Simple Summary
The Pacific blue shrimp (Litopenaeus stylirostris) is essential to the global seafood market, but intensive farming has increased disease and reduced genetic diversity. This study aimed to develop microsatellite markers for broodstock management using high-throughput transcriptome sequencing from various tissues. We identified 40 robust markers, revealing significant genetic diversity crucial for sustainable shrimp farming. These markers are valuable for breeding programs and help in the conservation and improvement of shrimp populations.
Abstract
The Pacific blue shrimp (Litopenaeus stylirostris) is a premium product in the international seafood market. However, intensified farming has increased disease incidence and reduced genetic diversity. In this study, we developed a transcriptome database for L. stylirostris and mined microsatellite markers to analyze their genetic diversity. Using the Illumina HiSeq 4000 platform, we identified 53,263 unigenes from muscle, hepatopancreas, the intestine, and lymphoid tissues. Microsatellite analysis identified 36,415 markers from 18,657 unigenes, predominantly dinucleotide repeats. Functional annotation highlighted key disease resistance pathways and enriched categories. The screening and PCR testing of 42 transcriptome-based and 58 literature-based markers identified 40 with successful amplification. The genotyping of 200 broodstock samples revealed that Na, Ho, He, PIC, and FIS values were 3, 0.54 ± 0.05, 0.43 ± 0.09, 0.41 ± 0.22, and 0.17 ± 0.27, respectively, indicating moderate genetic variability and significant inbreeding. Four universal microsatellite markers (CL1472.Contig13, CL517.Contig2, Unigene5692, and Unigene7147) were identified for precise diversity analysis in Pacific blue, Pacific white (Litopenaeus vannamei), and black tiger shrimps (Penaeus monodon). The transcriptome database supports the development of markers and functional gene analysis for selective breeding programs. Our findings underscore the need for an appropriate genetic management system to mitigate inbreeding depression, reduce disease susceptibility, and preserve genetic diversity in farmed shrimp populations.
1. Introduction
The sustainability of aquaculture has long been challenged by human factors, such as genetic breeding, feed nutrition, disease, water quality, and climate change [1]. The high fertility and early mortality of marine crustaceans can affect the stability of alleles in the population, thereby altering the genetic polymorphisms and structure of the natural population [2]. Over numerous breeding generations, populations exhibiting robust vigor and significant traits give rise to distinct genotypes, leading to a reduction in evolutionary plasticity [3] or the loss of genetic polymorphism caused by mutations or genetic drift [4]. The Convention on Biological Diversity has been closely focusing on the conservation of genetic diversity in both wild and farmed species, along with the development of effective methods for measuring it [5]. Therefore, the sustainable development of aquaculture also requires preserving genetic diversity in natural populations and maintaining genetic variability in farmed populations [6,7].
The Pacific blue shrimp (Litopenaeus stylirostris, Penaeidae) [8], also known as blue shrimp or western blue shrimp, is an economically valuable shrimp species with breeding potential [9]. It has gained popularity in foreign markets owing to its attractive flavor and high nutritional value [10,11]. Furthermore, L. stylirostris offers the advantages of rapid growth, disease resistance, wide salinity tolerance, high-temperature tolerance, low dissolved oxygen tolerance, and low food protein requirements. L. stylirostris are mainly distributed along the Pacific coasts of Latin America and Mexico, and their biological characteristics are similar to those of L.vannamei.
Pacific blue shrimp aquaculture not only meets the needs of consumers through shrimp farming but also reduces the potential fishing pressure on wild populations through stock enhancement [12,13]. However, environmental pollution and overharvesting have gradually reduced the number of natural blue shrimp populations [13]. This is evident from the increase in artificially bred shrimp populations, as demonstrated by the breeding data of shrimp seedlings in recent years. However, there is no systematic genetic management of breeding shrimps, and there is a lack of selection and breeding based on the body size and environmental adaptability of shrimps. This leads to varying body size and increased susceptibility to various diseases [14], such as yellow head disease [15], black gill disease, soft shell syndrome, and white muscle disease, which gradually develop during growth. Furthermore, the phenotypic depression of heritable traits in inbred species has led to a decline in the economic traits and production efficiency, resulting in serious economic losses to the shrimp industry [12,13]. Whether these problems arise from gene erosion in germplasms, or a lack of genetic management remains unclear [16,17]. Therefore, identifying the molecular markers to explore the genetic diversity and the strategic development of the genetic breeding management of this species is important [18].
The development of sequencing technologies, particularly next-generation sequencing (NGS), has revolutionized genomic research by generating vast amounts of transcriptome data across various species [19,20]. This advancement has facilitated the rapid identification of genetic markers, including microsatellites and genomic variations [21,22]. Importantly, NGS has enabled the annotation of functional genes and their associated traits, deepening our understanding of population genetics and evolutionary biology [23,24]. Transcriptome sequencing not only provides valuable insights into gene expression profiles but also allows for the annotation of functional genes, shedding light on the molecular mechanisms underlying various biological processes, including disease resistance in shrimp populations. Furthermore, the identification of microsatellites from transcriptome data offers a powerful tool for genetic analysis and management in aquaculture. These markers can be utilized to assess genetic diversity, population structure, and relatedness among individuals, aiding in the development of effective breeding strategies and conservation efforts [25,26].
Microsatellite markers are simple sequence repeats of highly polymorphic and codominant DNA motifs (n could range from 8 to 50) that are widely distributed throughout the genomes of eukaryotes and are widely used for different population genetic studies in many different species, such as genetic diversity [27,28,29], population structure, parentage determination [30,31,32], and constructing genetic linkage maps [33,34,35,36,37], animal identification, meat traceability [38,39] and animal breeding [40,41]. Studies on related aquatic species with highly duplicated genomes have demonstrated that microsatellites derived from transcriptomes are significantly more effective than those developed from the genome using traditional methods [42]. However, genetic studies on microsatellite markers and their related functions derived from next-generation transcriptome sequencing are limited. Furthermore, a few studies on other members of Palaemonidae, including Macrobrachium nipponense [23], Macrobrachium rosenbergii [43], and Palaemonetes sinensis [34], have explored the applicability of microsatellite markers in understanding the genetic diversity in these species. However, to date, no studies have been conducted on the genetic background of L. stylirostris-farmed populations using molecular markers related to functional genes in the transcriptome. We hypothesized that developing transcriptome-based microsatellite markers can help determine the possible reasons for the reduction in genetic diversity in the germplasm or gene weakening in L. stylirostris.
Therefore, the present study aimed to investigate the variation in different genotypes of transcriptome-derived microsatellite markers and analyze them in different cultured Pacific blue shrimp populations (different genetic backgrounds) using molecular marker-assisted selection to (i) rapidly identify whether the germplasm has gene-weakening problems and (ii) verify the applicability of the markers.
2. Materials and Methods
2.1. Experimental Animals
Pacific blue shrimps were collected from an experimental shrimp hatchery at the Golden Broodstock Development Center of the Golden Corporation Sendirian Berhad, Kampung Serambangun, Brunei, in July 2019. A total of 200 samples were obtained, comprising 100 male and 100 female shrimps from the broodstock population of the Pacific blue shrimp. This comprehensive sampling approach ensured a balanced representation of both sexes, allowing for a robust analysis of genetic diversity within the population. Detailed information on the size and growth stages of the shrimp was recorded. The average body weight was approximately 10.56 ± 0.26 g, and the body length ranged from 8 to 10 cm, categorizing them in the subadult stage. The subadult stage was selected because the shrimp were in a period of rapid growth, with high nutritional needs and stringent requirements for water quality and feed. This stage provides data that more accurately reflect current aquaculture conditions, offering valuable insights for future research.
In light of the challenges involved in acquiring wild Pacific blue shrimp specimens, we followed the methods outlined [44] and opted to utilize commercially available frozen Pacific white shrimp (L. vannamei) and black tiger shrimp (P. monodon) as control samples. Thirty samples of commercially available fresh aquatic products of Pacific white shrimp were purchased from the Taiwan supermarket in December 2021. The samples were obtained from Fuguo Freezing Co., Ltd. in Yilan, Taiwan (produced in Taiwan), Charoen Pokphand Enterprise (Taiwan) Co., Ltd. in Taipei, Taiwan (produced in Thailand), and Lequality Life Co., Ltd. in Kaohsiung, Taiwan (produced in Nicaragua). The samples were labeled as PWS-TW (n = 10), PWS-TH (n = 10), and PWS-NI (n = 10), respectively. Thirty samples of commercially available fresh aquatic products of black tiger shrimp were purchased from the Taiwan supermarket in December 2021. The samples were obtained from Sanheng International Trading Co., Ltd. in New Taipei, Taiwan (produced in Malaysia), Shangyi Fishery Co., Ltd. in Taichung, Taiwan (produced in Malaysia), and First Flight Frozen Food Co., Ltd. in Kaohsiung, Taiwan (produced in Vietnam). The samples were labeled as BTS-MY1 (n = 10), BTS-MY2 (n = 10), and BTS-VN (n = 10), respectively (Figure A1). Tissue samples were stored at −80 °C until further RNA extractions were made.
To ensure that commercially available shrimp samples met the experimental requirements, several key evaluation steps were undertaken. First, the sex ratio was examined to ensure representativeness, involving visual inspections and random sampling of 10% of each batch to achieve an approximately 1:1 male-to-female ratio. Secondly, detailed data on body size and growth stages were recorded by measuring length and weight to determine appropriate developmental stages for the experiment. Third, health assessments included both external and internal inspections for signs of damage, disease, or parasites to ensure reliable results. Fourth, genetic diversity was assessed using known microsatellite markers to ensure sufficient genetic variation. Sample size and quality control measures, including repeated testing, were verified to ensure data reliability and consistency. Finally, the experimental design included control groups from different sources and growth stages to ensure result comparability, along with multiple replicates to validate the findings. Through these comprehensive evaluations, we ensured that the commercially available Pacific white shrimp and black tiger shrimp samples met all experimental requirements, guaranteeing reliable and valid research outcomes.
Each type of shrimp and tissue sample served specific purposes in our study: Pacific blue shrimp (L. stylirostris) samples were collected to establish a comprehensive genetic repository and analyze the genetic diversity within the broodstock population. Various tissues, including the muscle (MU), hepatopancreas (HE), lymphoid organ (LY), and intestinal tract (IN), were collected to capture a wide range of genetic information. Pacific white shrimp (L. vannamei) control samples were used as a genetic control to compare and validate the genetic markers identified in Pacific blue shrimp. These samples were obtained from different regions and labeled as follows: Taiwan (PWS-TW), Thailand (PWS-TH), and Nicaragua (PWS-NI). Black Tiger Shrimp (P. monodon) control samples provided additional genetic control and facilitated cross-species comparisons of the identified genetic markers. Fresh aquatic product samples were sourced from commercial suppliers. Samples were sourced from Malaysia (BTS-MY1), Malaysia (BTS-MY2), and Vietnam (BTS-VN), ensuring a comparable and robust genetic analysis. This detailed sampling strategy, along with the use of control samples from related shrimp species, enabled us to perform a robust genetic diversity analysis, ensuring the reliability and validity of our findings.
2.2. Total RNA Extraction and Purification
Instruments required for the experiment, such as forceps, scissors, and tissue-grinding rods, were soaked in 3% H2O2 overnight and then sterilized and dried at 121 °C for 20 min using a wet heat sterilization method. The muscle (MU), hepatopancreas (HE), lymphoid (LY), and intestinal tract (IN) tissues were collected from 3-month-old Pacific blue shrimp, placed in 1.5 mL microcentrifuge tubes containing 500 µL of RNA laterTM (#2724011, Invitrogen, Vilnius, Lithuania), and stored at −20 °C until use. The tissues were later removed, and the RNA laterTM was pressed out. Total RNA was extracted using the TRIzol® (Invitrogen, Carlsbad, CA, USA) reagent. Tissue was weighed and homogenized in a microcentrifuge tube with a tissue-grinding rod, Bio Vortezer Mixer 1083-MC (BioSpec, Bartlesville, OK, USA), at a ratio of 100 mg/mL TRIzol® (50 mg/mL for HE) and incubated for 5 min. To this, 0.2 mL/mL of TRIzol® chloroform was added, shaken vigorously for 15 s, and incubated at room temperature for 3 min. The solution was then centrifuged at 12,000× g, 4 °C for 15 min, and the obtained supernatant was transferred to another microcentrifuge tube. To the solution, 0.5 mL of isopropanol (J.T Baker, Philipsburg, NJ, USA)/mL TRIzol® was added, mixed well by shaking, and incubated at room temperature for 10 min. The solution was then placed in a freezer overnight at −20 °C and centrifuged at 12,000× g, 4 °C for 10 min to obtain a white precipitate. The supernatant was removed, and the precipitate was washed with 1 mL of 75% EtOH/mL TRIzol®, shaken, and centrifuged at 7500× g, 4 °C for 5 min. The supernatant was removed, and the precipitate was air-dried in a 55 °C incubator for 2–3 min. DEPC (diethyl pyrocarbonate)-treated 2dH2O was added to the precipitate, and the solution was heated at 55 °C for 15 min to dissolve the RNA. The total RNA concentration was measured using a NanoDropTM 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) by measuring optical density OD260 and OD280. The RNA quality was checked by means of electrophoresis using fresh 1% agarose gel placed in the Muoid-2plus horizontal electrophoresis system (Advance, Tokyo, Japan), followed by staining with ethidium bromide for 10 min. DNA was removed using the DNase I, Amplification Grade (Invitrogen, Carlsbad, CA, USA). For DNase I digestion, 1 μg RNA, a 1 µL 10× DNase I Reaction Buffer, and 1 μL DNase I, AmpGrade (1 U/µL) were mixed with DEPC-treated 2dH2O to create a total volume of 10 μL, and the mixture was reacted at room temperature for 15 min on ice. Following that, 1 µL of 25 mM EDTA was added, and the solution was incubated at 65 °C for 10 min. The purified total RNA was stored at −80 °C for future use. Finally, the total RNAs obtained from the same organs of three individuals were pooled into separate tubes. Approximately 2 µg of total RNA was used to construct 14 high-quality cDNA libraries comprising 6 MU, 2 HE, 4 LY, and 2 IN samples. This pooling strategy was employed to reduce biological variability and achieve a more robust representation of the transcriptome for each tissue type. The different numbers of cDNA libraries for each tissue type were based on strategic decisions considering multiple factors, including the ease of obtaining high-quality RNA and the biological significance of each tissue type.
2.3. Establishment of Transcriptome Database and Annotation of Sequencing
To establish the transcriptome database, we employed the high-throughput sequencing platform Illumina HiSeq 4000 NGS system (Illumina, San Diego, CA, USA), which enabled the generation of large-scale transcriptome data. Clean reads were obtained after removing the reads with low quality, joint contamination, and a high N content of unknown bases in the original raw data. These were then used to calculate Q20, Q30, GC content, and sequence length. Subsequently, they were assembled de novo using Trinity V.2.0.6 [45]; noise from the assembled transcriptome was removed using the TGI Clustering tools (TGICL) [46], and the reads were sorted and screened to obtain unigenes. The obtained unigenes were divided into Clusters (representing a collection of unigenes with a similarity greater than 70%; the data names start with CL) and Singletons (representing individual unigenes; the data names start with unigenes). Finally, BLAST [47] was used to annotate the unigenes in the NT, NR, Clusters of Orthologous Groups (COG), Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and Swiss-Prot functional databases.
2.4. Genomic DNA Extraction
Approximately 0.06 ± 0.01 g of shrimp muscle tissue was placed in a 1.5 mL microcentrifuge tube, and 600 µL of 70% ethanol was added to it. The mixture was then stored at −20 °C until genomic DNA extraction. During DNA extraction, ethanol was discarded, and the tissue samples were washed twice with a 300 µL DE buffer (1 M Tris, 0.5 M EDTA, 10% SDS, and 2dH2O; pH 8). Afterward, a 600 µL DE buffer and 2 µL of proteinase K (Gene Mark, Taipei, Taiwan) were added to the tube, mixed evenly, and incubated in a rotary shaker (Evernew, Yu-Shing Biotech. Ltd., Taipei, Taiwan) at 55 °C and 80 rpm for 2–6 h until the tissue samples were completely dissolved. Following that, 500 µL of the sample was pipetted into a new 1.5 mL centrifuge tube, 300 µL of MPC Protein Precipitation Reagent (Epicentre Technologies Co., Madison, WI, USA) was added to it, and the contents were invert-mixed for approximately 2 min to precipitate the protein, and centrifuged at 10,000× g and 4 °C in a micro-refrigerated high-speed centrifuge for 10 min. Subsequently, 600 µL of the supernatant was transferred to a new 1.5 mL centrifuge tube, 600 µL of isopropanol (Sigma Chemical Co., St. Louis, MO, USA) was added to it, mixed for 20–40 min using a 3D rotary shaker (B3D1320, Benchmark Scientific, Sayreville, NJ, USA), and centrifuged at 4 °C and 10,000× g for 10 min in a micro-refrigerated high-speed centrifuge. After discarding the supernatant, 200 µL of 70% ethanol was added to wash the tube wall, and the mixture was allowed to stand for 10 min. After removing the alcohol, the pellet was washed using 200 µL of 70% ethanol and centrifuged at room temperature for 10 s using a mini again microcentrifuge (C1008, Benchmark Scientific, Sayreville, NJ, USA) to remove the alcohol in the tube completely before being oven-dried at a constant temperature of 55 °C. The pellet was then resuspended in 50 µL of 2dH2O and kept in a water bath maintained at 37 °C for 4–8 h until complete dissolution.
The concentration and absorbance of the DNA samples were measured using a NanoDropTM One spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) by recording the OD260 and OD280 values. The DNA quality was assessed by means of electrophoresis separation on a 0.8% agarose gel and stained with a GelRed Nucleic Acid Gel Stain (Biotium Inc., Fremont, CA, USA) for 20 min. The results were visualized using the Slite140 Fluorescent Photoresist System (Avegene Life Sciences, Taipei, Taiwan). The DNA samples were diluted to a working concentration of 25 ng/µL and stored at −20 °C until further use.
2.5. Genetic Diversity Analysis
Microsatellite markers were searched in the Pacific blue shrimp transcriptome dataset and NCBI database [48,49,50], and a common microsatellite marker for Penaeus was designed using Geneious V.9.1.8 software (https://www.geneious.com accessed on 3 April 2022). The primers were designed using the following criteria: predicted product size, 100–500 bp; GC content, 40–60%; average Tm, 52–62 °C; the self-adhesive area of the primer was avoided; and a self-adhesive temperature ≤37 °C. The 5′ ends of the forward primers of all microsatellite markers were connected with fluorescent primer adapters for the second PCR amplification. Specifically, markers focusing on the molecular markers involved in functional genes related to shrimp disease resistance traits were identified from the transcriptome dataset.
The first PCR amplification was carried out in a reaction volume of 10 µL comprising a 5 µL 2× Taq DNA polymerase Mix (Bioman, Taipei, Taiwan), 0.3 µL forward primer, 0.3 µL reverse primer, 2.4 µL 2dH2O, and 2 µL template DNA. PCR was performed using a 96-well polymerase chain reactor Veriti® thermal cycler or SimpliAMPTM thermal cycler (both from Applied Biosystems, Foster City, CA, USA) and the following reaction conditions: 94 °C for 5 min; 30 cycles of 94 °C for 40 s, Tm °C for 30 s, and 72 °C for 40 s; 72 °C for 8 min; and maintaining the temperature at 4 °C. The annealing temperatures (Tm) varied according to the primer and ranged from 56 °C to 60 °C for the Pacific blue shrimp transcriptome dataset and from 52 °C to 60 °C for the NCBI database.
For the second PCR amplification, the original forward primer was replaced with a fluorescent primer tagged with FAM (blue), ROX (red), NED (yellow), or JOE (green) dyes. The reaction mix (10 µL) comprised the same components as those in the first PCR amplification, except for the replacement of the forward primer and template DNA with 0.3 µL of the fluorescent primer and 2 µL of the first PCR product diluted tenfold, respectively. PCR was performed using the same conditions and PCR systems as the first PCR amplification. Finally, the second PCR products were separated by means of electrophoresis on 2% agarose gel, stained with the GelRed® Nucleic Acid Gel Stain for 25–30 min, and photographed using a Slite140 Compact Gel Documentation System (Avegene Life Sciences, Taipei, Taiwan).
Subsequently, 5 µL of each of the four fluorescently labeled PCR products were pooled with 0.2 µL GeneScanTM 600 LIZTM size standard and 10.8 µL HiDiTM deionized formamide (both from Applied Biosystems, Foster City, CA, USA) added to them, denatured at 94 °C for 3 min, and immediately transferred to ice for 5 min. The samples were then analyzed using an ABI PRISM® 3730xl DNA Analyzer (Applied Biosystems, Foster City, CA, USA) to separate short tandem repeats (STRs). The data were analyzed and interpreted using Geneious V9.1.8 software, and the results were exported to Cervus 3.0.7 (http://www.fieldgenetics.com/pages/home.jsp accessed on 3 April 2022) to calculate the following genetic parameters: the mean number of alleles (Na), sum of all alleles in the population, average observed heterozygosity (Ho), actual proportion of heterozygous individuals contained in each locus in the population; the average expected heterozygosity (He); and estimated heterozygosity in the population according to the Hardy–Weinberg equilibrium [51]. The expected value of the degree of convergence was calculated along with two key parameters used to assess genetic diversity and population structure: polymorphic information content (PIC), which represents the amount of polymorphic information in linkage analysis [52], and the gene fixation index (FIS), which measures the degrees of inbreeding between individuals within the population.
2.6. Development of Universal Microsatellite Markers
We designed 30 functional primer sets from our dataset, of which 15 primer sets were successfully amplified. In addition, we obtained 25 sets of primers from the Blue Shrimp Genetic Diversity Test panel that showed amplification under the set conditions of temperature testing. Finally, 40 primer pairs were evaluated to identify the universal microsatellite markers for shrimp.
The samples of L. stylirostris, L. vannamei, and P. monodon were categorized into the following five groups: (1) PBS-8, comprising a group of eight individuals including L. stylirostris; (2) PWS-1, comprising a single individual of L. vannamei; (3) PWS-8, comprising a group of eight individuals including L. vannamei; (4) BTS-1, comprising a single individual of P. monodon; and (5) BTS-8, comprising a group of eight individuals including P. monodon. Among these, eight shrimps of L. vannamei and P. monodon were sourced from three different origins. L. stylirostris was selected for the polymorphism analysis from genetically diverse species with different genotypes.
The PCR was performed in a reaction mixture of 10 µL comprising a 5 µL 2× Taq DNA polymerase Mix (Bioman, Taipei, Taiwan), 0.3 µL forward primer, 0.3 µL reverse primer, 2.4 µL 2dH2O, and 2 µL template DNA. The PCR conditions and amplification system were the same as those used for the first PCR amplification (described in Section 2.5).
The Tm was adjusted according to the primers, and the exact length of the marker was confirmed using capillary electrophoresis. Five microliters of the reaction solution from each group were mixed in five tubes. Geneious V9.1.8 software was used to interpret and analyze the shared shrimp microsatellite markers for the microsatellite marker data. After the analysis of NGS data, amplified and diverse microsatellite markers were used for STR verification. The PCRs were performed in two steps: first and second PCR amplifications and the data were analyzed and interpreted as described in Section 2.5.
2.7. Statistical Analysis of Population Diversity
The obtained genetic data were imported into GenAlEx 6.502 [53] to statistically analyze the Na of each microsatellite locus, as well as various coefficients for evaluating the diversity of the population, including Ho, He, PIC, and FIS.
3. Results
3.1. High-Throughput NGS and Alignment of Reference Gene Sequences
Fourteen cDNA libraries were established from MU (n = 6), HE (n = 2), LY (n = 4), and IN (n = 2) tissues collected from L. stylirostris (n = 18). A total of 7.04 Gb of averaged data was produced, and 687.0366 M sequence reads were generated with a Q30 score of more than 92.98% (Table S1). From these data, 53,263 unigenes were assembled, with a total length of 94,147,609 bp; an average length of 1767 bp; an N50 of 3478 bp; and GC content of 44.56% (Tables S2 and S3). As shown in Figure S1, the length of the reads varied from 300 to >3000 bp, with most unigenes (n = 9999) being 300 bp.
The functional annotation of the identified unigenes against publicly available databases is shown in Table S4. A total of 24,066 unigenes were annotated in at least one functional database (Table S4), wherein 3711 unigenes were co-annotated in all databases (Figure 1).
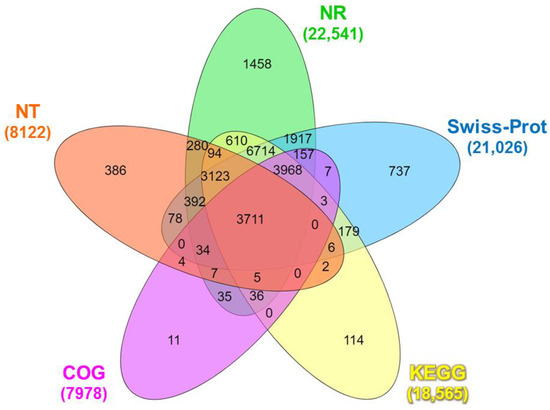
Figure 1.
Venn diagram of the number of unigenes of Litopenaeus stylirostris functionally annotated in five major databases: NR, NT, COG, Swiss-Prot, and KEGG. NR: Unigenes with NCBI non-redundant protein; NT: nucleotide database; COG: Clusters of Orthologous Groups; Swiss-Prot: A curated protein sequence database that strives to provide high levels of annotation; KEGG: Kyoto Encyclopedia of Genes and Genomes.
The Venn diagram analysis revealed that a total of 3711 unigenes were co-annotated in all databases (Figure 1). The BLASTx search against the NR protein database revealed substantial similarity between the unigene sequences and diverse organisms. The top matches included Nevada termite (Zootermopsis nevadensis; 12.99%), Daphnia pulex (Daphnia pulex; 6.92%), Atlantic horseshoe crab (Limulus polyphemus; 6.39%), Pacific white shrimp (L. vannamei; 3.69%), black tiger shrimp (P. monodon; 2.12%), and Pacific blue shrimp (L. stylirostris; 0.10%). The remaining matches accounted for 67.89% of the total hits (Figure S2). The BLASTn search of the unigenes sequences against the NT database identified 23,635 coding sequence (CDS) regions (p < 0.0001), while ESTScan predicted 7006 CDS regions (Table S5).
3.2. COG, GO, and KEGG Annotation Results
The possible positions of the predicted CDS unigenes in the system classification were analyzed through the prediction and possible functional classification of the unigenes in the COG database. A total of 7978 unigenes, derived from combined transcriptome data of the muscle (MU), hepatopancreas (HE), lymphoid (LY), and intestine (IN) tissues, were subdivided into 24 functional categories in the COG database. The largest category annotated with the most unigenes (n = 2187) was “general function prediction only”, followed by “post-translational modification, protein turnover, and chaperones” (n = 1046) (Figure 2).
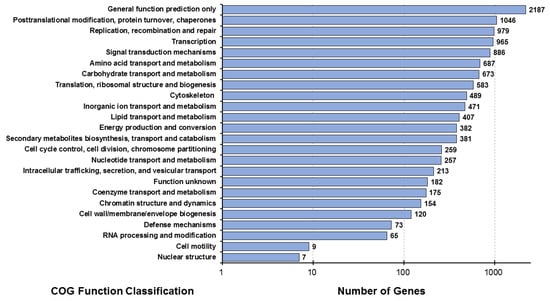
Figure 2.
Distribution map of COG functional annotations of Litopenaeus stylirostris. The abscissa represents the number of unigenes corresponding to the classification, while the ordinate represents the COG functional classification. COG, Clusters of Orthologous Groups.
A total of 8828 unigenes were annotated in the GO database. In the biological process category, “cellular component organization and or biogenesis” was the most annotated term, with 4294 unigenes, followed by the “metabolic process”, with 3712 unigenes, and “single-organism process”, with 3027 unigenes. In the cellular component category, most unigenes were annotated to the term “cell” (n = 2978), followed by “cell parts” (n = 2953) and “membrane” (n = 2160). In the molecular function category, “binding” was the most annotated term with 4283 unigenes, followed by “catalytic activity” (n = 3954) and “transporter activity” (n = 525) (Figure 3).
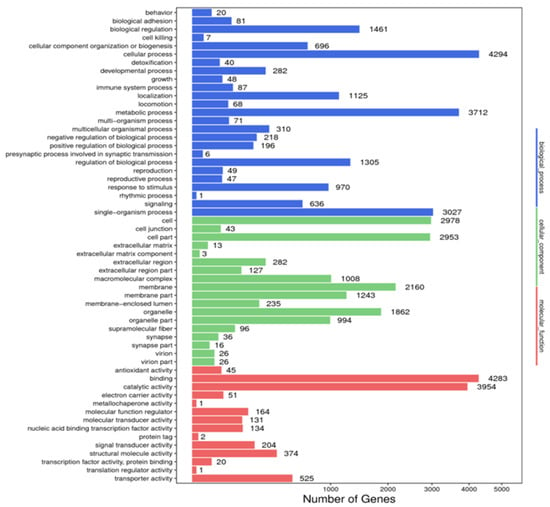
Figure 3.
Annotated map of GO function of Litopenaeus stylirostris. The abscissa represents the number of unigenes in the corresponding category, while the ordinate represents the annotated terms in different GO categories. According to the NR annotation results, the unigene GO annotation information was obtained using BLAST2GO software (https://www.blast2go.com/, accessed on 28 May 2022). To preliminarily understand the gene function distribution of L. stylirostris, the sequenced unigenes, and assembled transcripts were compared with the GO database, and unigenes were mapped to different functional categories after predicting their possible functions. GO, Gene Ontology.
Annotation of the unigenes with the KEGG database mapped 18,565 unigenes to 42 pathways, among which “signal transduction” was the most frequently annotated pathway, with 2981 unigenes, followed by “global and overview maps”, with 2587 unigenes, and “cancers: overview”, with 2336 unigenes. Other predominant pathways included “transport and catalyst” (n = 1805) and “cellular community” (n = 1700) (Figure 4).
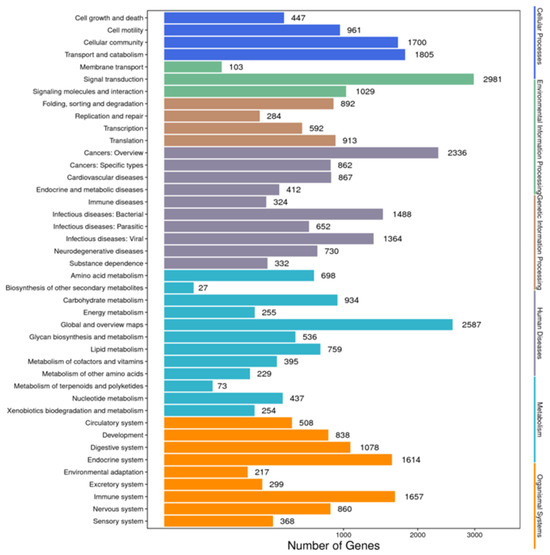
Figure 4.
Functional annotations of the unigenes of Litopenaeus stylirostris using the KEGG pathway database. We predicted the potential functions of the unigenes in biochemical pathways and performed functional classification statistics to understand the overall distribution of gene functions in L. stylirostris. The x-axis represents the number of unigenes in each corresponding category, while the y-axis displays the functional subcategories of the KEGG pathway database. The six functional categories in the KEGG pathway database—cellular processes, environmental information processing, genetic information processing, human diseases, metabolism, and organismal systems—are represented on the right-hand y-axis. KEGG, Kyoto Encyclopedia of Genes and Genomes.
3.3. Microsatellite DNA Marker Detection
The annotation of the unigenes using BLAST and ESTScan identified 30,641 CDSs. Among these, 36,415 microsatellites were detected in 18,657 unigenes using the MISA software (https://webblast.ipk-gatersleben.de/misa/ accessed on 8 June 2022) [54] based on the assembled unigene as a reference sequence. We identified 7400 single-, 17,735 double-, 8844 triple-, 1296 four-, 327 five-, and 813 six-base repeats (Figure 5, Table 1) through GO annotation, KEGG annotation, pathway enrichment analysis, locus detection software, function classification, microsatellite variation sequence, and other biological information analyses. Based on the predicted microsatellite position on the genomic DNA sequence (CDS), we identified 30 functional genes from the L. stylirostris transcriptome to develop candidate molecular markers (Table A1 and Table S6).
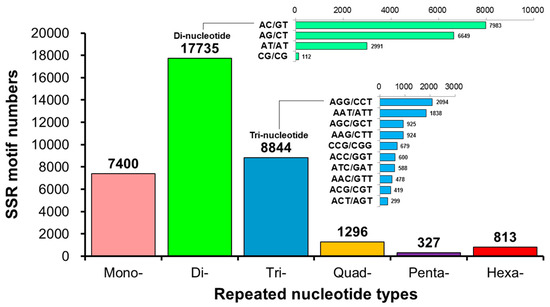
Figure 5.
A statistical map of the distribution of microsatellite marker (or simple sequence repeat; SSR) length in Litopenaeus stylirostris. The horizontal axis displays the length of the repeat unit and specific repeat type, while the vertical axis represents the number of microsatellites. ‘Number of Repeats’ refers to the length of the repeated bases; ‘Mono-Nucleotide Repeats’, ‘Di-Nucleotide Repeats’, ‘Tri-Nucleotide Repeats’, ‘Quad-Nucleotide Repeats’, ‘Penta-Nucleotide Repeats’, and ‘Hexa-Nucleotide Repeats’ indicate the number of single-, double-, triple-, four-, five-, and six-base repeats, respectively.

Table 1.
Statistics of the number of simple sequence repeat (SSR) marker types of transcripts in Litopenaeus stylirostris.
3.4. Genetic Diversity Analysis of Pacific Blue Shrimp Broodstock
A total of 100 microsatellite markers, including 42 designed from transcriptomes (Type I microsatellite markers; Table A1 and Table S6) and 58 obtained from previous studies on the diversity of Litopenaeus populations (Type II microsatellite markers; Table S7), were used to study the genetic diversity of the cultured L. stylirostris species in Brunei. After the PCR temperature test, 22 sets (52.38%) of Type I markers and 18 sets (31.03%) of Type II markers were successfully amplified (Figure S3).
An analysis of the broodstock population of 200 L. stylirostris revealed that three DNA markers were polymorphic (CL1472.Contig9, LV13, and LV29), which could be stably amplified in the population of L. stylirostris and analyzed using STRs. Among the remaining 37 markers, CNM4 had a single-base slip, which was inconsistent with the predicted length of the microsatellite sequence, and the other 36 markers were monomorphic, exhibiting only one allele. The alleles detected by each marker were expressed in the order A, B, C, etc., according to the alleles of different fragment sizes and lengths. Among them, LV13 detected the most alleles (n = 4 alleles), wherein both LV13 and CL1472.Contig9 yielded four genotypes (Table 2).
Table 2.
Allele and genotype analysis of polymorphic microsatellite markers in Litopenaeus stylirostris populations 1.
Next, we used these three markers (CL1472.Contig9, LV13, and LV29) to assess genetic variation in the L. stylirostris population. The Ho values for these markers were 0.600, 0.505, and 0.513, respectively, with an average of 0.54 ± 0.05, while He values were 0.537, 0.375, and 0.390, respectively, with an average of 0.43 ± 0.09. The average PIC and FIS values of these three markers were 0.41 ± 0.22 (0.333, 0.246, and 0.663, respectively) and 0.17 ± 0.27 (0.315, 0.339, and −0.135, respectively). The average PIC value indicated that this population was moderately polymorphic (0.25 < PIC < 0.5), whereas the positive average FIS value suggested that the Pacific blue shrimp broodstock lacked genetic diversity and was in a state of inbreeding. The CL1472.Contig9 locus showed a negative FIS value, indicating that this locus presents more heterozygotes. Therefore, it is necessary to supplement the stock with new seedlings to maintain the genetic polymorphism (Table 2).
3.5. Development of Universal Microsatellites for Shrimp
Forty markers were bonded to the DNA of the three shrimp species using PCR, and NGS was performed after grouping and mixing to analyze the bonding effects of the markers and the three shrimp species (Figure 6). In PBS-8, 37 markers were successfully amplified, but only 2 markers, CNM13 and LV13, showed diversity, while the rest amplified only one or two alleles. In PWS-1, 35 markers were successfully amplified, of which 3 were not specific enough; the amplified sequences were less and inconsistent, so they were not included in the follow-up STR verification analysis (Table A2). In PWS-8, among the 31 markers that were stably amplified, 21 showed gene diversity, and the remaining 10 resulted in only one or two genotypes. In BTS-1, 18 of the 28 successfully amplified markers showed insufficient specificity and less and inconsistent amplified sequences; therefore, they were excluded from the follow-up STR verification analysis. In BTS-8, 4 of the 10 stably amplified markers showed gene diversity, and the remaining 6 amplified only one or two alleles; these 4 markers also showed diversity in L. vannamei. Although there were still three markers that failed to amplify stably or showed poor adhesion efficiency in L. stylirostris species, the amplified detection ratio was still 92.5%. Upon calculating the expansion success rate of the other two penaeid shrimp species, the proportions tested for successful amplification in L. vannamei and P. monodon were 77.5% and 25%, respectively.
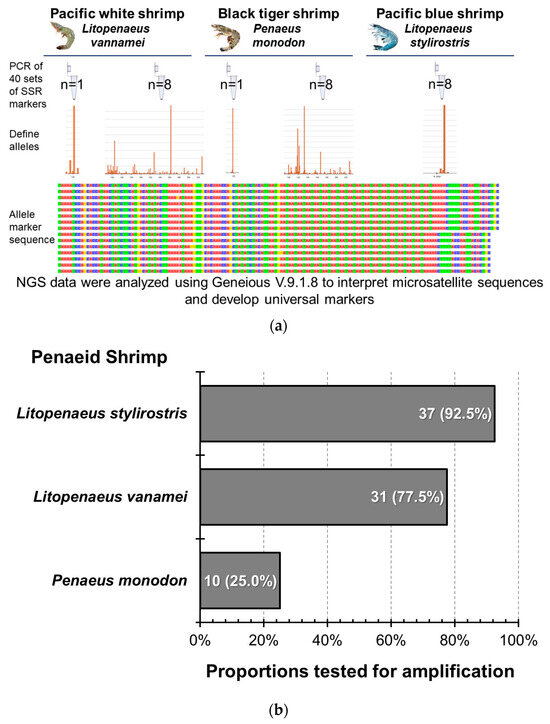
Figure 6.
The amplification efficiency of microsatellite markers shared by different penaeid shrimp species. (a) Diagram of the total marker testing methodology using 40 microsatellite markers. (b) The successful amplification rate of the 40 markers in different Penaeid species. The colors in the allele marker sequence represent different bases: red for adenine (A), green for thymine (T), blue for cytosine (C), and yellow for guanine (G).
Based on the results of the shrimp and marker amplification mentioned above, 17 microsatellite markers were successfully amplified in Pacific blue and white shrimps. Pacific white shrimps exhibited diversity, with four microsatellite markers successfully associated with Pacific blue, Pacific white, and black tiger shrimps, indicating stable expansion and diversity. To confirm whether the above markers could be used to analyze related genetic parameters, using techniques such as STR genotyping and diversity analyses, each marker was verified via STR analysis in shrimps. Based on the above amplification results, the STRs were compared with those of the shrimps in PBS-8, PWS-1, PWS-8, BTS-1, and BTS-8 groups; these were then bonded with PCR fluorescent primers and sequenced.
Among the 17 markers stably amplified from the two types of shrimps, 7 markers (CNM1, CNM8, CNM12, CNM22, LV29, CL4833.Contig1, and Unigene6789) yielded consistent results with the expected fragment lengths. Although the remaining 10 markers had stable fluorescent signals, the length of the fluorescent signal fragments did not match the expected fragment length. A comparison of the fluorescence sequencing results of these 10 markers with the results of NGS revealed that 7 markers (CNM2, CNM3, CNM4, CNM13, CNM15, CL3247.Contig3, and Unigene7536) contained the originally predicted microsatellites. However, there were single-base variations at different positions within the microsatellite sequences, resulting in no diversity as initially anticipated. It also showed that the variation in another marker (CL5798) bonded to the STRs in Pacific white shrimp was a single-base variation, and the sequence did not contain the predicted microsatellites. A comparison of the bonding part of LV3 and the STRs in Pacific white shrimp with the NGS results identified other single-base microsatellite variations in the sequence in addition to the originally predicted diversity of microsatellites. Therefore, the primer used in this study was not suitable for the future diversity analysis of the target species. A comparison of the NGS results with the bonding part of CNM23 in the Pacific white shrimp genome revealed no variation in the originally predicted three-base repeat microsatellite. In contrast, diversity was observed in a two-base repeat microsatellite within the sequence, indicating its potential suitability for the diverse analysis of shrimp; however, the target microsatellite should be changed from TTA to GT repeats. The analysis revealed that the three prawns shared the microsatellite part for CL1472.Contig13, CL517.Contig2, Unigene5692, and Unigene7147. These four markers produced stable signals in STR fluorescence analysis; Unigene5692 yielded one genotype in the three prawns, CL517.Contig2 and Unigene7147 resulted in two or more genotypes in Pacific white and black tiger shrimps and CL1472.Contig9 identified eight genotypes in Pacific white shrimps (Table 3). These four markers can be used as common primers for the population diversity analyses of Pacific blue, Pacific white, and black tiger shrimps.
Table 3.
Allelic statistics of microsatellite markers applied in penaeid shrimp.
In addition to our findings on the genetic diversity of Litopenaeus stylirostris, we identified four microsatellite markers derived from the shrimp transcriptome database. These markers, namely CL1472.Contig9 [(GAG)7], CL517.Contig2 [(AAT)8], Unigene5692 [(GAG)6], and Unigene7147 [(TGA)5] exhibited cross-species amplification across Litopenaeus stylirostris, Litopenaeus vannamei, and Penaeus monodon, making them valuable tools for genetic studies in multiple shrimp species. Furthermore, the length and quantity of alleles at these microsatellite loci could be utilized to distinguish between different shrimp species. Importantly, our data revealed a significant reduction in genetic diversity in Litopenaeus stylirostris. Each of the four microsatellite loci corresponds to functional genes involved in crucial biological processes. CL1472.Contig9 [(GAG)7] corresponds to myosin light chain kinase, CL517.Contig2 [(AAT)8] to caspase, Unigene5692 [(GAG)6] to smooth muscle-like transcript variant X3, and Unigene7147 [(TGA)5] to eukaryotic translation initiation factor 2 (Table S8). Through bioinformatics analysis, we found that all four microsatellite sequences were located within the Open Reading Frame (ORF) of their respective genes. Interestingly, three of the microsatellite loci, CL1472.Contig9 [(GAG)7], Unigene5692 [(GAG)6], and Unigene7147 [(TGA)5] were situated within the Coding DNA Sequence (CDS). Furthermore, CpG islands were discovered within the CL1472.Contig9 [(GAG)7] and Unigene5692 [(GAG)6] loci, highlighting potential regulatory elements within these genomic regions. These findings underscore the importance of microsatellite markers in assessing genetic diversity and understanding functional gene variation in L. stylirostris. By elucidating the relationship between genetic markers and functional genes, our study contributed to the field of shrimp molecular genetics and offered insights into the genetic management of shrimp populations (Figure 7).
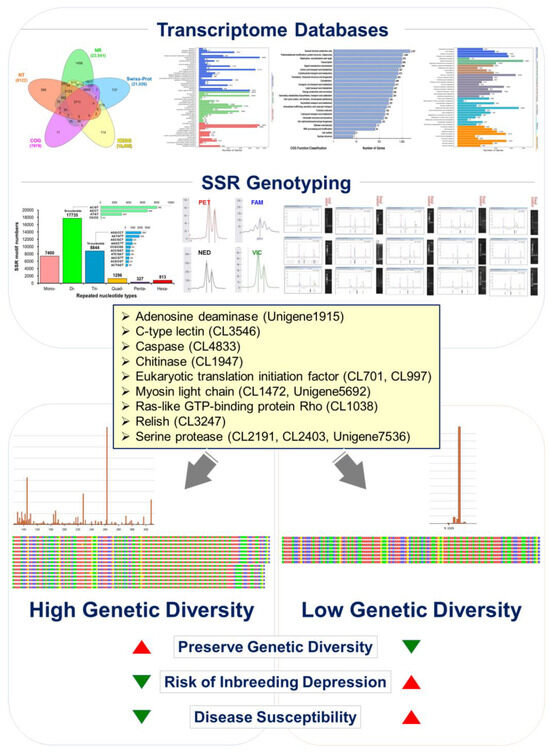
Figure 7.
Illustrating genetic management strategies to mitigate the inbreeding decline in aquaculture blue shrimp (Litopenaeus stylirostris) populations. Drawing insights from the blue shrimp transcriptome gene database, we targeted functional genes associated with White Spot Syndrome Virus (WSSV) and Acute Hepatopancreatic Necrosis Disease (AHPND) resistance. These genes include adenosine deaminase (Unigene1915), c-type lectin (CL3546), caspase (CL4833), chitinase (CL1947), eukaryotic translation initiation factor (CL701, CL997), myosin light chain (CL1472, Unigene5692), ras-like GTP-binding protein Rho (CL1038), relish (CL3247), and serine protease (CL2191, CL2403, Unigene7536). By conducting the polymorphic analysis of each functional microsatellite DNA marker genotype, our study reveals that blue shrimp populations exhibit diminished genetic polymorphism due to prolonged inbreeding and insufficient genetic management. This decline in genetic diversity contributes to inbreeding depression, impacting economically vital breeding traits and disease resistance. Implementing effective genetic management strategies for genotyping can help alleviate inbreeding depression in aquaculture populations.
4. Discussion
L. stylirostris has great potential for the development of diverse species for farming in Asia [11]. However, the primary research areas concerning this species are its taxonomy [50,55] and disease [14,56,57], while research on the genetic management of germplasm conservation, selection, and breeding, as well as on the development of genetic polymorphism monitoring and measurement technology is limited [16,17]. To provide molecular tools for genetic studies or the identification of target aquatic species, it is essential to utilize reference marker information developed for a species over time [50] or cross-species marker sequences from closely related species. This should be followed by primer pair design and the testing of conditions [58]. However, in the case of non-mainstream species, the development of cross-species markers incurs high testing costs and is compounded by the limited availability of reference information.
In recent years, NGS has emerged as a cost-effective and efficient method for developing molecular markers in non-model species for which the genetic information available is limited and requires seed management [59,60,61,62]. The genome sequences of the members of Penaeidae, such as L. vannamei [63], P. monodon [64], and Marsupenaeus japonicus [65], were assembled using the Illumina platform. A transcriptome database can be used as a comparative reference for L. stylirostris, providing more genomic information for subsequent genetic breeding research and assisting in the development of the aquaculture industry and genetic management research on this species. This study used a high-throughput transcript sequencing platform that is widely applied to develop molecular markers. In addition to making significant progress in identifying population-scale and high-throughput functional markers, the study results have great research value.
In this study, the Illumina HiSeq NGS system was used for the first time to sequence, assemble, and annotate the transcriptomes of the MU, HE, LY, and IN tissues of the Pacific blue shrimp to establish a gene bank. The Q20 (97.40%), Q30 (93.66%), and GC (45.25–52.46%) values of each cDNA dataset were similar to those in the L. vannamei transcriptome database [66,67,68], indicating that the transcriptome was of good quality. In addition, in the distribution of species annotations in the NR database, because of the limited reference materials for L. vannamei, only 0.10% of the unigenes were annotated to L. stylirostris. In comparison, 3.69% and 2.12% of the unigenes were annotated to L. vannamei and P. monodon, respectively. The transcript sequencing data generated in this study can be used to analyze differentially expressed genes and identify key genes underlying important economic traits, such as those related to biotic and abiotic stresses. Previous studies have explored the potential of transcriptomics in identifying white spot syndrome virus in M. rosenbergii [69], neuronecrosis virus infection in seven-band grouper (Hyporthodus septemfasciatus) [70], growth-related traits in kuruma shrimp (M. japonicus) [71], and melanin deposition in the fillets of Atlantic salmon (Salmo salar) [72].
The annotation of the identified unigenes with COG, KEGG, and GO databases revealed the distribution of functional genes in L. stylirostris, identifying the potential functional genes and predicting their expression. The unigenes identified the “cellular process” (n = 4294), “cell” (n = 2978), and “binding” (n = 4283) terms in the biological processes, cellular components, and molecular function GO categories, respectively. KEGG database [73,74] annotation identified the enriched pathways in L. stylirostris. The generated data can be used to understand the interactions and functional effects of various molecular regulatory information transfers involved in the metabolism, genetic information processing, environmental information processing, various other cellular processes, and diseases of L. stylirostris. These data can also allow for follow-up research on gene function and metabolic pathways in cells and help discover the regulatory mechanisms of specific functional genes of L. stylirostris.
Among the total 53,263 unigenes in the Pacific blue shrimp transcriptomics dataset, 36,415 microsatellites were detected in 35.03% of unigenes (n = 18,657), which is lower than those detected in giant freshwater prawns (M. rosenbergii; 48.76%) [75] and banana shrimp (Fenneropenaeus merguiensis; 43.5%) [76], but higher than those detected in ridgetail white prawn (Exopalaemon carinicauda; 10.97%) [77]. However, this proportion of unigenes containing microsatellites in Pacific blue shrimp was similar to that in Pacific white shrimp (L. vannamei; 34.36%) [78] and freshwater ornamental shrimp (Neocaridina denticulata; 38.77%) [79]. On average, the microsatellites were 2.58 kbp distant, whichis a distance higher than the distances in F. merguiensis (1.02 kbp) and M. rosenbergii (0.93 kbp) but lower than those in E. carinicauda (6.6 kbp) and N. denticulate (4.95 kbp), and similar to those in L. vannamei (2.34 kbp). Among the mined SSRs, the proportion of mono- (7400, 20.32%), di- (17,735, 48.70%), and tri- (8844, 24.29%) nucleotides accounted for more than 93%. These observations are consistent with those of edible or ornamental crustacean species [75,76,77,78,79]. The two shrimp species L. stylirostris and L. vannamei showed a high degree of genetic similarity in the microsatellite structure analysis of transcripts.
The detection of polymorphisms in transcriptome-derived microsatellite markers can help us understand the genetic variation structure of functional genes and has several advantages, including high efficiency, strong transferability, and correlation with potential genes [80]. Some studies have indicated that the probability of polymorphism in transcriptome-derived microsatellites may not have a significant positive correlation with the strictness of evolutionary constraints [32,81]. However, when selecting microsatellites, the following recommendations are typically followed: (1) microsatellites that are composed of adenine (A) and thymine (T) bases, which may easily lead to sequence self-adhesion (hairpin) are avoided; (2) the primer length is optimized between 18 and 25 bp, and it is ensured that the annealing temperatures of all primers fall within ±1 °C; (3) the total length of the PCR product is ensured to be within the range of 100–300 bp, with GC content in the range of 40–60%; (4) primer–microsatellite repeat pairs with a distance of less than 20 bp are avoided [82]. In this experiment, the designs of the other primers were consistent with all the above recommendations, except that the total length of the PCR products in the case of all three sets of primers exceeded 300 bp. In addition, it has been pointed out that if cross-species amplification is to be performed, the PCR annealing temperature of DNA and primer bonding should first be adjusted for each species. The amplification primers from different species can be adjusted at different DNA bonding temperatures to identify the profile providing the clearest amplified products; the annealing temperature should be raised as much as possible to avoid non-specific amplification; most primers give the best results at 58 °C [83], which is consistent with the optimal temperature for most experimental designs. Although our study initially tested 100 sets of loci in L. stylirostris, only three markers proved effective for reliable genotyping. The low proportion of polymorphic loci may be attributed to limited genetic variation within the broodstock population or the specific genomic regions targeted by these markers. While the identified microsatellite markers provide valuable insights into blue shrimp genetic diversity and broodstock management, the full genetic variability within a population may not be captured using a limited number of loci. We acknowledge this limitation and recommend that future studies incorporate more microsatellite or single nucleotide polymorphism (SNP) markers and utilize advanced high-throughput sequencing and genotyping technologies for a comprehensive genetic assessment. By exploring a broader range of loci and including more diverse populations, future research can provide a more complete understanding of the genetic variability within and among blue shrimp populations.
In this study, among the 42 transcriptome-derived microsatellite marker primers, 22 successfully amplified the product; they exhibited a reliable peak signal pattern upon capillary electrophoresis, which was obtained through test amplification among cross-species samples of different Penaeidae members. Nine markers were identified as polymorphic, and 25 scorable fragments of 111–296 bp were obtained, with an average of 6.25 amplicons per primer and NA values ranging from 1 to 10 (mean = 3.5). Furthermore, the degree of polymorphism ranged from 50.0% to 100%, with an average of 81.52%, which is similar to that observed in previous studies investigating transcriptome-derived microsatellite markers in other crustaceans [32,79]. These nine markers could detect and present the most polymorphic information in L. vannamei, followed by P. monodon and L. stylirostris. Because samples of wild blue shrimp are not easy to obtain, white shrimp were used as a control sample with high diversity for comparison. Using 42 microsatellite markers developed from the blue shrimp transcriptome, it was observed that 9 (42.86%) loci retained diversity, although the diversity of blue shrimp was only 4.55%. In addition, 58 sets (Table S7) of polymorphic primer pairs that have been used to construct the genetic linkage map were selected from the microsatellite marker library of L. vannamei. They were identified with 80% diversity in the same species, which is in agreement with previous analyses of the same species [49,50]. However, the diversity of blue shrimp was only 11.11%, and the overall polymorphism was reduced by approximately 50.83%. The analysis revealed that the microsatellite markers CL1472.Contig13, CL517.Contig2, Unigene5692, and Unigene7147 were successfully amplified and shared among the three prawn species (Pacific blue shrimp, Pacific white shrimp, and black tiger shrimp). While these markers have demonstrated utility in differentiating between species, their effectiveness for detailed population analysis within a single species may be limited. Future studies should incorporate additional microsatellite loci or SNPs to enhance the resolution of population genetic assessments. These findings underscore the necessity for the careful selection of genetic markers tailored to specific research objectives, whether it be species identification or population genetic diversity analysis.
Insufficient genetic management in aquaculture populations can undermine their resilience, rendering them more susceptible to diseases, environmental stress, and other adverse factors. Therefore, effective genetic management is crucial to safeguard the health and adaptability of aquaculture populations. When genes associated with immunity [84,85], reproduction [86,87], environmental stress response [68,88,89,90], behavior [91], and other functions experience a loss of diversity, the population’s capacity to adapt is compromised, thereby heightening the risk of encountering various ecological pressures. The analysis of functional markers corresponds to key functional genes associated with resistance traits against White Spot Syndrome Virus (WSSV) and Acute Hepatopancreatic Necrosis Disease (AHPND) found in the transcriptome sequencing dataset. These genes include adenosine deaminase, c-type lectin [92], caspase, chitinase, eukaryotic translation initiation factor, myosin light chain, ras-like GTP-binding protein Rho, relish, and serine protease (SP), which play crucial roles in disease resistance mechanisms. Numerous studies have underscored their critical role in disease resistance mechanisms [93,94,95,96,97,98]. However, it is evident that these disease resistance-related gene markers have experienced a reduction in genetic polymorphism within the blue shrimp broodstock, likely due to recent potential inbreeding depression affecting disease resistance capabilities. Furthermore, through detailed explanations of the development of microsatellite markers and their association with functional genes, this study systematically investigated the potential factors contributing to the decline in economic disease resistance traits in Pacific blue shrimp aquaculture due to prolonged inbreeding practices and a lack of effective genetic management. This comprehensive analysis not only elucidates the scientific significance of the developed microsatellite markers but also sheds light on the genetic management challenges faced by blue shrimp aquaculture. These findings have the potential to advance research in this field and provide valuable insights for developing strategies to mitigate the adverse effects of reduced genetic diversity and inbreeding depression in shrimp populations.
Subsequent research will investigate the impact of genetic diversity on the adaptive strategies of Pacific blue shrimp populations in response to environmental changes and new challenges. Furthermore, previous studies have documented the vertical transmission of potential microbial communities from shrimp eggs and nauplii derived from breeding to the juvenile stage [99]. Hence, in addition to the proper genetic management of aquaculture organisms, it is crucial to manage microbial communities in aquaculture pond water [100,101]. Therefore, the relationship between the genetic diversity of superior populations and their gut microbiome should be explored for more precise germplasm management.
5. Conclusions
This study aimed to develop transcriptome-derived cross-species microsatellite markers for genotyping and evaluating genetic polymorphisms in the Pacific blue shrimp species (L. stylirostris). Our findings demonstrate the successful development and utilization of these markers. The established transcriptome database offers and facilitates not only rapid primer selection for closely related species but also lays the groundwork for functional gene analysis and molecular marker development, particularly for economically important traits in selective breeding programs. Additionally, this study highlights the importance of establishing a suitable genetic management system to mitigate the risks of inbreeding depression and loss of genetic diversity when introducing new breeds in the future. Furthermore, the developed microsatellite markers play a crucial role in addressing key issues in blue shrimp aquaculture genetic management. By examining their association with functional genes, this study provides valuable insights into the underlying genetic mechanisms underlying disease resistance traits in aquaculture populations. These insights can inform future research directions and contribute to the development of effective strategies for maintaining the health and adaptability of aquaculture populations. The findings of this study can contribute to the genetic management analysis of the L. stylirostris population and enhance our understanding of the molecular genetics of this species. Moreover, the study aids in conserving and improving the shrimp species, ultimately facilitating the sustainable development of Pacific blue shrimps in aquaculture.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani14111685/s1, Figure S1: Statistics of the length distribution of unigenes identified in Litopenaeus stylirostris transcriptomic datasets. The abscissa represents the length, while the ordinate represents the number of unigenes; Figure S2: Species annotation distribution map of Litopenaeus stylirostris transcriptome alignment in the NR database. A total of 22,539 unigene entries were compared to the NR database. NR: Unigenes with NCBI non-redundant protein; Figure S3: Proportions of the amplified and un-amplified microsatellite primers for genetic diversity analysis in Litopenaeus stylirostris; Table S1: Statistics of the amount of information in the next-generation sequencing of the transcriptome of Litopenaeus stylirostris; Table S2: Assembly quality statistics of next-generation sequencing of Litopenaeus stylirostris transcriptomes; Table S3: Quality statistics of unigenes assembly for next-generation sequencing of Litopenaeus stylirostris transcriptomes; Table S4: Functional annotation of the Litopenaeus stylirostris transcriptome dataset against publicly available databases; Table S5: Statistics of prediction results of unigene coding sequence regions in the Litopenaeus stylirostris transcriptome; Table S6: Functional genes associated with shrimp disease resistance traits were identified from the Litopenaeus stylirostris transcriptome database, and primer pairs were developed to assess genetic polymorphisms in broodstock; Table S7: Primer pairs designed from the NCBI literature database and available for genetic polymorphism testing of broodstock in Litopenaeus stylirostris; Table S8: Correspondence between microsatellite loci and functional genes in Litopenaeus stylirostris.
Author Contributions
All authors contributed to the conception and design of the study. C.-W.H. participated in planning and overseeing project management. C.-W.H., T.-H.H., H.-Y.G. and I.C.L. conceived and designed the study. Y.-C.Y., P.-Y.C. and C.-C.C. performed biomaterial DNA sample preparation, data collection and analysis, and organized data for high-throughput SSR research. W.-C.Y. provided resources and supervision for the transcriptome sequencing of next-generation biomaterials. Y.-C.Y. prepared the manuscript. T.-H.H., H.-Y.G., I.C.L. and C.-W.H. discussed the results and commented on the manuscript. C.-W.H. revised and finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received partial support from the Ministry of Education through the Study Abroad and New Southbound Study Abroad Program (Grant numbers: 108001200N01), the New Agriculture Interdisciplinary Talent Cultivation Program (Grant numbers: J10929801), and the Fisheries Agency, Ministry of Agriculture, Taiwan, through the Annual Science and Technology Project of Research On Establishing Standardization And Conversion Criteria For Aquaculture Seedling Cultivation (Grant number: 112AS-6.1.1-FA-FB and 113AS-6.1.1-FA-05).
Institutional Review Board Statement
All experimental procedures, as well as the management and handling of all experimental animals, adhered to the guidelines approved by the Institutional Animal Care and Use Committee (IACUC) of National Taiwan Ocean University (Approval No. 113012), ensuring compliance with internationally recognized standards for the care and use of animals in research.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw reads of the RNA sequencing data of L. stylirostris generated in this study have been submitted to the NCBI sequence read and deposited in the GenBank Data Archive under the BioProject accession number PRJNA957851.
Acknowledgments
We thank the Ministry of Education (MoE) 2018–2019 Study Abroad and New Southbound Study Abroad Program for supporting the Department of Aquaculture, National Taiwan Ocean University (NTOU), the Fisheries Agency, Ministry of Agriculture, and Brunei Golden Corporation Sdn Bhd for facilitating overseas internships and bilateral industry–university cooperation. We give special thanks to Golden Company Chairman Richard Chuang, Kenneth Pang, Jacqueline Yeo, and other relevant colleagues for their invaluable assistance in collecting shrimp samples and coordinating the itinerary.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A


Figure A1.
Sample collection of commercially available fresh aquatic products of Pacific white and black tiger shrimps for genetic diversity analysis. (A) Thirty Pacific white shrimp (L. vannamei) samples with a nearly 1:1 male-to-female ratio were obtained from Fuguo Freezing Co., Ltd. in Yilan, Taiwan (produced in Taiwan), Charoen Pokphand Enterprise (Taiwan) Co., Ltd. in Taipei, Taiwan (produced in Thailand), and Lequality Life Co., Ltd. in Kaohsiung, Taiwan (produced in Nicaragua). The samples were labeled PWS-TW (n = 10), PWS-TH (n = 10), and PWS-NI (n = 10), respectively. (B) Thirty black tiger shrimp (P. monodon) samples with a nearly 1:1 male-to-female ratio were obtained from Sanheng International Trading Co., Ltd. in New Taipei, Taiwan (produced by Malaysia), Shangyi Fishery Co., Ltd. in Taichung, Taiwan (produced by Malaysia), and First Flight Frozen Food Co., Ltd. in Kaohsiung, Taiwan (produced in Vietnam). The samples were labeled BTS-MY1 (n = 10), BTS-MY2 (n = 10), and BTS-VN (n = 10), respectively. These samples were obtained from three Taiwanese supermarkets on 15–22 December 2021 and were selected to represent the genetic diversity of commercially available shrimps in this region. The samples were used for DNA extraction and genetic diversity analysis.
Figure A1.
Sample collection of commercially available fresh aquatic products of Pacific white and black tiger shrimps for genetic diversity analysis. (A) Thirty Pacific white shrimp (L. vannamei) samples with a nearly 1:1 male-to-female ratio were obtained from Fuguo Freezing Co., Ltd. in Yilan, Taiwan (produced in Taiwan), Charoen Pokphand Enterprise (Taiwan) Co., Ltd. in Taipei, Taiwan (produced in Thailand), and Lequality Life Co., Ltd. in Kaohsiung, Taiwan (produced in Nicaragua). The samples were labeled PWS-TW (n = 10), PWS-TH (n = 10), and PWS-NI (n = 10), respectively. (B) Thirty black tiger shrimp (P. monodon) samples with a nearly 1:1 male-to-female ratio were obtained from Sanheng International Trading Co., Ltd. in New Taipei, Taiwan (produced by Malaysia), Shangyi Fishery Co., Ltd. in Taichung, Taiwan (produced by Malaysia), and First Flight Frozen Food Co., Ltd. in Kaohsiung, Taiwan (produced in Vietnam). The samples were labeled BTS-MY1 (n = 10), BTS-MY2 (n = 10), and BTS-VN (n = 10), respectively. These samples were obtained from three Taiwanese supermarkets on 15–22 December 2021 and were selected to represent the genetic diversity of commercially available shrimps in this region. The samples were used for DNA extraction and genetic diversity analysis.


Appendix B
Table A1.
Functional genes associated with shrimp disease resistance traits were identified from the Litopenaeus stylirostris transcriptome database, and primer pairs were developed to assess genetic polymorphisms in broodstock.
Table A1.
Functional genes associated with shrimp disease resistance traits were identified from the Litopenaeus stylirostris transcriptome database, and primer pairs were developed to assess genetic polymorphisms in broodstock.
| Locus | GeneBank | Annotation Gene | Idenity | SSR Motif | Primer Sequence (5′-3′) | Tm (°C) | Size (bp) |
|---|---|---|---|---|---|---|---|
| Unigene1915 | XM_027362881.1:2331-3836 | double-stranded RNA-specific editase Adar-like | 93.06% | (AAT)7 | F: GTGGTCCACTGGTGCTATCC R: TCTAAGCGTGCTACTGTGATCA | 58 | 349 |
| CL3546.Contig1 | XM_027364033.1:34-1020 | c-type lectin | 93.51% | (GGA)5 | F: TCAACGAGATCATCAAAGCCCAG R: GAAAAACACTCGCTCATCACCTC | 58 | 124 |
| CL4833.Contig1 | XM_027351422.1:734-2935 | caspase | 95.1% | (GAA)5 | F: AGAGAAAGGAAGATCAAAAGAGGGA R: ACAAACTGTCGAAAGGAAGAAGC | 56 | 153 |
| CL517.Contig2 | XM_027376248.1:76-3636 | caspase | 95.35% | (AAT)8 | F: CATGGCCGTATAAAATCCAGCAC R: AGTCTTCCGCAGTGTAATTGGTA | 57 | 144 |
| CL5798.Contig1 | XM_027358108.1:14-908 | caspase Dronc-like | 95.08% | (AAG)6 | F: TGCTGTTCCTATGCAGTGGTAAT R: ATTTCAATCCCAGAGGCAGATGT | 58 | 129 |
| CL1947.Contig2 | XM_037936541.1:1062-2532 | chitinase | 85.46% | (ATA)5 | F: TCGACAAGAAAAACGAAATCCCT R: TTTTTGTAGAGGATGGGGGTGAC | 57 | 141 |
| CL701.Contig3 | XM_063730582.115-4747 | eukaryotic translation initiation factor 3 subunit A-like | 91.85% | (GTG)8 | F: CTTGGCGAACATCAGGACCT R: TCATCTTCTTCGCGGGATGG | 59 | 196 |
| CL997.Contig10_1 | XM_027378552.1:192-4841 | eukaryotic translation initiation factor 4 gamma 1-like | 98.69% | (GAG)6 | F: GTTACCAAGCCAACCCCAGA R: AGGGCTCCACTGATCATCCT | 59 | 355 |
| CL997.Contig10_2 | XM_027378552.1:192-4841 | eukaryotic translation initiation factor 4 gamma 1-like | 98.69% | (CCCAGC)5 | F: AGCACAGCATCAACCACAGA R: GGGACTGAGGTTGGGGTTG | 59 | 167 |
| CL997.Contig14_1 | XM_027378552.1:192-4846 | eukaryotic translation initiation factor 4 gamma 1-like | 98.69% | (GTG)6 | F: GCACCTTCTACTCCTGGCTG R: TGGTCGCGTCTTACAGGATG | 59 | 198 |
| CL997.Contig14_2 | XM_027378552.1:192-4846 | eukaryotic translation initiation factor 4 gamma 1-like | 98.69% | (TGGGGC)5 | F: GGGACTGAGGTTGGGGTTG R: CATCAACCACAGAAGCAGAAGTC | 58 | 160 |
| CL997.Contig14_3 | XM_027378552.1:192-4846 | eukaryotic translation initiation factor 4 gamma 1-like | 98.69% | (TCC)6 | F: TTTACAGGGCTCCACTGATCATC R: GAAGGAGAGGGAAGTGAATAGCA | 58 | 123 |
| CL997.Contig1 | XM_027378552.1:192-4846 | eukaryotic translation initiation factor 4 gamma 1-like | 98.69% | (GAG)6 | F: GAAGGAGAGGGAAGTGAATAGCA R: GATTTACAGGGCTCCACTGATCA | 58 | 125 |
| Unigene1091_1 | XM_047615203.1:41-3526 | eukaryotic translation initiation factor 5B-like | 91.76% | (GAT)5 | F: TGTCAAAGACGACTGGGACG R: CACTTCACCCTCCTGCACAT | 59 | 358 |
| Unigene1091_2 | XM_047615203.1:41-3526 | eukaryotic translation initiation factor 5B-like | 91.76% | (AAG)5 | F: CGCGGGCAAGAAGAAAAACA R: CCTCCTCCTTCTCTGCCTCT | 59 | 224 |
| Unigene159_1 | XM_027383341.1:566-7093 | eukaryotic translation initiation factor 2-alpha kinase 3-like | 96.6% | (GCTCCA)4 | F: TGGTTTGTGGTGTTGCTTGC R: GTAAACTGGGGTGTCTGGGG | 59 | 322 |
| Unigene159_2 | XM_027383341.1:566-7093 | eukaryotic translation initiation factor 2-alpha kinase 3-like | 96.6% | (GAG)5 | F: CCCTTTCTCTCATGGGGCTG R: GCAAACCCTCTGACTCCTCC | 60 | 239 |
| Unigene7147 | XM_027375483.1:74-2064 | eukaryotic translation initiation factor 2 | 98.75% | (TGA)5 | F: GCCCCCAAGATTGTCACAGA R: TAGCCACTTTCACAGCCTCG | 59 | 272 |
| CL1472.Contig13 | XM_043035729.1:18-7440 | myosin light chain kinase | 94.80% | (GAG)6 | F: AGAAAAGGAGGACGAACAGCAAT R: CAGGATCTCAGCTTCAGTGTCTT | 58 | 100 |
| Unigene5692 | XM_037925428.1:23-5851 | smooth muscle-like transcript variant X3 | 95.11% | (GAG)6 | F: GAGCTGCGACGAAGAAAAGGA R: GTTCTCCGTCTTCACCGTGT | 59 | 258 |
| Unigene6789_1 | XM_063745732.1:6-6814 | myosin light chain kinase | 91.31% | (GAC)5 | F: TGGAGCGGACGAAGAAGAAC R: TTCACCTCCAACTGACTCGC | 59 | 316 |
| Unigene6789_2 | XM_063745732.1:6-6814 | myosin light chain kinase | 91.31% | (AGG)5 | F: CGAGGAGAAGCTGGTCAAGG R: CCTGTCTCTCCAGCTCCTCT | 60 | 161 |
| CL1038.Contig1 | XM_027372703.1:146-2612 | ras-like GTP-binding protein Rho | 98.67% | (AAG)5 | F: AAGTTTTCGAGACAGCAACAAGG R: CAGACTGCATTGTTCATCCCATT | 57 | 119 |
| CL3247.Contig3 | XM_027361504.1:13-1366 | relish | 94.73% | (TTG)6 | F: AACCCAATGACTGCAGATTT R: GCACATGTGGATCCATCTGC | 55 | 89 |
| Unigene5458 | XM_027379198.1:3-934 | solute carrier family 35 member F6-like | 90.32% | (TGTT)5 | F: TGTCACACGCTCTGCAGAAT R: CCTTAGAACCCTCGGCATGG | 59 | 131 |
| CL2191.Contig1 | XM_047631710.1:12-1941 | serine protease | 84.08% | (TCC)6 | F: TGATGATGTTCGAGGGCCTC R: AGATGAAGGTGACGCAGGTG | 59 | 423 |
| Unigene7399 | XM_027353100.1:32-1233 | phenoloxidase-activating factor 2-like | 91.81% | (GGC)5 | F: TTGAGGTGCAACAAGTGTGA R: CGCACATTGGTTTGGAAGGG | 57 | 255 |
| CL2403.Contig1 | XM_027355035.1:264-2999 | serine protease | 95.22% | (CAG)6 | F: CAACGAAAAGATGAGGGCCG R: GAGGTTGACAGTCGGGTTGA | 58 | 265 |
| Unigene11623 | XM_027364731.1:17-2657 | phenoloxidase-activating factor 2-like | 95.67% | (GGT)5 | F: TGTTCCCTCCTCCATTGTGC R: CCTTCGACGTTTGCTGCAAG | 59 | 165 |
| Unigene7536 | XM_027382057.1:222-2273 | serine protease | 96.36% | (GGT)6 | F: CAGAGGGCCAGATGGTGAAG R: AGGAACGCGTTACCACTGAG | 59 | 281 |
| CL433.Contig2 | XM_047638789.1:82-6089 | large proline-rich protein BAG6-like | 94.24% | (GTG)6 | F: AGATGAGCTAGGACCTTGTGTTG R: CAGTTCCTCAAGCAATGGAGC | 58 | 132 |
| CL770.Contig4 | XM_027369909.1:43-7399 | leucine-rich repeat extensin-like protein 3 | 98.01% | (TGAA)5 | R: TGTTTACTCTGAATCCCTAACACA F: TCCTTTTTCTTCTTTCTTTCGTTTTTCC | 55 | 156 |
| CL997.Contig17 | XM_027378552.1:192-4843 | eukaryotic translation initiation factor | 98.69% | (TCC)6 | F: AGTGGGTTATTCTGGAGCTCAAG R: GCGAGAGGTTAACAATGTTCACG | 58 | 156 |
| CL997.Contig25 | XM_027378552.1:192-4846 | eukaryotic translation initiation factor | 98.69% | (GAG)6 | F: GAAGGAGAGGGAAGTGAATAGCA R: GATTTACAGGGCTCCACTGATCA | 58 | 125 |
| CL271.Contig8 | XM_037947861.1:1-3261 | MOG interacting and ectopic P-granules protein 1-like | 94.55% | (GGA)5 | F: AAGGAAAAGAAAGAGGGGGACAG R: TCCTTCCTCTTTCTTCTCCTCCT | 58 | 138 |
| CL1082.Contig1 | XM_027351461.1:621-3208 | BTB/POZ domain-containing protein 10-like | 98.88% | (GGA)8 | F: AGGATGAGAAGAAGAGGAGGAGT R: CTGCTCTTCTTCCTCCTCTTCC | 58 | 159 |
| CL1381.Contig28 | XM_027370486.1:668-5794 | serine/arginine repetitive matrix protein 2-like | 97.13% | (CGT)6 | F: AATTGATGAAAGTGCAGTGGTCG R: GTCGGAGGAAAGCGAAGAGAG | 58 | 122 |
| CL1472.Contig9 | XM_043035729.1:18-7440 | myosin light chain kinase | 94.80% | (GAG)7 | F: GAAAAGGAGGACGAACAGCAATC R: CAGGATCTCAGCTTCAGTGTCTT | 58 | 102 |
| CL1971.Contig3 | XR_010143192.1:355-2570 | NA | 90.87% | (ATC)6 | F: TTGGGTGTGGTTACTAGAGTTCG R: AAAGACAAAAAGGGAGTTGGTGG | 58 | 151 |
| CL1683.Contig2 | XM_027376841.1:5-1237 | serine/arginine repetitive matrix protein 2-like | 98.30% | (AAG)6 | F: GTTCGACGAAGAAGGACAAGAAG R: GCTCTTCTTCGACTTGTGCTTTT | 57 | 104 |
| CL2056.Contig2 | XM_027360043.1:1772-3836 | fibrous sheath CABYR-binding protein-like | 97.87% | (AGC)6 | F: CAGACCTCCAGTTTCAGTTCGAG R: AACTTCTCCATGATCTGCTCCTG | 58 | 131 |
Table A2.
Microsatellite marker tests available for the amplification of Litopenaeus stylirostris, Litopenaeus vannamei, and Penaeus monodon.
Table A2.
Microsatellite marker tests available for the amplification of Litopenaeus stylirostris, Litopenaeus vannamei, and Penaeus monodon.
| Locus | Motif | L. stylirostris | L. vannamei | P. monodon |
|---|---|---|---|---|
| Unigene1915 | (AAT)7 | ○ | ○ | – |
| CL4833.Contig1 | (GAA)5 | ○ | ● | △ |
| CL517.Contig2 | (AAT)8 | ○ | ● | ● |
| CL5798.Contig1 | (AAG)6 | ○ | ● | – |
| CL1947.Contig2 | (ATA)5 | ○ | – | △ |
| CL701.Contig3 | (GTG)8 | ○ | ○ | △ |
| CL997.Contig17 | (TCC)6 | ○ | ○ | – |
| Unigene159 | (GCTCCA)4 | – | – | – |
| Unigene7147 | (TGA)5 | ○ | ● | ● |
| CL1472.Contig13 | (GAG)6 | ○ | △ | △ |
| Unigene5692 | (GAG)6 | ○ | ● | ● |
| Unigene6789 | (GAC)5 | ○ | ● | ○ |
| CL3247.Contig3 | (TTG)6 | ○ | ● | △ |
| Unigene5458 | (TGTT)5 | ○ | △ | ○ |
| CL2191.Contig1 | (TCC)6 | ○ | △ | △ |
| Unigene11623 | (GGT)5 | ○ | ○ | ○ |
| Unigene7536 | (GGT)6 | ○ | ● | △ |
| CL433.Contig2 | (GTG)6 | – | – | – |
| CL770.Contig4 | (TGAA)5 | ○ | ○ | △ |
| CL1381.Contig28 | (CGT)6 | ○ | ○ | △ |
| CL1472.Contig9 | (GAG)7 | ● | ● | ● |
| CL1971.Contig3 | (ATC)6 | ○ | ○ | △ |
| CNM1 | (AAAAT)4 | ○ | ● | – |
| CNM2 | (AAAAT)3 | ○ | ● | △ |
| CNM3 | (ATTTT)4 | ○ | ● | ○ |
| CNM4 | (AAAAC)3 | ○ | ● | △ |
| CNM7 | (GCAA)4 | ○ | ○ | △ |
| CNM8 | (AAAT)4 | ○ | ● | △ |
| CNM11 | (TTGT)4 | ○ | ○ | – |
| CNM12 | (TAAA)4 | ○ | ● | △ |
| CNM13 | (AAAG)4 | ● | ● | – |
| CNM15 | (GATT)3 | ○ | ● | – |
| CNM18 | (CAA)6 | △ | – | ○ |
| CNM22 | (TGA)5 | ○ | ● | – |
| CNM23 | (TTA)5 | ○ | ● | △ |
| CNM26 | (ATT)5 | ○ | ○ | ○ |
| LV3 | (AG)9 | ○ | ● | – |
| LV13 | (CA)16 | ● | – | – |
| LV27 | (TC)14 | ○ | – | ○ |
| LV29 | (TAA)12 | ○ | ● | △ |
●: Can be successfully amplified with polymorphism; ○: can be successfully amplified without polymorphism; △: nonsense fragments are amplified; –: no amplified fragment.
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, L.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Robainas Barcia, A.; Espinosa López, G.; Hernández, D.; García-Machado, E. Temporal variation of the population structure and genetic diversity of Farfantepenaeus notialis assessed by allozyme loci. Mol. Ecol. 2005, 14, 2933–2942. [Google Scholar] [CrossRef] [PubMed]
- Murren, C.J.; Auld, J.R.; Callahan, H.; Ghalambor, C.K.; Handelsman, C.A.; Heskel, M.A.; Kingsolver, J.G.; Maclean, H.J.; Masel, J.; Maughan, H.; et al. Constraints on the evolution of phenotypic plasticity: Limits and costs of phenotype and plasticity. Heredity 2015, 115, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Masel, J.; King, O.D.; Maughan, H. The loss of adaptive plasticity during long periods of environmental stasis. Am. Nat. 2007, 169, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Laikre, L.; Hoban, S.; Bruford, M.W.; Segelbacher, G.; Allendorf, F.W.; Gajardo, G.; Rodríguez, A.G.; Hedrick, P.W.; Heuertz, M.; Hohenlohe, P.A.; et al. Post-2020 goals overlook genetic diversity. Science 2020, 367, 1083–1085. [Google Scholar] [CrossRef]
- Mondal, D.; Dutta, S.; Chakrabarty, U.; Mallik, A.; Mandal, N. Development and characterization of white spot disease linked microsatellite DNA markers in Penaeus monodon, and their application to determine the population diversity, cluster and structure. J. Invertebr. Pathol. 2019, 168, 107275. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, C.; Yu, Y.; Li, F. Analysis of genetic diversity and population structure of the ridgetail white prawn Exopalaemon carinicauda in China. Aquac. Rep. 2022, 27, 101369. [Google Scholar] [CrossRef]
- Stirton, W. Notes on North American Crustacea in the Museum of the Smithsonian Institution. No. III. Ann. Lyceum Nat. Hist. New York 1871, 10, 92–136. [Google Scholar]
- Liao, I.C. Penaeus monodon culture in Taiwan: Through two decades of growth. Int. J. Aquacult. Fish. Technol. 1989, 1, 16–24. Available online: http://ntour.ntou.edu.tw:8080/ir/handle/987654321/44620 (accessed on 2 February 2024).
- Bing, X.; Wang, J. A comparative study of nutritional quality in the muscle of Penaeus stylirostris and Penaeus vannamei in the cultured-pond. Acta Hydrobiol. Sin. 2006, 30, 458. Available online: https://api.semanticscholar.org/CorpusID:87831992 (accessed on 2 February 2024).
- Wyban, J. Shrimp farming diversification with SPF blue shrimp in 2011. In AQUA Culture Asia Pacific Magazine; Aqua Culture Asia Pacific: Nankin Row, Singapore, 2010; Volume 6, pp. 8–12. [Google Scholar]
- Bondad-Reantaso, M.G.; Lovell, E.R.; Arthur, J.R.; Hurwood, D.; Mather, P.B. Pathogen and Ecological Risk Analysis for the Introduction of Blue Shrimp, Litopenaeus stylirostris, from Brunei Darussalam to Fiji; SPC Aquaculture Technical Papers; Secretariat of the Pacific Community, Aquaculture Section: Noumea Cedex, New Caledonia, 2005; p. 80. [Google Scholar]
- Bondad-Reantaso, M.G.; Arthur, J.R. Pathogen risk analysis for aquaculture production. In Understanding and Applying Risk Analysis in Aquaculture; FAO Fisheries and Aquaculture Technical Paper. No. 519; Bondad-Reantaso, M.G., Arthur, J.R., Subasinghe, R.P., Eds.; FAO: Rome, Italy, 2008; pp. 27–46. [Google Scholar]
- de Lorgeril, J.; Saulnier, D.; Janech, M.G.; Gueguen, Y.; Bachère, E. Identification of genes that are differentially expressed in hemocytes of the Pacific blue shrimp (Litopenaeus stylirostris) surviving an infection with Vibrio penaeicida. Physiol. Genom. 2005, 21, 174–183. [Google Scholar] [CrossRef]
- Castro-Longoria, R.; Quintero-Arredondo, N.; Grijalva-Chon, J.M.; Ramos-Paredes, J. Detection of the yellow-head virus (YHV) in wild blue shrimp, Penaeus stylirostris, from the Gulf of California and its experimental transmission to the Pacific white shrimp, Penaeus vannamei. J. Fish Dis. 2008, 31, 953–956. [Google Scholar] [CrossRef]
- Bierne, N.; Beuzart, I.; Vonau, V.; Bonhomme, F.; Bédier, E. Microsatellite-associated heterosis in hatchery-propagated stocks of the shrimp Penaeus stylirostris. Aquaculture 2000, 184, 203–219. [Google Scholar] [CrossRef]
- Goyard, E.; Arnaud, S.; Vonau, V.; Bishoff, V.; Mouchel, O.; Pham, D.; Wyban, J.; Boudry, P. Residual genetic variability in domesticated populations of the Pacific blue shrimp (Litopenaeus stylirostris) of New Caledonia, French Polynesia and Hawaii and some management recommendations. Aquat. Living Resour. 2003, 16, 501–508. [Google Scholar] [CrossRef]
- Orosco, F.L.; Lluisma, A.O. Prevalence, diversity and co-occurrence of the white spot syndrome virus, monodon baculovirus and Penaeus stylirostris densovirus in wild populations of Penaeus monodon in the Philippines. Dis. Aquat. Organ. 2017, 125, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E.; Sonesson, A.K.; Gebregiwergis, G.; Woolliams, J.A. Management of Genetic Diversity in the Era of Genomics. Front. Genet. 2020, 11, 472717. [Google Scholar] [CrossRef]
- Choi, E.; Kim, S.H.; Lee, S.J.; Jo, E.; Kim, J.; Kim, J.H.; Parker, S.J.; Chi, Y.M.; Park, H. A First Genome Survey and Genomic SSR Marker Analysis of Trematomus loennbergii Regan, 1913. Animals 2021, 11, 3186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chiodini, R.; Badr, A.; Zhang, G. The impact of next-generation sequencing on genomics. J. Genet. Genom. 2011, 38, 95–109. [Google Scholar] [CrossRef]
- Chu, P.Y.; Li, J.X.; Hsu, T.H.; Gong, H.Y.; Lin, C.Y.; Wang, J.H.; Huang, C.W. Identification of Genes Related to Cold Tolerance and Novel Genetic Markers for Molecular Breeding in Taiwan Tilapia (Oreochromis spp.) via Transcriptome Analysis. Animals 2021, 11, 3538. [Google Scholar] [CrossRef]
- Ma, K.Y.; Feng, J.B.; Xie, N.; Feng, X.Y.; Li, J.L. Microsatellite analysis of genetic variation of the oriental river prawn Macrobrachium nipponense in Qiantang River. Zool. Res. 2011, 32, 363–370. [Google Scholar] [CrossRef]
- Lu, X.; Kong, J.; Meng, X.; Cao, B.; Luo, K.; Dai, P.; Luan, S. Identification of SNP markers associated with tolerance to ammonia toxicity by selective genotyping from de novo assembled transcriptome in Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 73, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Vu, N.T.T.; Zenger, K.R.; Silva, C.N.S.; Guppy, J.L.; Jerry, D.R. Population Structure, Genetic Connectivity, and Signatures of Local Adaptation of the Giant Black Tiger Shrimp (Penaeus monodon) throughout the Indo-Pacific Region. Genome Biol. Evol. 2021, 13, evab214. [Google Scholar] [CrossRef] [PubMed]
- Bonthala, B.; Abdin, M.Z.; Arya, L.; Pandey, C.D.; Sharma, V.; Yadav, P.; Verma, M. Genome-wide SSR markers in bottle gourd: Development, characterization, utilization in assessment of genetic diversity of National Genebank of India and synteny with other related cucurbits. J. Appl. Genet. 2022, 63, 237–263. [Google Scholar] [CrossRef] [PubMed]
- Vaseeharan, B.; Rajakamaran, P.; Jayaseelan, D.; Vincent, A.Y. Molecular markers and their application in genetic diversity of penaeid shrimp. Aquacult. Int. 2013, 21, 219–241. [Google Scholar] [CrossRef]
- Rezaee, S.; Farahmand, H.; Nematollahi, M.A. Genetic diversity status of Pacific white shrimp (Litopenaeus vannamei) using SSR markers in Iran. Aquacult. Int. 2016, 24, 479–489. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y.; Chen, Q.; Wang, M.; Li, Q.; Zhou, W.; Cheng, Y. Genetic Diversity Analysis of the Red Swamp Crayfish Procambarus clarkii in Three Cultured Populations Based on Microsatellite Markers. Animals 2023, 13, 1881. [Google Scholar] [CrossRef]
- Zhu, K.; Yu, W.; Huang, J.; Zhou, F.; Guo, H.; Zhang, N.; Jiang, S.; Zhang, D. Parentage determination in black tiger shrimp (Penaeus monodon) based on microsatellite DNA markers. Aquacult. Int. 2017, 25, 827–836. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, C.; Lin, K.; You, W.; Yang, Z. Genetic Diversity and Genetic Structure among Four Selected Strains of Whiteleg Shrimp (Litopenaeus vannamei) Using SSR Markers. Fishes 2023, 8, 544. [Google Scholar] [CrossRef]
- Kaya, S.; Kabasakal, B.; Erdoğan, A. Geographic Genetic Structure of Alectoris chukar in Türkiye: Post-LGM-Induced Hybridization and Human-Mediated Contaminations. Biology 2023, 12, 401. [Google Scholar] [CrossRef]
- An, H.S.; Shin, E.H.; Lee, J.W.; Nam, M.M.; Myeong, J.I.; An, C.M. Comparative genetic variability between broodstock and offspring populations of Korean starry flounder used for stock enhancement in a hatchery by using microsatellite DNA analyses. Genet. Mol. Res. 2013, 12, 6319–6330. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.; Li, Z.; Xu, W.; Dong, J.; Wei, H.; Li, Y.; Li, X. Genetic diversity and structure of Chinese grass shrimp, Palaemonetes sinensis, inferred from transcriptome-derived microsatellite markers. BMC Genet. 2019, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.R.; Barateli, L.O.; de Souza, U.J.B.; Pereira, A.M.S.; Bertoni, B.W.; Telles, M.P.C. Development and characterization of microsatellite markers in Stryphnodendron adstringens (Leguminosae). Physiol. Mol. Biol. Plants 2020, 26, 2095–2101. [Google Scholar] [CrossRef]
- Bernardes, V.; Murakami, D.M.; Bizão, N.; Souza, T.N.; da Silva, M.J.; Telles, M.P.C. Transferability and characterization of microsatellite markers from Byrsonima cydoniifolia A. Juss. (MALPIGHIACEAE) in seven related taxa from Cerrado biome reveal genetic relationships. Mol. Biol. Rep. 2021, 48, 4039–4046. [Google Scholar] [CrossRef] [PubMed]
- Alves, F.; Martins, F.M.S.; Areias, M.; Muñoz-Mérida, A. Automating microsatellite screening and primer design from multi-individual libraries using Micro-Primers. Sci. Rep. 2022, 12, 295. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, T.; Zhu, C.; Jiang, X.; Zhao, Y.; Xu, Z.; Yang, S.; Chen, A. Selection and use of microsatellite markers for individual identification and meat traceability of six swine breeds in the Chinese market. Food Sci. Technol. Int. 2018, 24, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Natonek-Wiśniewska, M.; Piestrzynska-Kajtoch, A.; Koseniuk, A.; Krzyścin, P. Current Analytical Methods and Research Trends Are Used to Identify Domestic Pig and Wild Boar DNA in Meat and Meat Products. Genes 2022, 13, 1825. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, C.C.; Huang, C.W.; Lin, C.Y.; Ho, C.H.; Pham, H.N.; Hsu, T.H.; Lin, T.T.; Chen, R.H.; Yang, S.D.; Chang, C.I.; et al. Development of Disease-Resistance-Associated Microsatellite DNA Markers for Selective Breeding of Tilapia (Oreochromis spp.) Farmed in Taiwan. Genes 2022, 13, 99. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Weng, Z.; Yang, Y.; Hua, S.; Zhang, H.; Meng, Z. Genetic Evaluation of Black Sea Bream (Acanthopagrus schlegelii) Stock Enhancement in the South China Sea Based on Microsatellite DNA Markers. Fishes 2021, 6, 47. [Google Scholar] [CrossRef]
- Gaynor, K.M.; Solomon, J.W.; Siller, S.; Jessell, L.; Duffy, J.E.; Rubenstein, D.R. Development of genome- and transcriptome-derived microsatellites in related species of snapping shrimps with highly duplicated genomes. Mol. Ecol. Resour. 2017, 17, e160–e173. [Google Scholar] [CrossRef]
- Mohanty, P.; Sahoo, L.; Pillai, B.R.; Jayasankar, P.; Das, P. Genetic divergence in indian populations of M. rosenbergii using microsatellite markers. Aquac. Res. 2016, 47, 472–481. [Google Scholar] [CrossRef]
- Min, J.G.; Jeong, H.D.; Kim, K.I. Identification of Various InDel-II Variants of the White Spot Syndrome Virus Isolated from Frozen Shrimp and Bivalves Obtained in the Korean Commercial Market. Animals 2023, 27, 3348. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef]
- de Hoon, M.J.; Imoto, S.; Nolan, J.; Miyano, S. Open source clustering software. Bioinformatics 2004, 20, 1453–1454. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Vonau, V.; Ohresser, M.; Bierne, N.; Delsert, C.; Beuzart, I.; Bedier, E.; Bonhomme, F. Three polymorphic microsatellites in the shrimp Penaeus stylirostris. Anim. Genet. 1999, 30, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Meehan, D.; Xu, Z.; Zuniga, G.; Alcivar-Warren, A. High frequency and large number of polymorphic microsatellites in cultured shrimp, Penaeus (Litopenaeus) vannamei [Crustacea:Decapoda]. Mar. Biotechnol. 2003, 5, 311–330. [Google Scholar] [CrossRef]
- Pérez, F.; Ortiz, J.; Zhinaula, M.; Gonzabay, C.; Calderón, J.; Volckaert, F.A. Development of EST-SSR markers by data mining in three species of shrimp: Litopenaeus vannamei, Litopenaeus stylirostris, and Trachypenaeus birdy. Mar. Biotechnol. 2005, 7, 554–569. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Thiel, T.; Michalek, W.; Varshney, R.K.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Peregrino-Uriarte, A.B.; Varela-Romero, A.; Muhlia-Almazán, A.; Anduro-Corona, I.; Vega-Heredia, S.; Gutiérrez-Millán, L.E.; De la Rosa-Vélez, J.; Yepiz-Plascencia, G. The complete mitochondrial genomes of the yellowleg shrimp Farfantepenaeus californiensis and the blue shrimp Litopenaeus stylirostris (Crustacea: Decapoda). Comp. Biochem. Physiol. Part D Genom. Proteom. 2009, 4, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Hizer, S.E.; Dhar, A.K.; Klimpel, K.R.; Garcia, D.K. RAPD markers as predictors of infectious hypodermal and hematopoietic necrosis virus (IHHNV) resistance in shrimp (Litopenaeus stylirostris). Genome 2002, 45, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Arulmoorthy, M.P.; Anandajothi, E.; Vasudevan, S.; Suresh, E. Major viral diseases in culturable penaeid shrimps: A review. Aquacult. Int. 2020, 28, 1939–1967. [Google Scholar] [CrossRef]
- Chow, L.H.; Ma, K.Y.; Hui, J.H.L.; Chu, K.H. Isolation and Characterization of Polymorphic Microsatellite Loci for Caridina cantonensis and Transferability Across Eight Confamilial Species (Atyidae, Decapoda). Zool. Stud. 2018, 57, e19. [Google Scholar] [CrossRef]
- Kawahara-Miki, R.; Wada, K.; Azuma, N.; Chiba, S. Expression profiling without genome sequence information in a non-model species, Pandalid shrimp (Pandalus latirostris), by next-generation sequencing. PLoS ONE 2011, 6, e26043. [Google Scholar] [CrossRef] [PubMed]
- Grogan, K.E.; McGinnis, G.J.; Sauther, M.L.; Cuozzo, F.P.; Drea, C.M. Next-generation genotyping of hypervariable loci in many individuals of a non-model species: Technical and theoretical implications. BMC Genom. 2016, 17, 204. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Lee Abdullah, T.; Yusop, M.R.; Hanafi, M.M.; Sahebi, M.; Azizi, P.; Shamshiri, R.R. Mining and development of novel SSR markers using next generation sequencing (NGS) data in plants. Molecules 2018, 23, 399. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, S.; Liu, Y.; Wang, P.; Zhang, Z.; Chen, D. A pipeline for effectively developing highly polymorphic simple sequence repeats markers based on multi-sample genomic data. Ecol. Evol. 2022, 12, e8705. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Chen, X.; Peng, J.; Yang, C.; Peng, M.; Zhu, W.; Xie, D.; He, P.; Wei, P.; Lin, Y.; et al. Single-molecule long-read sequencing facilitates shrimp transcriptome research. Sci. Rep. 2018, 8, 16920. [Google Scholar] [CrossRef]
- Van Quyen, D.; Gan, H.M.; Lee, Y.P.; Nguyen, D.D.; Nguyen, T.H.; Tran, X.T.; Nguyen, V.S.; Khang, D.D.; Austin, C.M. Improved genomic resources for the black tiger prawn (Penaeus monodon). Mar. Genom. 2020, 52, 100751. [Google Scholar] [CrossRef] [PubMed]
- Kawato, S.; Nishitsuji, K.; Arimoto, A.; Hisata, K.; Kawamitsu, M.; Nozaki, R.; Kondo, H.; Shinzato, C.; Ohira, T.; Satoh, N.; et al. Genome and transcriptome assemblies of the kuruma shrimp, Marsupenaeus japonicus. G3 2021, 11, jkab268. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhuang, X.; Liao, M.; Huang, L.; Cui, Q.; Liu, C.; Dong, W.; Wang, F.; Liu, Y.; Wang, W. Transcriptome analysis of Pacific white shrimp (Litopenaeus vannamei) hepatopancreas challenged by Vibrio alginolyticus reveals lipid metabolic disturbance. Fish Shellfish Immunol. 2022, 123, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhai, Y.; He, P.; Jia, R. Comprehensive Transcriptomic and Metabolomic Analysis of the Litopenaeus vannamei Hepatopancreas After WSSV Challenge. Front Immunol. 2022, 13, 826794. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, J.; Zhang, X.; Yu, Y.; Li, F. Comparative transcriptomic analysis unveils a network of energy reallocation in Litopenaeus vannamei responsive to heat-stress. Ecotoxicol. Environ. Saf 2022, 238, 113600. [Google Scholar] [CrossRef]
- Rao, R.; Bhassu, S.; Bing, R.Z.; Alinejad, T.; Hassan, S.S.; Wang, J. A transcriptome study on Macrobrachium rosenbergii hepatopancreas experimentally challenged with white spot syndrome virus (WSSV). J. Invertebr. Pathol. 2016, 136, 10–22. [Google Scholar] [CrossRef]
- Kim, J.O.; Kim, J.O.; Kim, W.S.; Oh, M.J. Characterization of the Transcriptome and Gene Expression of Brain Tissue in Sevenband Grouper (Hyporthodus septemfasciatus) in Response to NNV Infection. Genes 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; He, Z.; Chen, X.; Huang, Y.; Xie, J.; Qin, X.; Ni, Z.; Sun, C. Growth trait gene analysis of kuruma shrimp (Marsupenaeus japonicus) by transcriptome study. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100874. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, A.; Moghadam, H.; Larsson, T.; Afanasyev, S.; Mørkøre, T. Gene expression profiling in melanised sites of Atlantic salmon fillets. Fish Shellfish Immunol. 2016, 55, 56–63. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, D354–D357. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yuan, X.; Qiu, Q.; Huang, G.; Jiang, Q.; Fu, P.; Zhang, Y.; Jia, Y.; Yang, X.; Jiang, H. Comparative transcriptome analysis of gonads for the identification of sex-related genes in giant freshwater prawns (Macrobrachium Rosenbergii) using RNA sequencing. Genes 2019, 10, 1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, S.; Wang, C.; Shi, L.; Guo, H.; Chan, S. Gill transcriptomes reveal involvement of cytoskeleton remodeling and immune defense in ammonia stress response in the banana shrimp Fenneropenaeus merguiensis. Fish Shellfish Immunol. 2017, 71, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ge, Q.; Li, J.; Gao, H.; Li, J.; Zhao, F. Identification of novel EST-SSR markers by transcriptome sequencing in ridgetail white prawn Exopalaemon carinicauda. Genes Genom. 2018, 40, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wei, P.; Zhang, B.; Zhao, Y.; Zeng, D.; Chen, X.; Li, M.; Chen, X. Gonadal transcriptomic analysis and differentially expressed genes in the testis and ovary of the Pacific white shrimp (Litopenaeus vannamei). BMC Genom. 2015, 16, 1006. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Chu, P.Y.; Wu, Y.F.; Chan, W.R.; Wang, Y.H. Identification of Functional SSR Markers in Freshwater Ornamental Shrimps Neocaridina denticulata Using Transcriptome Sequencing. Mar. Biotechnol. 2020, 22, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.H.; Chiu, Y.T.; Lee, H.T.; Gong, H.Y.; Huang, C.W. Development of EST-Molecular Markers from RNA Sequencing for Genetic Management and Identification of Growth Traits in Potato Grouper (Epinephelus tukula). Biology 2021, 10, 36. [Google Scholar] [CrossRef]
- Bernard, A.M.; Richards, V.P.; Stanhope, M.J.; Shivji, M.S. Transcriptome-derived microsatellites demonstrate strong genetic differentiation in pacific white sharks. J. Hered. 2018, 109, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Li, L.F.; Yin, D.X.; Song, N.; Tang, E.H.; Xiao, H.X. Genomic and EST microsatellites for Rhododendron aureum (Ericaceae) and cross-amplification in other congeneric species. Am. J. Bot. 2011, 98, e250–e252. [Google Scholar] [CrossRef]
- Marasinghe, M.S.L.R.P.; Nilanthi, R.M.R.; Hathurusinghe, H.A.B.M.; Sooriyabandara, M.G.C.; Chandrasekara, C.H.W.M.R.B.; Jayawardana, K.A.N.C.; Kodagoda, M.M.; Rajapakse, R.C.; Bandaranayake, P.C.G. Revisiting traditional SSR based methodologies available for elephant genetic studies. Sci. Rep. 2021, 11, 8718. [Google Scholar] [CrossRef]
- Dai, X.; Huang, X.; Zhang, Z.; Zhang, R.; Cao, X.; Zhang, C.; Wang, K.; Ren, Q. Molecular cloning and expression analysis of two type II crustin genes in the oriental river prawn, Macrobrachium nipponense. Fish Shellfish Immunol. 2020, 98, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, S.; Dou, Y. Hormonal regulation changes in shrimp infected with Enterocytozoon hepatopenaei: A transcriptomic analysis. Aquacult. Int. 2023, 32, 2841–2863. [Google Scholar] [CrossRef]
- Chen, H.Y.; Toullec, J.Y.; Lee, C.Y. The Crustacean Hyperglycemic Hormone Superfamily: Progress Made in the Past Decade. Front. Endocrinol. 2020, 11, 578958. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Na, R.; Chen, H.; Lou, B.; Niu, B. RNA sequencing and functional analysis of adult gonadal tissue to identify candidate key genes in Macrobrachium rosenbergii sex development. Aquacult. Int. 2021, 29, 2805–2821. [Google Scholar] [CrossRef]
- Farhadi, A.; Tang, S.; Huang, M.; Yu, Q.; Xu, C.; Li, E. Identification of key immune and stress related genes and pathways by comparative analysis of the gene expression profile under multiple environmental stressors in pacific white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2023, 135, 108695. [Google Scholar] [CrossRef]
- Wang, Z.; Liao, G.; Chen, B.; Fan, L. Impacts of acute cold-stress in Pacific white shrimp Litopenaeus vannamei: Investigating the tissue-specific stress resistance response. Aquacult. Int. 2023, 31, 2649–2663. [Google Scholar] [CrossRef]
- Duan, Y.; Nan, Y.; Zhu, X.; Yang, Y.; Xing, Y. The adverse impacts of ammonia stress on the homeostasis of intestinal health in Pacific white shrimp (Litopenaeus vannamei). Environ. Pollut. 2024, 340, 122762. [Google Scholar] [CrossRef]
- Rodríguez-Viera, L.; Alpízar-Pedraza, D.; Mancera, J.M.; Perera, E. Toward a More Comprehensive View of α-Amylase across Decapods Crustaceans. Biology 2021, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.K.; Dettori, A.; Roux, M.M.; Klimpel, K.R.; Read, B. Identification of differentially expressed genes in shrimp (Penaeus stylirostris) infected with White spot syndrome virus by cDNA microarrays. Arch. Virol. 2003, 148, 2381–2396. [Google Scholar] [CrossRef]
- Niu, S.; Yang, L.; Zuo, H.; Zheng, J.; Weng, S.; He, J.; Xu, X. A chitinase from pacific white shrimp Litopenaeus vannamei involved in immune regulation. Dev. Comp. Immunol. 2018, 85, 161–169. [Google Scholar] [CrossRef]
- Huang, X.; Huang, Z.; Sun, L.; Qiu, M.; Deng, Q.; Fang, Z.; Wang, Y. Protective mechanisms of three antioxidants against T-2 toxin-induced muscle protein deterioration in shrimp. J. Sci. Food. Agric. 2022, 102, 4883–4891. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Q.; Wang, S.; He, J.; Li, C. Stimulator of interferon genes defends against bacterial infection via IKKβ-mediated Relish activation in shrimp. Front. Immunol. 2022, 13, 977327. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Shen, M.; He, Y.; Li, B.; Pu, L.; Xia, G.; Yang, M.; Wang, G. Analysis of differentially expressed proteins after EHP-infection and characterization of caspase 3 protein in the whiteleg shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2023, 135, 108698. [Google Scholar] [CrossRef] [PubMed]
- Wongdontri, C.; Jaree, P.; Somboonwiwat, K. PmKuSPI is regulated by pmo-miR-bantam and contributes to hemocyte homeostasis and viral propagation in shrimp. Fish Shellfish Immunol. 2023, 137, 108738. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zheng, M.; Liu, J.; Jin, Z.; Wang, C.; Gao, M.; Zhang, H.; Zhang, X.; Xia, X. LDLa containing C-type lectin mediates phagocytosis of V.anguillarum and regulates immune effector genes in shrimp. Fish Shellfish Immunol. 2024, 145, 109361. [Google Scholar] [CrossRef] [PubMed]
- Giraud, C.; Callac, N.; Boulo, V.; Lam, J.S.; Pham, D.; Selmaoui-Folcher, N.; Wabete, N. The Active Microbiota of the Eggs and the Nauplii of the Pacific Blue Shrimp Litopenaeus stylirostris Partially Shaped by a Potential Vertical Transmission. Front. Microbiol. 2022, 13, 886752. [Google Scholar] [CrossRef] [PubMed]
- Callac, N.; Boulo, V.; Giraud, C.; Beauvais, M.; Ansquer, D.; Ballan, V.; Maillez, J.R.; Wabete, N.; Pham, D. Microbiota of the Rearing Water of Penaeus stylirostris Larvae Influenced by Lagoon Seawater and Specific Key Microbial Lineages of Larval Stage and Survival. Microbiol. Spectr. 2022, 10, e0424122. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, P.; Li, Y.; Song, D.; Long, W.; Wang, Z.; Yi, S.; Jiang, L. Gut microbiota, host genetics and phenotypes in aquatic animals: A review. Aquac. Rep. 2023, 31, 101648. [Google Scholar] [CrossRef]
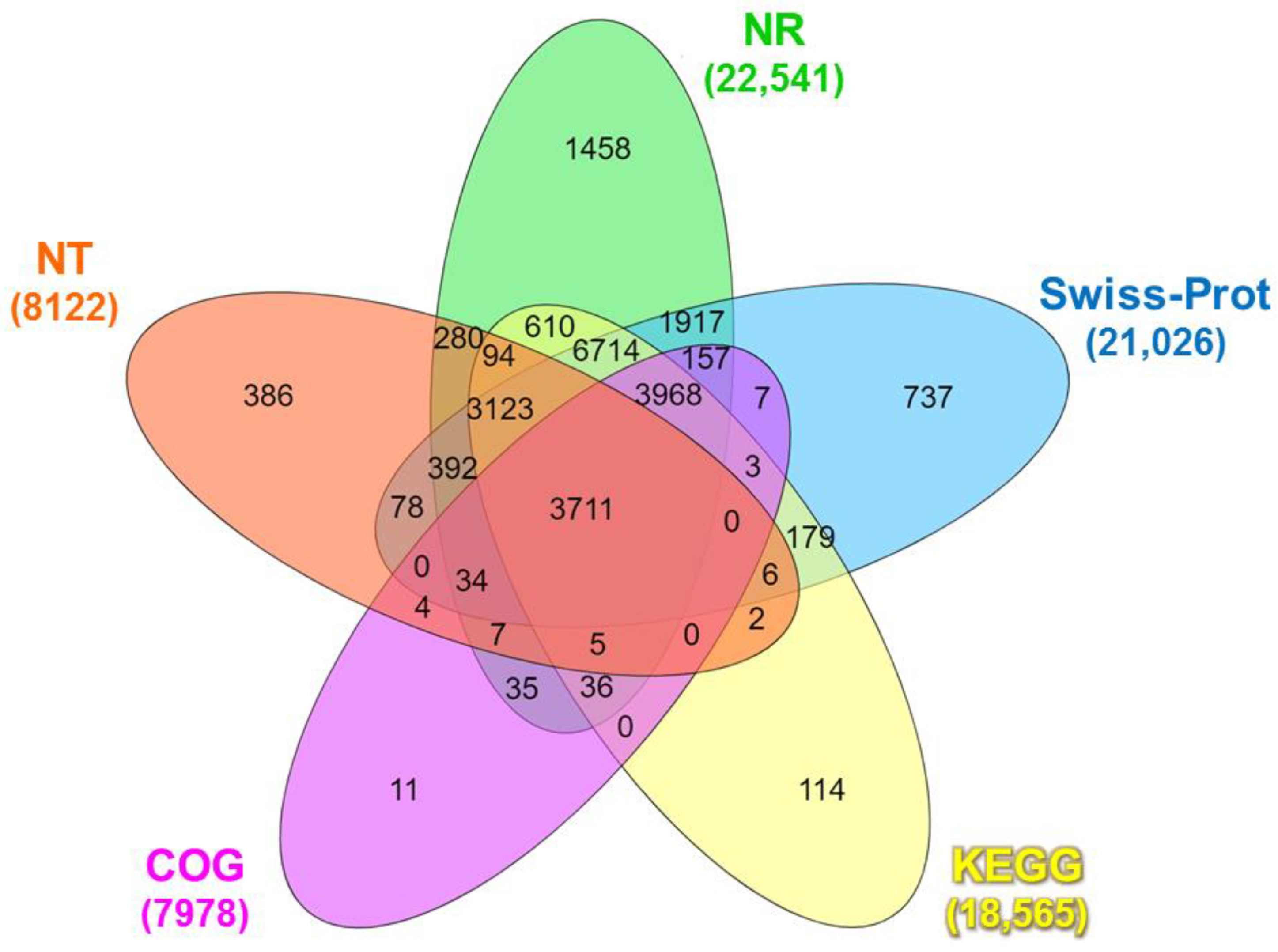
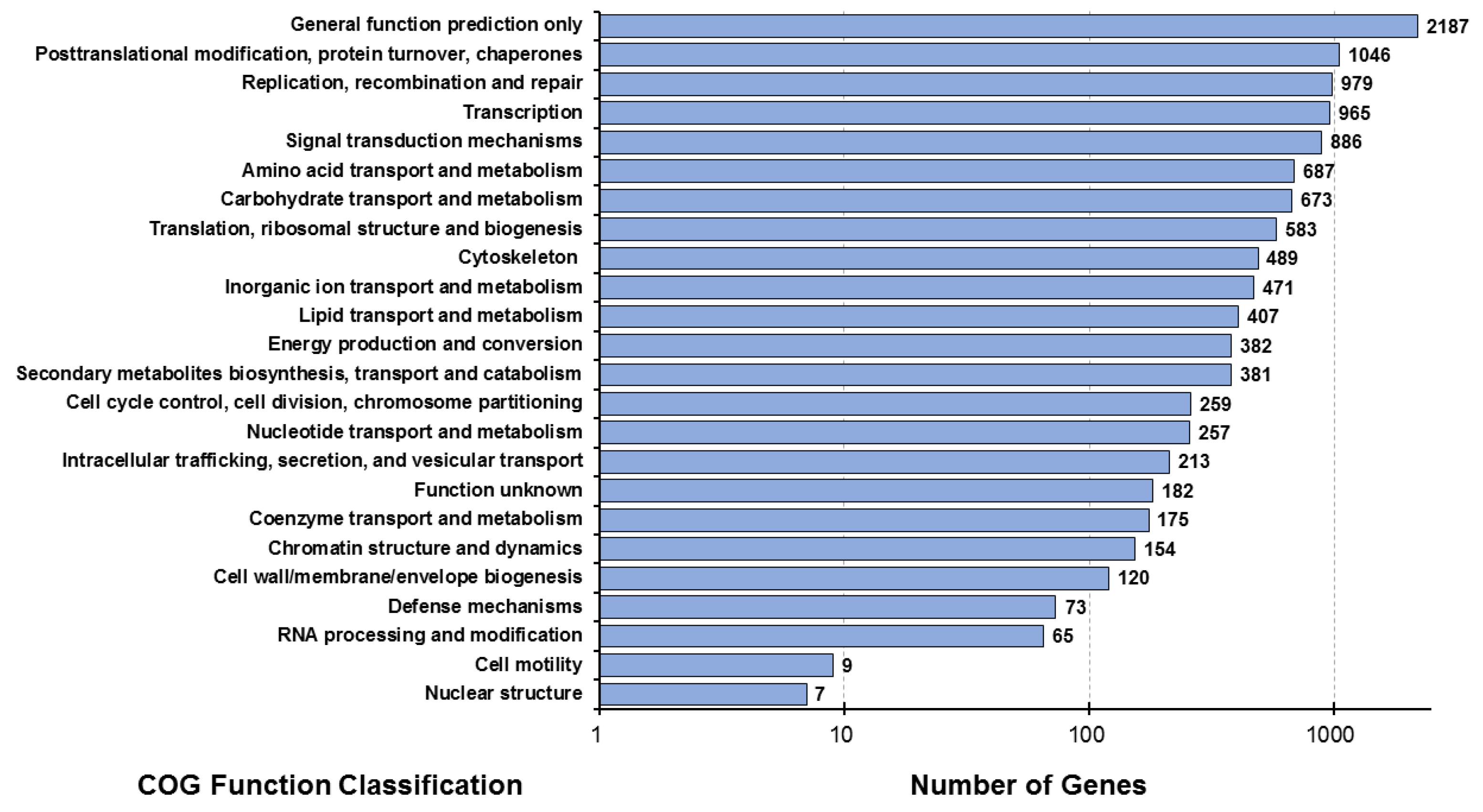
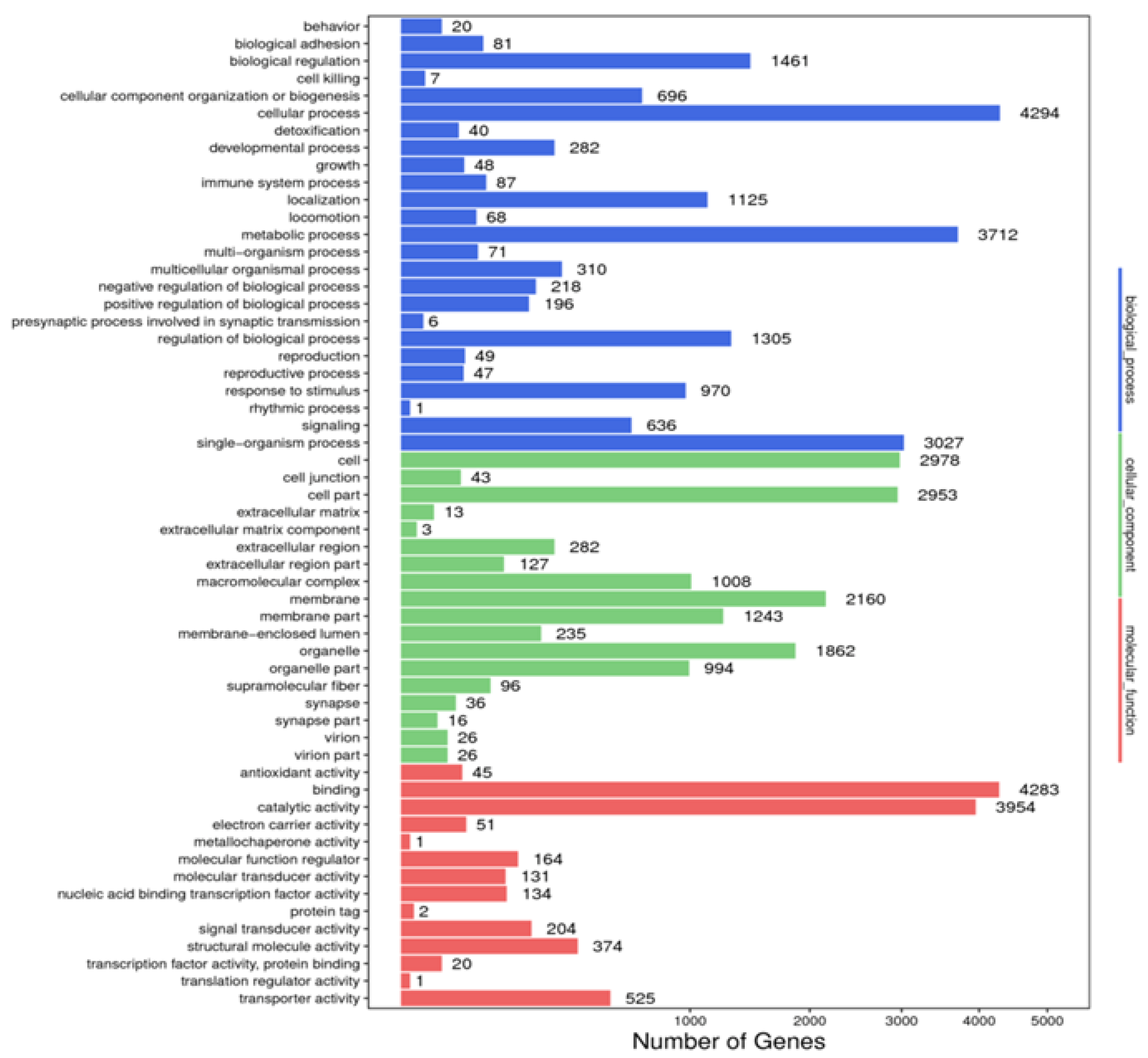
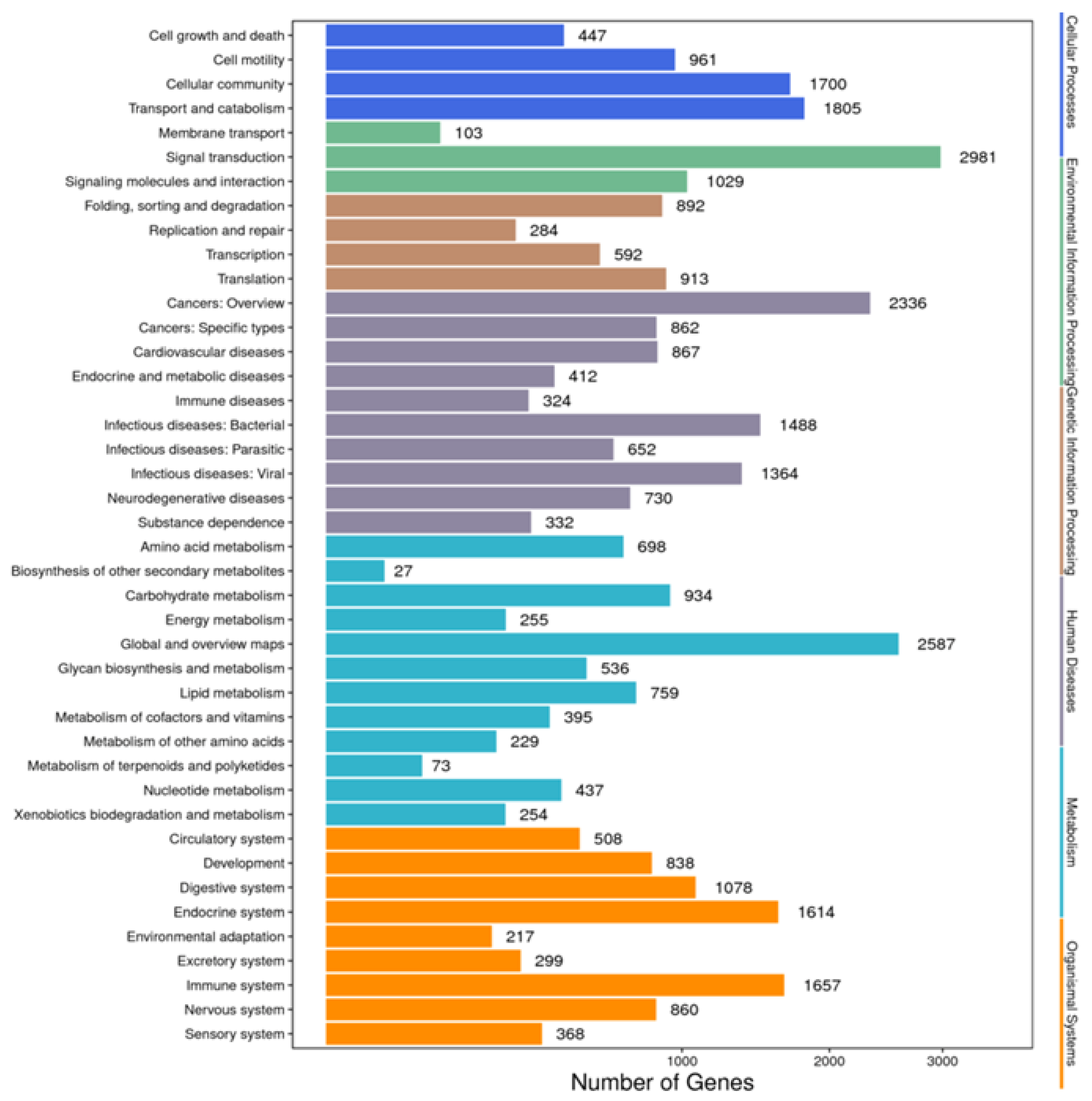
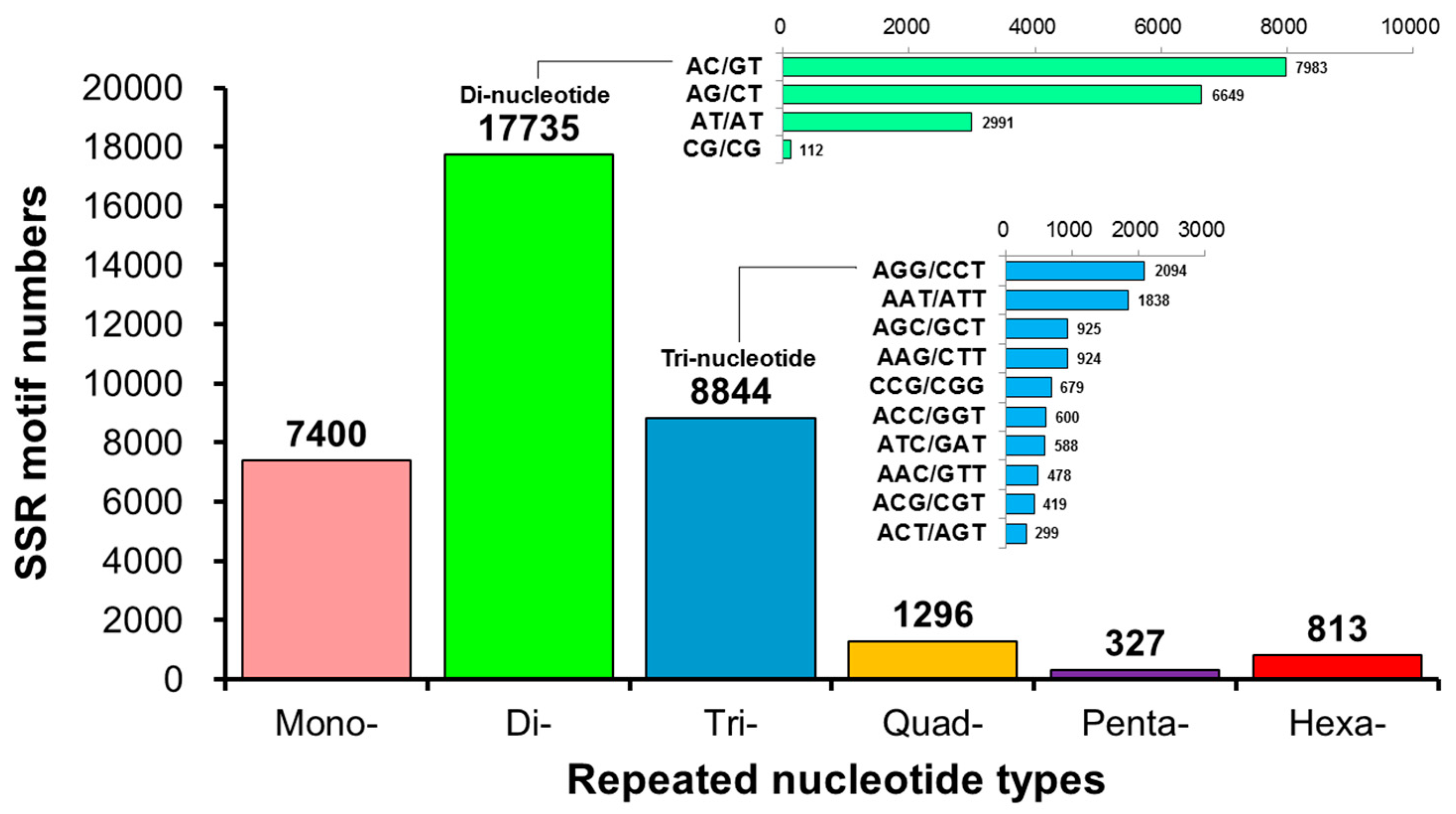
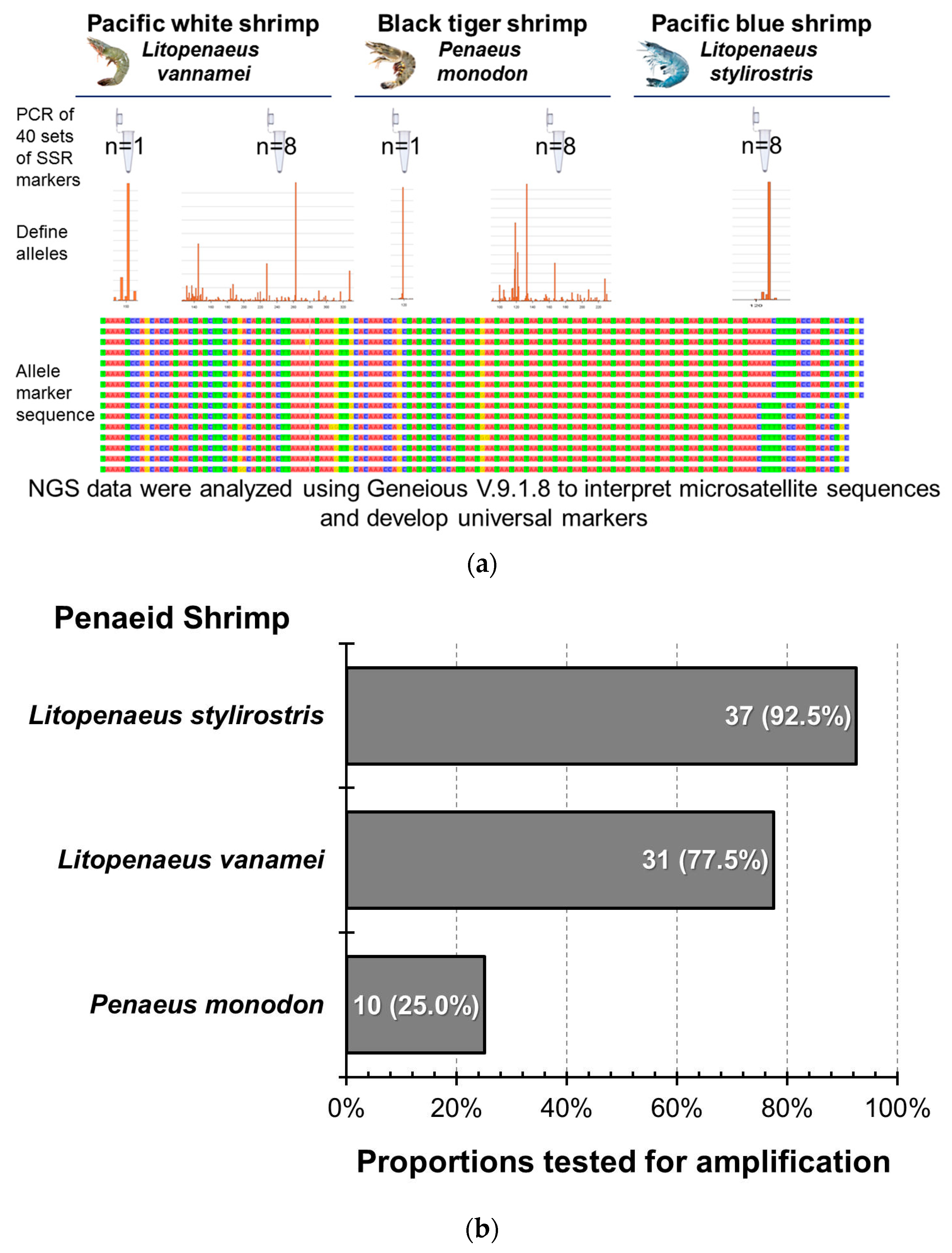
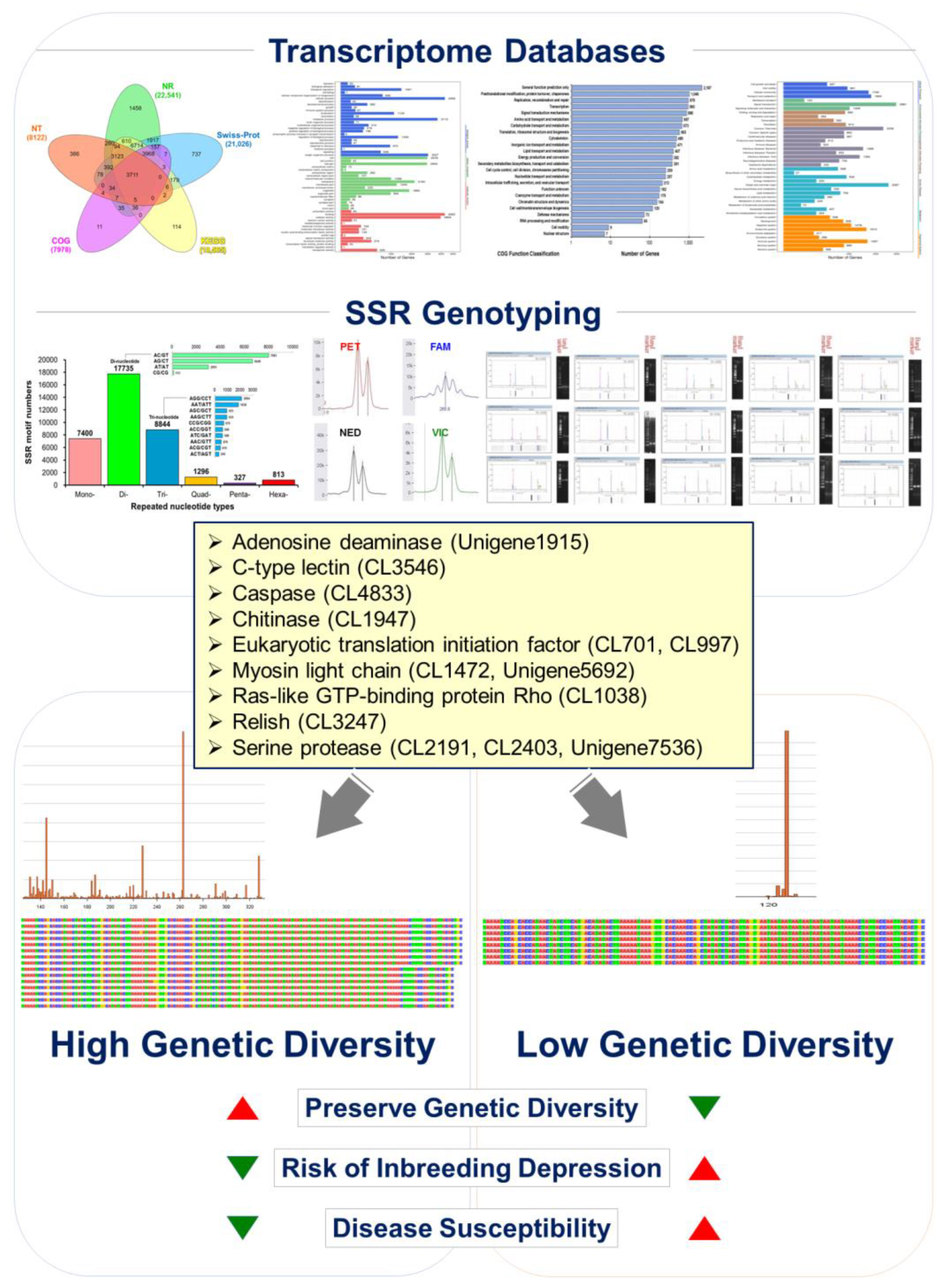
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).