–134. [Google Scholar] [CrossRef]
- Dubey, J.P. Experimental infections of Sarcocystis cruzi, Sarcocystis tenella, Sarcocystis capracanis and Toxoplasma gondii in red foxes (Vulpes vulpes). J. Wildl. Dis. 1983, 19, 200–203. [Google Scholar] [CrossRef]
- Ford, G.E. Hosts of two canid genera, the red fox and the dog, as alternate vectors in the transmission of Sarcocystis tenella from sheep. Vet. Parasitol. 1987, 26, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, S.S.; Gjerde, B. The red fox (Vulpes vulpes) and the arctic fox (Vulpes lagopus) are definitive hosts of Sarcocystis alces and Sarcocystis hjorti from moose (Alces alces). Parasitology 2010, 137, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Irie, T.; Uraguchi, K.; Ito, T.; Yamazaki, A.; Takai, S.; Yagi, K. First report of Sarcocystis pilosa sporocysts in feces from red fox, Vulpes vulpes schrencki, in Hokkaido, Japan. Int. J. Parasitol. Parasites Wildl. 2020, 11, 29–31. [Google Scholar] [CrossRef]
- Fayer, R.; Esposito, D.H.; Dubey, J.P. Human infections with Sarcocystis species. Clin. Microbiol. Rev. 2015, 28, 295–311. [Google Scholar] [CrossRef]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Doležel, D.; Koudela, B.; Jirků, M.; Hypša, V.; Obornık, M.; Votýpka, J.; Modrý, D.; Šlapeta, J.R.; Lukeš, J. Phylogenetic analysis of Sarcocystis spp. of mammals and reptiles supports the coevolution of Sarcocystis spp. with their final hosts. Int. J. Parasitol. 1999, 29, 795–798. [Google Scholar] [CrossRef]
- Prakas, P.; Butkauskas, D.; Rudaitytė, E.; Kutkienė, L.; Sruoga, A.; Pūraitė, I. Morphological and molecular characterization of Sarcocystis taeniata and Sarcocystis pilosa n. sp. from the sika deer (Cervus nippon) in Lithuania. Parasitol. Res. 2016, 115, 3021–3032. [Google Scholar] [CrossRef]
- Dahlgren, S.S.; Gjerde, B. Genetic characterisation of six Sarcocystis species from reindeer (Rangifer tarandus tarandus) in Norway based on the small subunit rRNA gene. Vet. Parasitol. 2007, 146, 204–213. [Google Scholar] [CrossRef]
- Irie, T.; Ichii, O.; Nakamura, T.; Ikeda, T.; Ito, T.; Yamazaki, A.; Takai, S.; Yagi, K. Molecular characterization of three Sarcocystis spp. from wild sika deer (Cervus nippon yesoensis) in Hokkaido, Japan. Vet. Parasitol. Reg. Stud. Rep. 2019, 18, 100327. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B. Sarcocystis species in red deer revisited: With a re-description of two known species as Sarcocystis elongata n. sp. and Sarcocystis truncata n. sp. based on mitochondrial cox1 sequences. Parasitology 2014, 141, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Máca, O.; González-Solís, D. Role of three bird species in the life cycle of two Sarcocystis spp. (Apicomplexa, Sarcocystidae) in the Czech Republic. Int. J. Parasitol. Parasites Wildl. 2022, 17, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Basso, W.; Rojas, C.A.A.; Buob, D.; Ruetten, M.; Deplazes, P. Sarcocystis infection in red deer (Cervus elaphus) with eosinophilic myositis/fasciitis in Switzerland and involvement of red foxes (Vulpes vulpes) and hunting dogs in the transmission. Int. J. Parasitol. Parasites Wildl. 2020, 13, 130–141. [Google Scholar] [CrossRef]
- Abe, N.; Matsuo, K.; Moribe, J.; Takashima, Y.; Irie, T.; Baba, T.; Gjerde, B. Morphological and molecular characteristics of seven Sarcocystis species from sika deer (Cervus nippon centralis) in Japan, including three new species. Int. J. Parasitol. Parasites Wildl 2019, 10, 252–262. [Google Scholar] [CrossRef]
- Prakas, P.; Rehbein, S.; Rudaitytė-Lukošienė, E.; Butkauskas, D. Molecular identification of Sarcocystis species in sika deer (Cervus nippon) of free-ranging populations in Germany and Austria. Vet. Res. Commun. 2023, 47, 2165–2171. [Google Scholar] [CrossRef]
- Porter, R.A.; Ginn, P.E.; Dame, J.B.; Greiner, E.C. Evaluation of the shedding of Sarcocystis falcatula sporocysts in experimentally infected Virginia opossums (Didelphis virginiana). Vet. Parasitol. 2001, 95, 313–319. [Google Scholar] [CrossRef]
- Dubey, J.P.; Streitel, R.H. Shedding of Sarcocystis in feces of dogs and cats fed muscles of naturally infected food animals in the midwestern United States. J. Parasitol. 1976, 62, 828–830. [Google Scholar] [CrossRef]
- Gorman, T.R.; Alcaíno, H.A.; Muñuz, H.; Cunazza, C. Sarcocystis sp. in guanaco (Lama guanicoe) and effect of temperature on its viability. Vet. Parasitol. 1984, 15, 95–101. [Google Scholar] [CrossRef]
- Tsukada, H.; Nonaka, N. Foraging behavior of red foxes Vulpes vulpes schrencki utilizing human food in the Shiretoko National Park, Hokkaido. Mammal Study 1996, 21, 137–151. [Google Scholar] [CrossRef]
- Saito, M.; Itagaki, H. Experimental infection of raccoon dogs with Sarcocystis cruzi and S. miescheriana. J. Vet. Med. Sci. 1994, 56, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Solcan, C.; Acatrinei, D.; Floristean, V.; Solcan, G.; Şlencu, B.G.; Fântânariu, M. An unusual case of megacolon due to Sarcocystis spp. infection and local amyloidosis in a Husky dog. Pak. Vet. J. 2015, 35, 531–533. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, Y.; Lee, S.J.; Bia, M.M.; Choe, S.; Jeong, D.-H. First Report of Sarcocystis pilosa from a Red Fox (Vulpes vulpes) Released for the Re-Introduction Project in South Korea. Animals 2024, 14, 89. https://doi.org/10.3390/ani14010089
Jo Y, Lee SJ, Bia MM, Choe S, Jeong D-H. First Report of Sarcocystis pilosa from a Red Fox (Vulpes vulpes) Released for the Re-Introduction Project in South Korea. Animals. 2024; 14(1):89. https://doi.org/10.3390/ani14010089
Chicago/Turabian StyleJo, Yeonghoon, Sook Jin Lee, Mohammed Mebarek Bia, Seongjun Choe, and Dong-Hyuk Jeong. 2024. "First Report of Sarcocystis pilosa from a Red Fox (Vulpes vulpes) Released for the Re-Introduction Project in South Korea" Animals 14, no. 1: 89. https://doi.org/10.3390/ani14010089
APA StyleJo, Y., Lee, S. J., Bia, M. M., Choe, S., & Jeong, D.-H. (2024). First Report of Sarcocystis pilosa from a Red Fox (Vulpes vulpes) Released for the Re-Introduction Project in South Korea. Animals, 14(1), 89. https://doi.org/10.3390/ani14010089